基于轉錄組測序的火龍果果肉不同發育時期淀粉和蔗糖代謝途徑相關基因差異表達分析
2021-08-04 07:31:38楊運良李建勛馬革農
熱帶作物學報 2021年6期
楊運良 李建勛 馬革農

摘? 要:采用Illumina Novaseq 6000測序技術對火龍果果肉3個時期(幼果期、轉色期、成熟期)的樣品進行轉錄組測序,對獲得的Unigene進行7大數據庫注釋,共有3617個Unigene成功注釋在所有數據庫中,進一步篩選與淀粉和蔗糖代謝途徑相關的5個酶基因并進行實時熒光定量PCR(qPCR)基因表達驗證。結果表明:轉錄組測序共得到115 193條轉錄本和47 855條Unigene,N50為2000。GO功能富集分析結果表明,18 582個Unigene在GO功能注釋中被分為生物過程、細胞組成及分子功能3大類。KOG功能分類結果表明,6203個Unigene在KOG注釋中被分為25個組,其中翻譯后修飾、蛋白質周轉、伴侶包含的Unigene最多,共854個。KEGG代謝通路注釋結果表明,6554個Unigene獲得功能注釋,共有119條代謝途徑,其中碳水化合物代謝所占比例最高。篩選出UGP2、glgC、glgA、Cluster-12747.5831、Cluster-12747.16617等5個參與淀粉和蔗糖代謝合成途徑的關鍵酶基因,qPCR檢測表達水平與轉錄組測序結果一致。
關鍵詞:火龍果;果肉;轉錄組測序;淀粉;蔗糖
中圖分類號:S667.9????? 文獻標識碼:A
Differential Expression Analysis of Genes Related to Starch and Sucrose Metabolism Pathway in Different Developmental Stages of Dragon Fruit Pulp Based on Transcriptome
YANG Yunliang, LI Jianxun*, MA Genong
Cotton Research Institute, Shanxi Agricultural University, Yuncheng, Shanxi 044000, China
Abstract: In this study, Illumina Novaseq 6000 sequencing was used to sequence the samples from three stages of dragon fruit pulp (young fruit stage, color conversion stage and mature stage). The unigenes obtained was annotated in seven databases. A total of 3617 unigenes were successfully annotated in all databases. Five enzyme genes related to starch and sucrose metabolism pathway were further screened and verified by quantitative Real-time PCR (qPCR). The results showed that 115 193 transcripts and 47 855 transcripts were obtained by transcriptome sequencing and unigene N50 was 2000. The results of GO functional enrichment analysis showed that 18 582 unigenes were divided into three categories: biological process, cell composition and molecular function in GO functional annotation. The results of KOG functional classification showed that 6203 unigenes were divided into 25 group, in KOG annotation, of which post-translational modification, protein turnover and chaperone contained the most unigenes. The results of KEGG metabolic pathway annotation showed that 6554 unigenes got functional annotation, and there were 119 metabolic pathways, among which carbohydrate metabolism accounted for the highest proportion. Five key enzyme genes involved in starch and sucrose metabolism and synthesis pathway, such as UGP2, glgC, glgA, Cluster-12747.5831 and Cluster-12747.16617, were screened. The expression level of qPCR was consistent with that of transcriptome sequencing.
Keywords: dragon fruit; pulp; transcriptome sequencing; starch; sucrose
DOI: 10.3969/j.issn.1000-2561.2021.06.003
火龍果營養豐富,富含糖、有機酸、蛋白質、礦質元素、維生素、膳食纖維等[1-2],是我國南方重要的果樹樹種。近年來,隨著火龍果產業迅速發展,用于栽培的火龍果良種較少,通過分子生物學手段進行品種遺傳改良研究具有必要性。截至目前,火龍果的全基因組測序尚未完成,遺傳信息還不清楚,與果實發育相關的基因種類和功能尚不明確,這些均阻礙了火龍果果實品質形成機理等相關基礎研究的進行。淀粉和蔗糖是果實內在品質形成的主要因子。目前,對火龍果果實發育過程中淀粉和蔗糖代謝途徑研究較少,參與淀粉和蔗糖代謝合成的相關酶及酶基因尚未見報道,淀粉和蔗糖代謝過程是如何進行催化反應還不清楚。探究火龍果果肉中淀粉和蔗糖合成代謝途徑的分子調控機制,對改善果實品質具有重要的意義。
RNA測序[3](RNA Sequencing,簡稱RNA- Seq)又稱轉綠組測序技術,具有靈敏度高、定量準確、可重復性高、檢測范圍廣、分析更可靠等特點,被廣泛應用。該技術具有獲取全部mRNA轉錄本的豐度信息,從中挖掘功能基因和未知基因,解析基因調控狀況,揭示不同生物學過程與基因表達之間關系等方面具有獨特的優勢[4-5]。近年來,火龍果在轉錄水平上的研究取得了一系列進展,主要集中在逆境(低溫[6]、干旱[7]、水淹[8]、鹽脅迫[9])、火龍果果肉顏色[10-11]轉錄調控、甜菜堿[12-13]生物合成相關基因表達、病蟲害等[14]方面,然而基于火龍果果肉不同發育時期轉錄組測序方面的研究尚未見報道。
本研究以3個不同發育時期的紅肉火龍果果肉為研究對象,進行轉錄組測序研究,采用生物信息學技術方法,對得到的Unigene進行分類和功能注釋,分析差異表達基因富集的代謝通路,對參與火龍果果實發育過程中淀粉和蔗糖代謝過程的差異表達的酶基因進行篩選及qPCR驗證,旨在探索火龍果果實發育過程中關于淀粉和蔗糖代謝過程中酶的催化情況及酶基因的表達規律,為進一步實現人為調控火龍果果實糖分積累、提升果實品質提供理論依據。
1? 材料與方法
1.1? 材料
研究材料采自山西農業大學(山西省農業科學院)棉花研究所南花農場的15株紅肉火龍果樹,樹齡為3 a,品種為‘蜜寶,生長狀況良好。通過前期對15株紅肉火龍果樹物候期觀察,于2019年7月15日開始采樣,每5 d采1次樣,共采5次樣,選取3個關鍵時期進行研究,即:2019年7月15日(幼果期,Y)、2019年7月26日(轉色期,C)、2019年7月31日(成熟期,M),每次分別在3株樹的相同部位采集果實,采樣完成后迅速返回實驗室,將3個果實逐一去皮,切片(從果實中部橫切約0.3 cm),并立即將果肉放入液氮中冷凍,在?80 ℃保存備用。
1.2? 總RNA提取及檢測、cDNA文庫構建和Illumina測序
1.2.1? 總RNA提取及檢測? 使用TRNzol-A* Reagent提取火龍果果肉中的總RNA。采用瓊脂糖凝膠電泳分析火龍果果肉中RNA完整性及是否存在DNA污染,NanoPhotometer spectrophotometer檢測RNA純度,Agilent 2100 bioanalyzer檢測RNA完整性,以備合格的火龍果果肉RNA進行轉錄組測序。
1.2.2? cDNA文庫構建、質檢及測序? 火龍果果肉RNA檢測合格后,構建cDNA文庫。純化后的雙鏈cDNA,經過末端修復、加A尾并連接測序接頭,用AMPure XP beads篩選250~300 bp左右的cDNA,進行PCR擴增并再次使用AMPure XP beads純化PCR產物,最終獲得文庫。使用Agilent 2100 bioanalyzer對文庫的insert size進行檢測。庫檢合格后,把不同文庫按照有效濃度及目標下機數據量的需求pooling后進行Illumina測序。
1.3? 測序數據分析
1.3.1? 數據質控? 為了保證數據分析的質量及可靠性,需要對原始數據進行過濾。主要包括去除帶接頭的reads、含N的reads、低質量reads。同時,對clean data進行Q20、Q30和GC含量計算。后續所有分析均是基于clean data進行高質量分析。
1.3.2? 轉錄本拼接、Corset層次聚類及轉錄本質量評估? 轉錄組數據使用Trinity[15]軟件對clean data進行從頭組裝獲得轉錄本。Corset[16]層次聚類,在Trinity軟件拼接基礎上,根據轉錄本間Shared Reads將轉錄本聚合為許多cluster,再結合不同樣本間的轉錄本表達水平及H-Cluster算法,將樣本間有表達差異的轉錄本從原cluster分離,建立新的cluster,最終每個cluster被定義為“Gene”。該方法可以聚合冗余轉錄本,并提高差異表達基因的檢出率。本研究采用BUSCO[17]軟件進行轉錄本質量評估。
1.3.3? 參考序列比對? 將Trinity拼接得到的轉錄組作為參考序列(Ref),將每個樣品的clean reads往Ref上做mapping,分別過濾掉比對質量值低于10的reads,非成對比對上的reads,比對到基因組多個區域的reads。比對采用RSEM[18]軟件,RSEM中使用到的bowtie2參數mismatch0 (bowtie2默認參數)。
1.3.4? 基因功能注釋? 基于Nr、Nt、Pfam、Swiss-Prot進行序列相似性檢索蛋白質功能進行注釋。利用GO、KEGG、KOG數據庫分別進行
分子功能及相關代謝通路的比對預測分析。
1.4? 差異基因富集分析
本研究采用GOseq[19]和KOBAS[20]軟件對差異基因進行GO功能富集分析和KEGG通路富集分析等。
1.5? 熒光定量PCR分析
根據qPCR引物設計原則,采用Beacon Designer 7.9軟件進行引物設計,設計各候選基因的qPCR引物(表1)。引物由魔獾生物科技(上海)有限公司合成。本研究以火龍果3個發育時期的果肉為材料,YLS8為內參基因[21],采用SYBR GREEN I檢測進行qPCR反應,利用2?CT計算各個發育時期果肉的基因相對表達量。反應總體系為10 μL,其中包括2×SYBR? Green Supermix 5 μL、Forward prime 0.5 μL、Reverse primer 0.5 μL、cDNA 1 μL、ddH2O 3 μL。反應程序為:95 ℃ 3 min;95 ℃ 10 s,60 ℃ 30 s,39個循環。擴增完畢后,以1 ℃/4 s的速率從60 ℃逐步遞增到95 ℃,獲得溶解曲線。反應過程中,每個樣品設置3個生物學重復。
2? 結果與分析
2.1? 火龍果果肉轉錄組測序數據評估統計及拼接
本研究以‘蜜寶火龍果3個不同果肉發育時期即幼果期、轉色期、成熟期為樣本進行轉錄組測序,共獲得508 793 664個clean reads,包含76.32 Gb的核苷酸序列,Q20、Q30堿基百分比平均值均不小于97.95%、94.02% (表2)。將clean reads進行拼接組裝,共獲得火龍果果肉Transcripts 115 193條,Unigene 47 855條(表3)。Transcripts的N50和N90分別是2164和682,Genes的N50和N90分別是2000和508(表4)。結果表明,火龍果果肉測序數據質量高,可進行后續的生物信息學分析。
2.2? 火龍果3個不同果肉發育時期差異表達基因比較
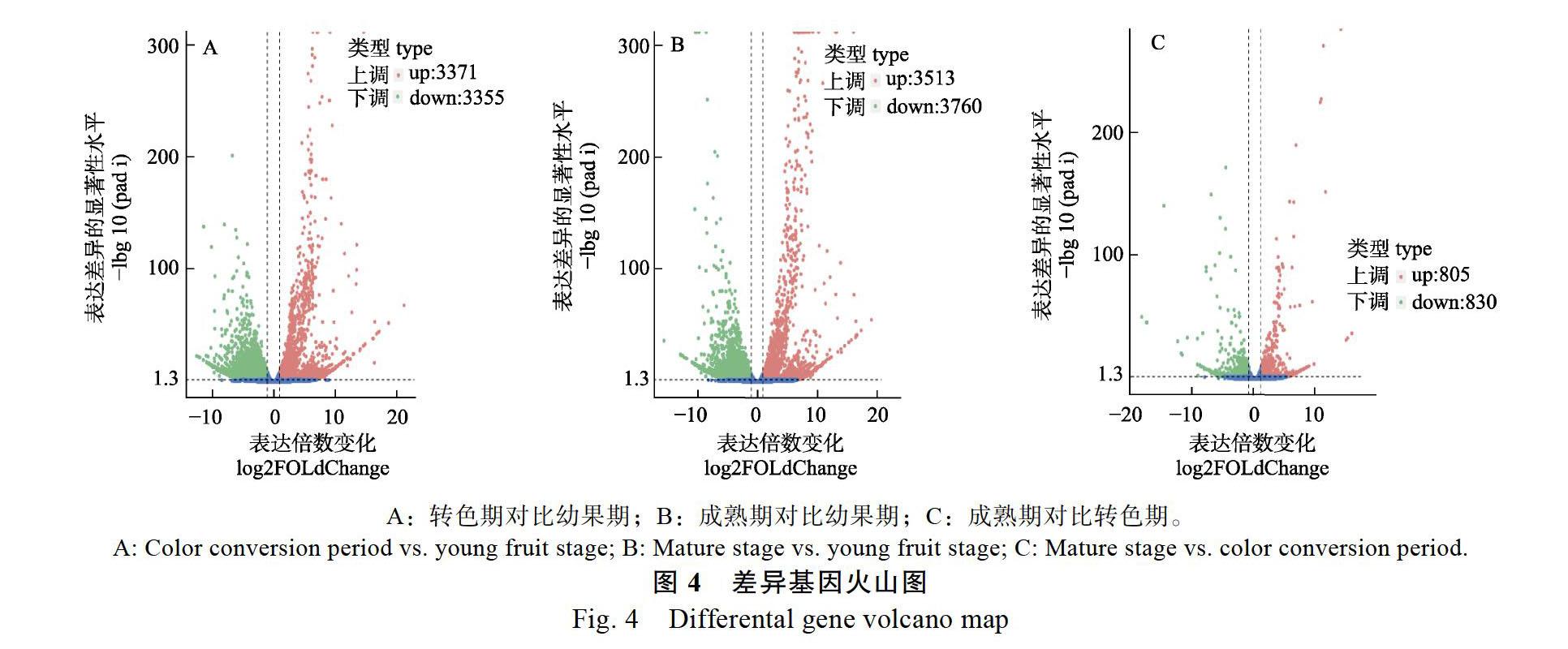
‘蜜寶火龍果3個不同果肉發育時期基因表達差異,兩兩比對,結果見表5。結果表明:轉色期與幼果期相比,有7226條Unigene差異基因表達,其中3371個基因上調表達,3855個基因下調表達;成熟期與幼果期相比,共有7273個差異表達基因,其中上調基因3513個,下調基因3760個;成熟期與轉色期相比,共有1635個差異表達基因,其中805個基因上調,830個基因下調。上述結果表明,火龍果果肉在成熟期的差異表達基因所占比例高于其他2個時期,說明DEGs在火龍果成熟期發育階段起著非常重要的作用。
2.3? 基因注釋和功能分類
2.3.1? 基因注釋? 獲得轉錄本后,需對轉錄本進行功能注釋,以獲得gene的功能信息。為了解火龍果果肉轉錄本的注釋情況,與7大數據庫進行比對,共有47 855個Unigene成功注釋(表6),由于火龍果的基因組信息尚未公布,缺少參考基因組信息,部分序列未能注釋。與NR數據庫進行比對獲得的基因注釋信息最高,占全部注釋序列的49.2%。與PFAM和GO數據庫比對獲得的基因注釋信息次之,占全部注釋序列均為38.82%。與KOG數據庫比對獲得的基因注釋信息最少,占全部注釋序列的12.96%。
2.3.2? 火龍果果肉發育過程中差異表達基因GO功能富集? ‘蜜寶火龍果3個不同果肉發育時期差異表達基因經兩兩比對,18 582個Unigene在GO功能注釋中被分為3大類,且每一類注釋種類和數量各不相同。由圖1可知,生物過程主要集中在細胞過程、代謝過程和單一有機體過程。在生物過程中,多數差異基因主要參與到細胞過程、代謝過程、單一生物過程;細胞組件中,多數差異基因主要參與了細胞、細胞組成、細胞器、細胞器組成、大分子復合物、膜、膜組成;分子功能中,多數差異基因富集到綁定、催化活性。
2.3.3? 火龍果果肉發育過程中差異表達基因KOG注釋分類統計? 對火龍果果肉發育過程中差異表達基因進行KOG數據庫比對分析,6203個Unigene在KOG注釋中被分為25個group,結果如圖2所示。翻譯后修飾、蛋白質周轉、伴侶包含的Unigene最多,共854個;其次是一般功能預測,Unigene 803個;翻譯,核糖體結構與生物發生,Unigene 538個;細胞內運輸,分泌和囊泡運輸,Unigene 483個;RNA加工和修飾,Unigene 450個;未知功能,Unigene 426個;信號轉導機制,Unigene 420個;能量生產和轉換,Unigene 352個;轉錄,Unigene 351個;脂質轉運與代謝,Unigene 307個;碳水化合物轉運與代謝,Unigene 298個;氨基酸轉運與代謝,Unigene 289個;最少的是細胞運動性,注釋到2個Unigene。可見,在火龍果果肉發育過程中,除了轉錄、翻譯、修飾、運輸等最基本的生命活動外,信號轉導,脂質、碳水化合物及氨基酸轉運與代謝占重要地位,說明信號轉導,脂質、碳水化合物及氨基酸轉運在火龍果果肉發育過程中起到重要作用。
2.4? 火龍果果肉發育過程中差異表達基因的KEGG分類
對基因做KO注釋后,根據它們參與的KEGG代謝通路進行分類(圖3),共分為5類,分別為:細胞過程、環境信息處理、遺傳信息處理、代謝及有機系統,其中代謝占的比例最大。在細胞過程中,以運輸和分解代謝為主;在環境信息處理中,以信號轉導所占比例最高,膜運輸所占比例最小;遺傳信息處理中,以翻譯所占比例最高,復制和修復所占比例最小;代謝中,以碳水化合物代謝所占比列最高,多糖生物合成所占比例最小;在有機系統中,以環境適應為主。
2.5? 火龍果3個不同果肉發育時期差異基因(DEGs)富集的代謝通路分析
KEGG是有關Pathway的主要公共數據庫,通過Pathway顯著性富集,能夠確定火龍果果肉在3個不同發育時期的差異表達基因參與的主要代謝途徑。
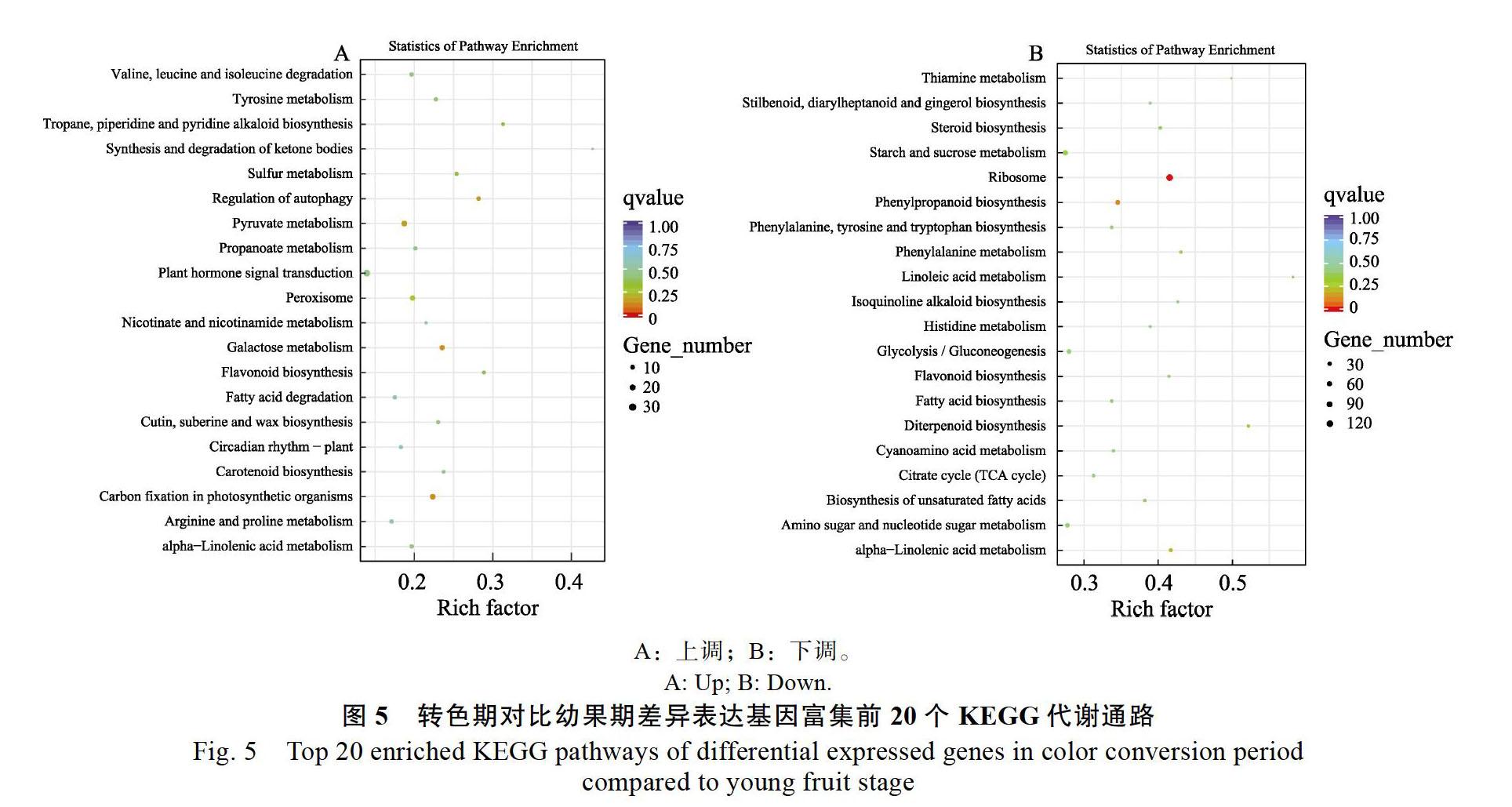
2.5.1? 轉色期對比幼果期? 轉色期與幼果期共有7226個差異基因(padj<0.05|&log2FoldChange|> 1),其中上調基因3371個,下調基因3855個(圖4A)。轉色期對比幼果期的差異基因共參與到119種代謝通路,其中富集于前20的KEGG代謝通路見圖5。轉色期上調的差異基因主要富集在植物激素信號轉導、丙酮酸代謝、光合生物的固碳作用、半乳糖代謝、過氧化物酶體、類黃酮生物合成、類胡蘿卜素生物合成中,其中與類胡蘿卜素生物合成相關的酶基因表達量均上調,包括胡蘿卜素去飽和酶、玉米黃質環氧化酶、9-順-環氧羰基雙加氧酶、花椒毒素脫氫酶,推測轉色期果肉顏色形成與類胡蘿卜素生物合成有關(圖5A)。轉色期下調的差異基因主要富集在核糖體、淀粉和蔗糖代謝、苯丙素類生物合成、糖酵解/糖異生、氨基糖和核苷酸代謝、α-亞麻酸代謝、檸檬酸循環、脂肪酸生物合成、苯丙氨酸、酪氨酸和色氨酸生物合成等(圖5B)。
2.5.2? 成熟期對比幼果期? 成熟期與幼果期共有7273個差異基因(padj<0.05|&log2FoldChange|> 1),其中上調基因3513個,下調基因3760個(圖4B)。成熟期對比幼果期的差異基因共參與到119種代謝通路,其中富集于前20的KEGG代謝通路見圖6。成熟期上調的差異基因主要富集在內質網中的蛋白質加工、淀粉和蔗糖代謝、剪接體、丙酮酸代謝、光合生物的固碳作用等(圖6A)。成熟期下調的差異基因主要富集在核糖體、植物激素信號轉導、淀粉和蔗糖代謝、苯丙素類生物合成、糖酵解/糖異生等。隨著火龍果果實的生長發育,成熟期較幼果期的果實口感甜,這與成熟期上調的差異基因富集在淀粉和蔗糖代謝、光合生物的固碳作用、半乳糖代謝有關(圖6B)。
2.5.3? 成熟期對比轉色期? 成熟期與轉色期共有1635個差異基因(padj<0.05|&log2FoldChange|> 1),其中上調基因805個,下調基因830個(圖4C)。成熟期對比轉色期的差異基因共參與到95種代謝通路,其中富集于前20的KEGG代謝通路見圖7。成熟期上調的差異基因主要富集在內質網中的蛋白質加工、苯丙素類生物合成、淀粉和蔗糖代謝、光合生物的固碳作用、半胱氨酸和蛋氨酸代謝、類黃酮生物合成、果糖和甘露糖代謝、甘氨酸、絲氨酸和蘇氨酸代謝、花青素生物合成等(圖7A)。成熟期下調的差異基因主要富集在內質網中的蛋白質加工、植物激素信號轉導、半乳糖代謝、糖酵解/糖異生、光合生物的固碳作用等(圖7B)。
2.6? 火龍果果肉中淀粉和蔗糖代謝途徑相關酶的基因統計及表達模式
2.6.1? 火龍果果肉中淀粉和蔗糖代謝途徑相關的差異基因統計? 本研究根據火龍果在3個不同果肉發育時期基因表達水平的顯著差異,統計了參與淀粉和蔗糖代謝途徑相關的酶基因個數。結果表明:轉色期與幼果期比對,參與淀粉和蔗糖代謝途徑的差異表達基因共有80個,其中上調基因25個,下調基因55個,下調基因占69%,成熟期與幼果期比對,參與淀粉和蔗糖代謝途徑的差異表達基因共有76個,其中上調基因28個,下調基因48個,下調基因占63%;成熟期與轉色期比對,參與淀粉和蔗糖代謝途徑的差異表達基因共有15個,其中上調基因7個,下調基因8個,下調基因占53%;通過對火龍果3個不同果肉發育時期差異基因比對整體來看,淀粉和蔗糖途徑為下調基因富集的通路。
2.6.2? 火龍果果肉中淀粉和蔗糖代謝途徑相關的酶基因的表達模式? 以DESeq2padj<0.05|log2?F?o?l?d?Change|>1為篩選條件,結合KEGG富集通路,在火龍果3個不同果肉發育時期篩選參與淀粉和蔗糖代謝途徑的差異表達酶基因5個(表7)。
2.7? qPCR驗證
為了驗證轉錄組測序數據的準確性,以YLS8為內參基因,篩選出參與淀粉和蔗糖代謝途徑的關鍵酶基因UGP2、glgC、glgA、Cluster- 12747.5831、Cluster-12747.16617等5個,并采用RT-PCR技術對5個關鍵的酶基因在火龍果3個不同果肉發育時期進行驗證分析。結果表明,RT-PCR驗證的5個與淀粉和蔗糖代謝途徑相關的酶基因在火龍果3個不同果肉發育時期上調或下調的變化倍數與轉錄組測序數據不一致,但基因表達量變化趨勢與轉錄組測序結果中基因的表達量趨勢基本一致(圖8)。即UGP2在幼果期表達量較高,隨著果肉發育表達量持續下降(圖8A);glgC在轉色期表達量較高,幼果期次之,成熟期表達量較低(圖8B);glgA在成熟期表達量較高,幼果期次之,轉色期表達量較低(圖8C);Cluster-12747.5831在幼果期表達量較高,轉色期表達量較低,成熟期較轉色期的表達量略有回升(圖8D);Cluster-12747.16617在幼果期和轉色期的表達量較高且一致,成熟期表達量最低(圖8E)。以上結果表明,轉錄組測序數據可靠性較高。同時發現,火龍果果肉中UGP2、glgC、glgA、Cluster-12747.5831、Cluster-12747.16617表達受到明顯的發育調控。
3? 討論
本研究針對火龍果3個不同時期的果肉組織進行轉錄組測序,共得到76.32 Gb測序數據,組裝后共得到115 193條轉錄本,對獲得的Unigene進行七大數據庫(NR、NT、KO、Swiss-prot、PFAM、GO、KOG)比對,共有47 855個Unigene成功注釋。KEGG代謝通路注釋結果表明,6554個Unigene獲得功能注釋,共有119條代謝途徑,其中碳水化合物代謝所占比例最高。
結合KEGG分析,關于淀粉和蔗糖代謝途徑相關酶發現,合成淀粉和蔗糖的代謝途徑主要2條,一條代謝途徑為:D-葡萄糖-1-磷酸在UTP-葡萄糖-1-磷酸尿苷酰轉移酶(UGP2)催化作用下,生成UDP-葡萄糖,UDP-葡萄糖在糖原合酶(GYS)和顆粒結合型淀粉合成酶(WAXY)的共同催化作用下,生成直鏈淀粉,直鏈淀粉在1,4-α-葡聚糖分支酶(GBE1)催化作用下生成淀粉;另一條代謝途徑為:D-葡萄糖-1-磷酸在葡萄糖-1-磷酸腺苷酸轉移酶(glgC)催化作用下,生成ADP-葡萄糖,ADP-葡萄糖在淀粉合酶(glgA)和顆粒結合型淀粉合成酶(WAXY)共同催化作用下,生成直鏈淀粉,直鏈淀粉在1,4-α-葡聚糖分支酶(GBE1)催化作用下生成淀粉。
本試驗篩選出了5個參與淀粉和蔗糖代謝途徑的酶基因。其中,UTP-葡萄糖-1-磷酸尿苷酰轉移酶(UGP2)在一條淀粉和蔗糖代謝途徑的早期階段發揮作用,以D-葡萄糖-1-磷酸為底物,催化形成UDP-葡萄糖。UGP2的表達影響著UGP2的活性及UDP-葡萄糖生成。本研究結果顯示UGP2在火龍果果肉幼果期的表達量最高,隨著果肉發育表達量持續下降。推測在火龍果果肉幼果期生成的UDP-葡萄糖最多,隨著果肉的不斷發育成熟,生成的UDP-葡萄糖逐漸減少,成熟期最少。葡萄糖-1-磷酸腺苷酸轉移酶(glgC)在另一條淀粉和蔗糖代謝途徑的早期階段發揮作用,催化D-葡萄糖-1-磷酸生成ADP-葡萄糖。glgC的表達影響著glgC的活性及ADP-葡萄糖的生成。本研究中,glgC在火龍果果肉轉色期的表達量最高,幼果期的表達量次之,成熟期的表達量較幼果期的略低。推測ADP-葡萄糖在火龍果果肉轉色期生成最多,幼果期次之,成熟期ADP-葡萄糖的生成略低于幼果期ADP-葡萄糖的生成。
淀粉合酶(glgA)在淀粉合成中起關鍵作用。glgA是一種轉糖基酶,在馬鈴薯[22]、荸薺[23]、小麥[24]等上均有研究,但在果樹上研究的較少。在火龍果果肉淀粉和蔗糖代謝過程中,ADP-葡萄糖在glgA的催化下,生成直鏈淀粉,直鏈淀粉在1,4-α-葡聚糖分支酶(GBE1)催化作用下生成淀粉。glgA的表達影響著glgA的活性及淀粉的合成。本研究表明,在火龍果果肉成熟過程中,glgA的表達量總趨勢是先降低后升高,在成熟期時表達量達到高峰。說明在火龍果3個不同果肉發育期均有淀粉的合成,其中以成熟期合成淀粉最多。
β-葡萄糖苷酶是一種水解酶,為纖維素降解過程中的限速酶[25]。在火龍果果肉淀粉和蔗糖代謝過程中,β-葡萄糖苷酶主要催化纖維糊精生成纖維二糖,纖維二糖再次在β-葡萄糖苷酶的催化作用下生成葡萄糖;另外,β-葡萄糖苷酶還可催化β-D-葡萄糖苷生成葡萄糖。Cluster-12747.5831的表達影響著β-葡萄糖苷酶的活性及葡萄糖的合成。本研究表明,Cluster-12747.5831在幼果期的表達量最高,轉色期的表達量降至最低,成熟期表達量高于轉色期的表達量。這與段朝瑞等[26]以玫瑰香葡萄果肉為研究對象,在其不同成熟時期的VvBG1(β-葡萄糖苷酶基因)的表達趨勢相似。
蔗糖合酶具有分解和合成蔗糖的作用[27],既可催化蔗糖合成又可催化蔗糖分解,是植物蔗糖代謝的關鍵酶。在火龍果果肉淀粉和蔗糖代謝途徑中,蔗糖在蔗糖合酶的分解作用下生成UDP-葡萄糖、D-果糖;UDP-葡萄糖在蔗糖合酶的合成作用下生成蔗糖。Cluster-12747.16617的表達影響著蔗糖合酶的活性及蔗糖的分解和合成。張春華等[28]研究桃果實不同發育時期蔗糖合成酶基因表達情況,結果表明:花后45~65 d,蔗糖合成酶基因表達量略微提高,85 d時,表達量下降至最低,105 d時,表達量迅速提高。本研究表明,Cluster-12747.16617在火龍果果肉幼果期和轉色期的表達量最高且一致,成熟期降低。這可能是由于不同種或火龍果為非躍變型果實的的原因。推測幼果期和轉色期的果肉中以蔗糖合成為主,成熟期的果肉中以分解蔗糖為主。
綜上所述,在火龍果果肉淀粉和蔗糖代謝途徑中,5個基因在3個果肉發育時期的表達情況各不相同。UGP2、glgC在淀粉和蔗糖代謝途徑的早期階段發揮作用,為淀粉和蔗糖的合成提供中間產物;glgA調控淀粉的合成;Cluster-12747. 5831調控葡萄糖的合成;Cluster-12747.16617調控蔗糖的分解和合成。
參考文獻
[65]??? 王? 壯, 王立娟, 蔡永強, 等. 火龍果營養成分及功能性物質研究進展[J]. 中國南方果樹, 2014, 43(5): 25-29.
[66]??? 蔡永強, 向青云, 陳家龍, 等. 火龍果的營養成分分析[J]. 經濟林研究, 2008, 26(4): 53-56.
[67]??? 吳小梅, 張? 昕, 李南羿. 雙孢蘑菇子實體不同發育時期的轉錄組分析[J]. 菌物學報, 2017, 36(2): 193-203.
[68]??? 涂美艷, 李明章, 孫淑霞, 等. 紅肉獼猴桃果實發育期不同果肉部位比較轉錄組分析[J]. 分子植物育種, 2019, 17(3): 729-738.
[69]??? 張? 雪, 唐銘浩, 陳? 蒙, 等. 山葡萄不同著色時期果皮轉錄組測序分析[J]. 果樹學報, 2017, 34(7): 781-789.
[70]??? 陸? 涵. 低溫脅迫下火龍果生理響應特征及轉錄組差異基因分析[D]. 成都: 四川農業大學, 2017.
[71]??? 齊? 釗, 閆? 臻, 徐? 敏, 等. 火龍果干旱脅迫下根和莖的基因差異表達[J]. 熱帶生物學報, 2018, 9(3): 320-327.
[72]??? 閆? 臻, 齊? 釗, 熊? 睿, 等. 火龍果根系在淹水脅迫下的基因差異表達[J]. 熱帶生物學報, 2018, 9(3): 312-319.
[73]??? Nong Q D, Zhang M Y, Chen J T, et al. RNA-seq de novo assembly of red pitaya (Hylocereus polyrhizus) roots and differential transcriptome analysis in response to salt stress[J]. Tropical Plant Biology, 2019, 12(2): 55-66.
[74]??? 曾燦彬, 徐? 敏, 齊? 釗, 等. 火龍果果肉顏色相關MYB轉錄因子的表達分析[J]. 熱帶生物學報, 2017, 8(2): 153-158.
[75]??? 吳亞維, 徐? 娟, 韓秀梅, 等. 紅白果肉顏色火龍果果肉轉錄組分析和基因功能注釋[J]. 分子植物育種, 2019, 17(2): 400-410.
[76]??? 鄭乾明, 王? 壯, 蔡永強, 等. 紅肉火龍果(Hylocereus polyrhizus)果實與莖轉錄組測序分析[J]. 分子植物育種, 2019, 17(1): 55-64.
[77]??? Xi X, Zong Y, Li S, et al. Transcriptome analysis clarified genes involved in betalain biosynthesis in the fruit of red pitayas (Hylocereus costaricensis)[J]. Molecules, 2019, 24(3): 445.
[78]??? Xu M, Liu C L, Luo J, et al. Transcriptomic de novo analysis of pitaya (Hylocereus polyrhizus) canker disease caused by
Neoscytalidium dimidiatum[J]. BMC Genomics, 2019, 20(1): 10.
[79]??? Grabherr M G, Haas B J, Yassour M, et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome[J]. Nature Biotechnology, 2011, 29(7): 644-652.
[80]??? Davidson N M, Oshlack A. Corset: enabling differential gene expression analysis for de novo assembled transcriptomes[J]. Genome Biology, 2014, 15(7): 410.
[81]??? Seppey M, Manni M, Zdobnov E M. BUSCO: assessing genome assembly and annotation completeness[J]. Methods in Molecular Biology, 2019, 1962: 227-245.
[82]??? Li B, Dewey C N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome[J]. BMC Bioinformatics, 2011, 12: 323.
[83]??? Young M D, Wakefield M J, Smyth G K, et al. Gene ontology analysis for RNA-seq: accounting for selection bias[J]. Genome Biol, 2010, 11(2): R14.
[84]??? Mao X, Cai T, Olyarchuk J G, et al. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary[J]. Bioinformatics, 2005, 21(19): 3787-3793.
[85]??? 崇慧影, 文曉鵬. 發育期火龍果內參基因的篩選和驗證[J]. 生物技術, 2019, 29(1): 39-45, 56.
[86]??? 石? 瑛, 王金明, 唐宏亮, 等. 馬鈴薯塊莖淀粉合成酶基因表達與淀粉積累[C]//中國作物學會馬鈴薯專業委員會, 馬鈴薯產業與健康消費. 哈爾濱: 黑龍江科學技術出版社, 2019: 342-343.
[87]??? 俞梅珍. 荸薺顆粒結合型淀粉合成酶基因的克隆與表達分析[D]. 揚州: 揚州大學, 2017.
[88]??? 譚彩霞. 小麥籽粒淀粉合成酶基因表達與淀粉合成的關系[D]. 揚州: 揚州大學, 2009.
[89]??? 王國澤, 方? 杰, 王志國, 等. 地稍瓜β-葡萄糖苷酶和多聚半乳糖醛酸酶基因克隆[J]. 內蒙古科技大學學報, 2019, 38(4): 327-330.
[90]??? 段朝瑞, 陳? 佩, 張國軍, 等. 葡萄成熟過程中生理特征與β-葡萄糖苷酶基因的表達分析[J]. 中國農業大學學報, 2013, 18(2): 50-55.
[91]??? 王永章, 蔣家慧, 李培環, 等. 肉質果實碳水化合物代謝及調控研究進展[J]. 萊陽農學院學報, 2001, 18(4): 288-296, 279.
[92]??? 張春華, 俞明亮, 馬瑞娟, 等. 桃不同發育時期主要糖類含量和蔗糖合成酶基因表達水平的動態變化[J]. 江蘇農業學報, 2014, 30(6): 1456-1463.
責任編輯:黃東杰
