龍眼miRNA合成途徑相關(guān)基因的鑒定及表達(dá)分析
2021-08-04 07:31:38霍雯陳曉慧徐小萍申序李曉斐賴鐘雄
熱帶作物學(xué)報(bào) 2021年6期
霍雯 陳曉慧 徐小萍 申序 李曉斐 賴鐘雄

摘? 要:為了解龍眼miRNA合成途徑相關(guān)基因的生物學(xué)功能,本研究依據(jù)龍眼第3代基因組數(shù)據(jù)對(duì)龍眼miRNA合成途徑相關(guān)基因進(jìn)行了鑒定和生物信息學(xué)分析,并利用龍眼轉(zhuǎn)錄組對(duì)其在體胚發(fā)生早期定量表達(dá)進(jìn)行分析。研究結(jié)果如下:共鑒定到12條龍眼miRNA合成途徑相關(guān)基因,分別是DlDDL1、DlDDL2、DlDDL3、DlDCL1、DlSE、DlHYL1a、DlHYL1b、DlHYL1c、DlCBP20、DlCBP80、DlHEN1、DlHASTY。這些基因編碼的氨基酸數(shù)目400~1000 aa,蛋白分子質(zhì)量23.92~220.25 kDa;大部分蛋白均為酸性蛋白,所有蛋白均為不穩(wěn)定蛋白,除DlHASTY外,其余蛋白均為親水性蛋白;基因結(jié)構(gòu)分析表明,各成員內(nèi)含子數(shù)目差別較大,為2~23個(gè);所有成員均含有保守結(jié)構(gòu)域,數(shù)目差別較大,為1~7個(gè);啟動(dòng)子順式原件分析顯示,基因啟動(dòng)子區(qū)域均含有大量光反應(yīng)元件及激素響應(yīng)元件;蛋白互作預(yù)測(cè)分析顯示龍眼miRNA合成途徑相關(guān)基因具有較強(qiáng)的互作關(guān)系。對(duì)龍眼體胚發(fā)生早期過(guò)程中表達(dá)分析顯示,各基因在EC階段具有較高的轉(zhuǎn)錄水平。本研究表明:龍眼miRNA合成途徑相關(guān)基因可能參與龍眼體胚發(fā)育早期進(jìn)程且在EC階段發(fā)揮重要作用,同時(shí)響應(yīng)多種激素應(yīng)答及非生物脅迫,推測(cè)其在生物學(xué)功能方面具有多樣性和差異性。
關(guān)鍵詞:龍眼;miRNA合成途徑相關(guān)基因;生物信息學(xué)分析;qPCR分析
中圖分類號(hào):S667.2????? 文獻(xiàn)標(biāo)識(shí)碼:A
Identification and Expression Analysis of Genes Related to Longan miRNA Synthesis
HUO Wen, CHEN Xiaohui, XU Xiaoping, SHEN Xu, LI Xiaofei, LAI Zhongxiong
Institute of Horticultural Biotechnology, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China
Abstract: In order to understand the biological functions of genes related to the longan miRNA synthesis pathway, we identified and analyzed the miRNA synthesis pathway-related genes in the third-generation genomic data of longan. And longan transcriptomes in the early stage of somatic embryogenesis (SE) was used for miRNA synthesis pathway-related genes expression analysis. A total of 12 genes related to longan miRNA synthesis pathway were identified, namely DlDDL1, DlDDL2, DlDDL3, DlDCL1, DlSE, DlHYL1a, DlHYL1b, DlHYL1c, DlCBP20, DlCBP80, DlHEN1, DlHASTY. The number of amino acids encoded by these genes was mostly between 400-1000 aa, and the molecular weight of the protein was 23.92-220.25 kDa. Most proteins were acidic proteins, all the proteins were unstable proteins. Except DlHASTY which is hydrophobic protein, the others were hydrophilic proteins. Gene structure analysis showed that the number of introns of each member varied greatly from 2 to 23. All members contained conservative domains, and the number difference varied from 1 to 7. Analysis of cis-acting elements in the promoter regions of gens related to miRNA synthesis pathway showed that a large number of light-responsive elements and hormone-responsive elements were presented in the promoter region of these genes. Protein interaction prediction analysis showed that the longan miRNA synthesis pathway-related genes had strong interaction relationship. Analysis of expression during the early stage of SE in longan showed that each gene had a high level of transcription at the EC stage. This study indicates that the genes related to the longan miRNA synthesis pathway may participate in the early development of longan SE development and play an important role in the EC stage, as well as responding to multiple hormone responses and abiotic stresses. Moreover, it is suggested that miRNA synthesis pathway-related genes might have diversity and differences in biological functions.
Keywords: longan; miRNA synthesis pathway-related genes; bioinformatics analysis; qPCR analysis
DOI: 10.3969/j.issn.1000-2561.2021.06.001
MicroRNA(miRNA)是一類長(zhǎng)度約為21~24 nt的內(nèi)源性非編碼的單鏈負(fù)調(diào)控因子,主要在轉(zhuǎn)錄水平上通過(guò)介導(dǎo)基因的甲基化或在轉(zhuǎn)錄后水平上通過(guò)介導(dǎo)靶基因的降解等調(diào)節(jié)植物基因的表達(dá)。研究表明,在低等植物和高等植物中普遍存在很多miRNA家族,并且在進(jìn)化過(guò)程很保守,這些miRNA的功能幾乎涉及到生長(zhǎng)發(fā)育、信號(hào)轉(zhuǎn)導(dǎo)以及環(huán)境響應(yīng)等生命進(jìn)程的各個(gè)方面[1-3]。miRNA在植物體細(xì)胞胚發(fā)生過(guò)程中同樣起著很重要的調(diào)控作用,在玉米[4]、柑橘[5]、龍眼[6]等中均有相關(guān)研究。
植物miRNA的生物合成大致可以分為3個(gè)步驟,首先在細(xì)胞核內(nèi)由RNA聚合酶Ⅱ轉(zhuǎn)錄成初級(jí)轉(zhuǎn)錄本(pri-mi RNA),pri-miRNA是含有一個(gè)或多個(gè)非完全互補(bǔ)的發(fā)卡結(jié)構(gòu)[7];然后由DCLl (Dicer-like 1)切割pri-miRNA形成莖環(huán)狀前體miRNA(pre-miRNA),并在DCL1酶和RNA結(jié)合蛋白HYL1(Hyponastic Leavas 1)及C2H2鋅指蛋白SE(Serrate)的作用下形成miRNA:miRNA*雙鏈復(fù)合物[8];接著,由HUA ENHANCER 1(HEN1)甲基轉(zhuǎn)移酶對(duì)雙鏈結(jié)構(gòu)3'端堿基進(jìn)行甲基化,在HASTY蛋白協(xié)助下由細(xì)胞核運(yùn)至細(xì)胞質(zhì);成熟 miRNA的一條鏈在細(xì)胞質(zhì)中被降解,另一條成熟的miRNA進(jìn)入RNA誘導(dǎo)的基因沉默復(fù)合物RISC(RNA-induced silencing complex)中發(fā)揮作用[9]。越來(lái)越多研究表明miRNA生物合成過(guò)程除了需要以上蛋白參與外,還需要許多蛋白因子的協(xié)同作用來(lái)共同調(diào)控miRNA的產(chǎn)生,SE蛋白與帽子蛋白CBP20(cap binding protein 20)和CDP80(cap binding protein 80)在pri- miRNA剪切過(guò)程中相互作用發(fā)揮著重要功能;在擬南芥中除DCL1、HYL1、SE外還發(fā)現(xiàn)了一種DDL(dawdle)蛋白也參與miRNA合成過(guò)程,在缺少DDL的突變體中發(fā)現(xiàn)pri-miRNA和成熟miRNA的積累減少[10]。
龍眼是我國(guó)重要的熱帶木本植物,其果實(shí)大小、產(chǎn)量及品質(zhì)與胚胎發(fā)育過(guò)程有著密切的聯(lián)系[11]。本實(shí)驗(yàn)室已經(jīng)對(duì)龍眼體胚發(fā)育內(nèi)在分子及基因調(diào)控機(jī)制開(kāi)展了大量的研究,發(fā)現(xiàn)miRNA在龍眼體胚發(fā)生中具有重要作用[12-13],但關(guān)于龍眼miRNA合成途徑基因的研究還未見(jiàn)報(bào)道。因此本實(shí)驗(yàn)通過(guò)對(duì)龍眼miRNA合成途徑基因的鑒定及其理化性質(zhì)、染色體分布、基因結(jié)構(gòu)、蛋白保守結(jié)構(gòu)域、系統(tǒng)進(jìn)化樹(shù)、啟動(dòng)子中順式作用元件進(jìn)行分析,以及各基因在體胚發(fā)生早期不同階段特異表達(dá)FPKM分析,旨在了解龍眼miRNA合成途徑相關(guān)基因可能存在的生物學(xué)功能,為龍眼體胚發(fā)生過(guò)程的分子機(jī)制提供理論依據(jù)。
1? 材料與方法
1.1? 材料
采用在本實(shí)驗(yàn)室已發(fā)表的第2代基因組數(shù)據(jù)基礎(chǔ)上重新進(jìn)行3代測(cè)序精確組裝的龍眼基因組及轉(zhuǎn)錄組數(shù)據(jù)庫(kù)(待發(fā)表),第3代龍眼基因組測(cè)序平臺(tái)為PacBio Sequel,最終確定龍眼基因組染色體長(zhǎng)度為0.45 Gb,Contigs N50=4.4 Mb,Super scaffold N50=27.7 Mb,Contig長(zhǎng)度錨定率91.69%,Contig數(shù)量錨定率38.61%。擬南芥序列下載于TAIR-Home Page(https://www.arabidopsis. org/),其余物種如水稻、甜橙、楊樹(shù)等氨基酸序列及基因序列下載于NCBI(https://www.ncbi.nlm.nih.gov/)。
實(shí)驗(yàn)材料為福建農(nóng)林大學(xué)園藝植物生物工程研究所誘導(dǎo)并長(zhǎng)期繼代的龍眼品種‘紅核子松散型胚性愈傷組織(LC2細(xì)胞系),并參照賴鐘雄的培養(yǎng)方法獲得胚性愈傷組織(embrogenic cultures,EC),不完全胚性緊實(shí)結(jié)構(gòu)ICpEC(incomplete compact pro-embrogenic cultures,ICpEC),球形胚(globular embryos,GE)3個(gè)階段的胚性培養(yǎng)物,提取RNA后進(jìn)行反轉(zhuǎn)錄用于熒光定量PCR反應(yīng)[14]。
1.2? 方法
1.2.1? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因的鑒定及蛋白理化性質(zhì)分析? 在TAIR網(wǎng)站下載模式植物擬南芥miRNA合成途徑相關(guān)基因的氨基酸序列AtDDL(AT3G20550.1)、AtDCL1(AT1G 01040.1)、AtSE(AT2G27100.1)、AtHYL1(AT1G 09700.1)、AtCBP20(AT5G44200.1)、AtCBP80(AT2G13540.1)、AtHEN1(AT4G20910.1)、AtHASTY(AT3G05040.1)為種子序列,經(jīng)與龍眼基因組數(shù)據(jù)庫(kù)同源比對(duì),篩選獲得具有完整Open Reading Frame(ORF)的龍眼miRNA合成途徑相關(guān)基因的候選序列,根據(jù)基因組注釋,采用DNAMAN 6.0軟件對(duì)候選的基因組DNA序列(genomic DNA,gDNA)、編碼序列(coding sequence)CDS進(jìn)行分析和對(duì)比,結(jié)合NCBI Blast同源比對(duì)分析,然后根據(jù)在線網(wǎng)站Pfam(https:// pfam.xfam.org/)、NCBI Conserved Domain Search(https//www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb. cgi)以及HMMER (https://www.ebi.ac.uk/Tools/ hmm)對(duì)龍眼miRNA合成途徑相關(guān)候選基因的蛋白結(jié)構(gòu)域進(jìn)行篩選,最終確定了12條相關(guān)基因。
根據(jù)龍眼基因組中的查找注釋,參考模式植物擬南芥和水稻的命名方法對(duì)得到的12條龍眼基因進(jìn)行命名。通過(guò)ExPASy(https://web.expasy. org/protparam)在線網(wǎng)站對(duì)上述12條基因的蛋白理化性質(zhì)如氨基酸數(shù)目、相對(duì)分子量、等電點(diǎn)、不穩(wěn)定系數(shù)以及親水性指數(shù)等方面進(jìn)行分析。
1.2.2? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因結(jié)構(gòu)與蛋白結(jié)構(gòu)域分析? 采用工具TBtools[15]并利用龍眼gff及genome文件對(duì)獲得的12條基因序列進(jìn)行外顯子和內(nèi)含子特征分析;利用Pfam、CDD、HMMER等在線網(wǎng)站進(jìn)行基因蛋白結(jié)構(gòu)域的預(yù)測(cè),并通過(guò)TBtools進(jìn)行圖片可視化繪制。
1.2.3? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因進(jìn)化樹(shù)構(gòu)建? 通過(guò)NCBI下載其他物種相關(guān)基因的蛋白序列,最后采用MEGA軟件中的鄰近法(Neighbor-joining method)對(duì)龍眼及下載的蛋白序列進(jìn)行系統(tǒng)進(jìn)化樹(shù)構(gòu)建,自展系數(shù)法(Bootstrap)設(shè)置為1000次重復(fù)檢驗(yàn)。
1.2.4? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因啟動(dòng)子分析? 經(jīng)龍眼基因組數(shù)據(jù)庫(kù)提取獲得12條基因ATG上游2000 bp的序列,采用在線網(wǎng)站PlantCARE(http://bioinformatics.psb.ugent.be/ webtools/plantcare/html/)預(yù)測(cè)分析其啟動(dòng)子特征和順勢(shì)作用元件功能,采用TBtools進(jìn)行圖片可視化繪制。
1.2.5? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因蛋白互作預(yù)測(cè)分析? 利用在線網(wǎng)站STRING(https://string-db.org)進(jìn)行蛋白質(zhì)互作預(yù)測(cè),選用研究最深入的擬南芥作為參考,探究龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因蛋白之間的互作情況。
1.2.6? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因不同階段特異性表達(dá)分析? 通過(guò)龍眼轉(zhuǎn)錄組數(shù)據(jù)庫(kù)提取miRNA合成途徑相關(guān)基因在不同體胚發(fā)生階段:EC,ICpEC和GE特異表達(dá)的FPKM值,利用TBtools繪制表達(dá)熱圖,分析各基因在不同階段的表達(dá)情況。
1.2.7? 實(shí)時(shí)熒光定量PCR(qRT-PCR)分析? 提取篩選得到的12條基因CDS序列,通過(guò)DANMAN結(jié)合引物設(shè)計(jì)原則設(shè)計(jì)12對(duì)符合條件的引物(表1);參照Lin等[16]的具體操作步驟,采用20 μL反應(yīng)體系進(jìn)行熒光定量PCR測(cè)定;通過(guò)羅氏LightCyclers480熒光定量PCR儀,以EF-1a為內(nèi)參基因,采用2-ΔΔCT計(jì)算龍眼miRNA合成途徑相關(guān)基因在體胚發(fā)生早期中的表達(dá)情況;利用Graphpad Prism 7進(jìn)行作圖,并與其FPKM值進(jìn)行分析比較。
2? 結(jié)果與分析
2.1? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因的鑒定及蛋白理化性質(zhì)分析
首先從TAIR網(wǎng)站上下載擬南芥miRNA合成途徑相關(guān)基因編碼蛋白的氨基酸序列:經(jīng)過(guò)龍眼第3代基因組數(shù)據(jù)庫(kù)的比對(duì)以及NCBI Blast雙向比對(duì)結(jié)果,再結(jié)合Pfam、HMMER、CDD等網(wǎng)站進(jìn)行結(jié)構(gòu)域預(yù)測(cè),最后鑒定出12條龍眼miRNA合成途徑相關(guān)基因。最后參照模式植物擬南芥、水稻的命名方式將其命名為DlDDL1、DlDDL2、DlDDL3、DlDCL1、DlSE、DlHYL1a、DlHYL1b、DlHYL1c、DlCBP20、DlCBP80、DlHEN1、DlHASTY。
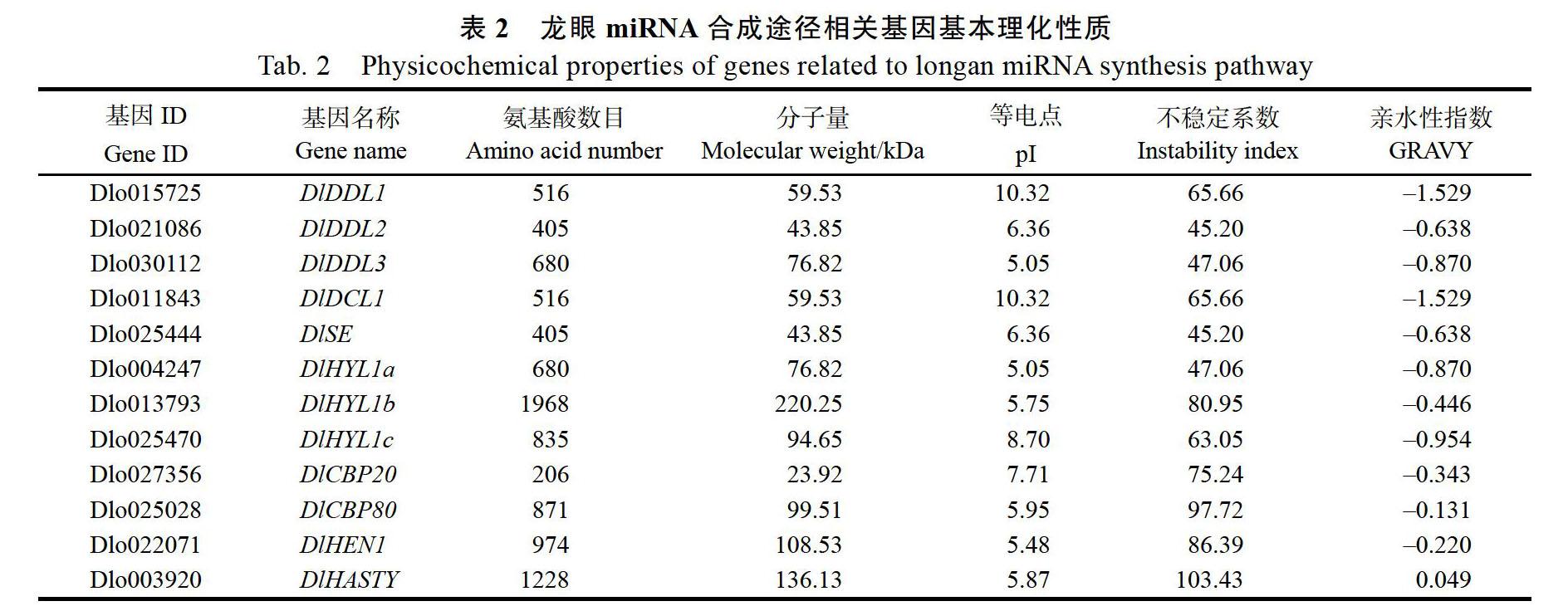
通過(guò)蛋白質(zhì)特性分析可以發(fā)現(xiàn)(表2),miRNA合成途徑相關(guān)基因編碼的蛋白在400~ 1000 aa之間變化,蛋白分子質(zhì)量23.92~220.25 kDa,其中DlHTL1b氨基酸數(shù)目最長(zhǎng)為1968 aa,DlCBP20最短為206 aa;等電點(diǎn)5.05~10.32,大部分為酸性蛋白;不穩(wěn)定系數(shù)45.20~103.43,均為不穩(wěn)定蛋白;DlHASTY親水性指數(shù)為0.049,是疏水性蛋白,而其余蛋白親水性指數(shù)為–1.529~ –0.131,為親水性蛋白。
2.2? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因結(jié)構(gòu)及蛋白結(jié)構(gòu)分析
為了進(jìn)一步了解龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因的結(jié)構(gòu)特征,對(duì)12條基因的外顯子-內(nèi)含子的結(jié)構(gòu)進(jìn)行研究,發(fā)現(xiàn)其表現(xiàn)出不同的外顯子-內(nèi)含子的組織模式(圖1)。基因長(zhǎng)度存在著明顯的差異,這很可能是因?yàn)樵诠δ苌系牟煌瑢?dǎo)致的。大部分基因的外顯子數(shù)量為10~20個(gè),DlDDL的3個(gè)成員中DlDDL2的外顯子數(shù)目最少,僅有2個(gè),DlDDL1和DlDDL3分別含有8和10個(gè)外顯子;DlDCL1、DlCBP80、DlHASTY的外顯子數(shù)目較多,均在20個(gè)左右,但DlCBP20的外顯子數(shù)目較少為7個(gè);DlHYL1的1a、1c兩個(gè)成員外顯子數(shù)目較為一致相差不大,1b的外顯子數(shù)量?jī)H為2個(gè)。12條基因呈現(xiàn)出不同的外顯子–內(nèi)含子數(shù)量分布,這可能與基因功能的分化有關(guān)。
蛋白保守結(jié)構(gòu)域的結(jié)果顯示(圖2):DlDDL三個(gè)成員均含有典型的FHA結(jié)構(gòu)域,但DlDDL1蛋白結(jié)構(gòu)域的位置和長(zhǎng)度均與DlDDL2、DlDDL3存在差異,推測(cè)它們?cè)诠δ苌峡赡苡兄煌姆止?DlDCL1顯示含有7個(gè)不同的蛋白結(jié)構(gòu)域,其中包括典型的Dicer與PAZ結(jié)構(gòu)域,且在不同植物中高度保守;DlSE顯示含有DUF3546和ARS2兩個(gè)結(jié)構(gòu)域;DlHYL1的3個(gè)成員中DlHYL1a和DlHYL1b均含有2個(gè)dsrm結(jié)構(gòu)域,DlHYL1c則含有3個(gè);DlCBP20和DlCBP80的結(jié)構(gòu)域不相同,DlCBP20只含有1個(gè)典型的RRM結(jié)構(gòu)域且長(zhǎng)度較長(zhǎng);DlHEN1顯示含有2個(gè)蛋白結(jié)構(gòu)域,分別為DSRM和baccter_Hen1;DlHASTY只含有1個(gè)蛋白結(jié)構(gòu)域Xpo1。
2.3? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因進(jìn)化樹(shù)構(gòu)建
為了進(jìn)一步了解龍眼miRNA合成途徑相關(guān)基因的進(jìn)化關(guān)系,在NCBI上下載不同物種相關(guān)基因編碼蛋白的氨基酸序列,并采用MEGA構(gòu)建系統(tǒng)發(fā)育進(jìn)化樹(shù)。結(jié)果顯示(圖3):DlDDL1與甜橙的DDL蛋白親緣關(guān)系最近,DlDDL2、DlDDL3分別與擬南芥和楊樹(shù)DDL蛋白親緣關(guān)系較近;DlDCL1與甜橙親緣關(guān)系最為接近;DlSE蛋白與木槿、擬南芥在進(jìn)化上保持較近的親緣關(guān)系,與甜橙、楊樹(shù)等親緣關(guān)系較遠(yuǎn);DlHYL1b、DlHYL1c與DlHYL1a不同,分別與擬南芥、油松親緣關(guān)系較近;DlCBP 20和DlCBP80在進(jìn)化關(guān)系上都與水稻在親緣關(guān)系上保持較遠(yuǎn)的距離;DlHEN1與橡膠樹(shù)、川桑親緣關(guān)系較近;DlHASTY與油松等親緣關(guān)系較遠(yuǎn)。
2.4? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因啟動(dòng)子分析
為進(jìn)一步了解龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因5端調(diào)控序列的功能,通過(guò)PlantCARE在線對(duì)12條基因起始密碼子ATG上游2000 bp序列進(jìn)行順式作用元件預(yù)測(cè),預(yù)測(cè)分析結(jié)果表明(圖4):12條基因均含有CAAT-box和TATA-box表明其都能夠進(jìn)行正常的轉(zhuǎn)錄,但各基因的CAAT-box和TATA-box的數(shù)量多少有明顯差異。對(duì)各基因含有的順式作用元件進(jìn)行分析發(fā)現(xiàn)(圖5),12條基因均含有大量的光學(xué)響應(yīng)元件及少量的厭氧誘導(dǎo)響應(yīng)元件,50%的基因都含有生長(zhǎng)素、乙烯、茉莉酸甲酯、水楊酸等激素響應(yīng)元件,75%的基因響應(yīng)脫落酸和赤霉素應(yīng)答;另外,還有一些基因含有與細(xì)胞周期、晝夜控制、干旱、脅迫、胚乳、分生組織、低溫等作用元件,提示這些基因可能參與生物鐘、抗旱抗寒、種子生長(zhǎng)、胚胎發(fā)育等過(guò)程。不同基因所含有的順式作用元件以及數(shù)量都有差異,推測(cè)其對(duì)不同的脅迫響應(yīng)具有一定的差異,并可能在龍眼生長(zhǎng)發(fā)育中承擔(dān)著不同的特異性功能。
2.5? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因蛋白互作預(yù)測(cè)分析
利用String蛋白互作在線數(shù)據(jù)庫(kù)對(duì)龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因蛋白之間的功能關(guān)系進(jìn)行預(yù)測(cè),選擇研究較為深入的模式植物擬南芥作為參考,蛋白互作預(yù)測(cè)結(jié)果分析顯示(圖6):miRNA合成途徑相關(guān)基因DCL1、SE、HASTY、DDL、HEN1、HYL1之間有很強(qiáng)的互作關(guān)系。
2.6? 龍眼體胚發(fā)生早期miRNA合成途徑相關(guān)基因不同階段FPKM值分析
為了進(jìn)一步了解miRNA合成途徑相關(guān)基因在龍眼體細(xì)胞胚胎發(fā)生中的功能特點(diǎn),結(jié)合龍眼第3代轉(zhuǎn)錄組數(shù)據(jù)庫(kù)中EC、ICpEC、GE 3個(gè)階段的FPKM值制作熱圖。結(jié)果顯示(圖7):在3個(gè)不同階段中,除DlHYL1c表達(dá)量很低,其他基因的表達(dá)量相對(duì)較高,其中DlCBP20的表達(dá)量最高。DlDDL在3個(gè)階段的表達(dá)量比較平穩(wěn),且DlDDL1的表達(dá)量較高;DlDCL1和DlSE在3個(gè)階段中呈現(xiàn)出先上調(diào)后下調(diào)的模式;DlHYL1a與DlHYL1b表達(dá)模式一致呈現(xiàn)出下調(diào)模式;DlCBP20與DlCBP80、DlHEN1呈現(xiàn)出下調(diào)模式;DlHASTY則呈現(xiàn)出先上調(diào)后下調(diào)的表達(dá)模式。總體上,miRNA合成途徑相關(guān)基因在龍眼體細(xì)胞早期發(fā)育過(guò)程中呈現(xiàn)出不同階段的表達(dá)模式,提示不同基因在不同的體胚發(fā)生階段有著不同的功能特點(diǎn)。
2.7? 龍眼體胚發(fā)生早期不同階段miRNA合成途徑相關(guān)基因的表達(dá)模式分析
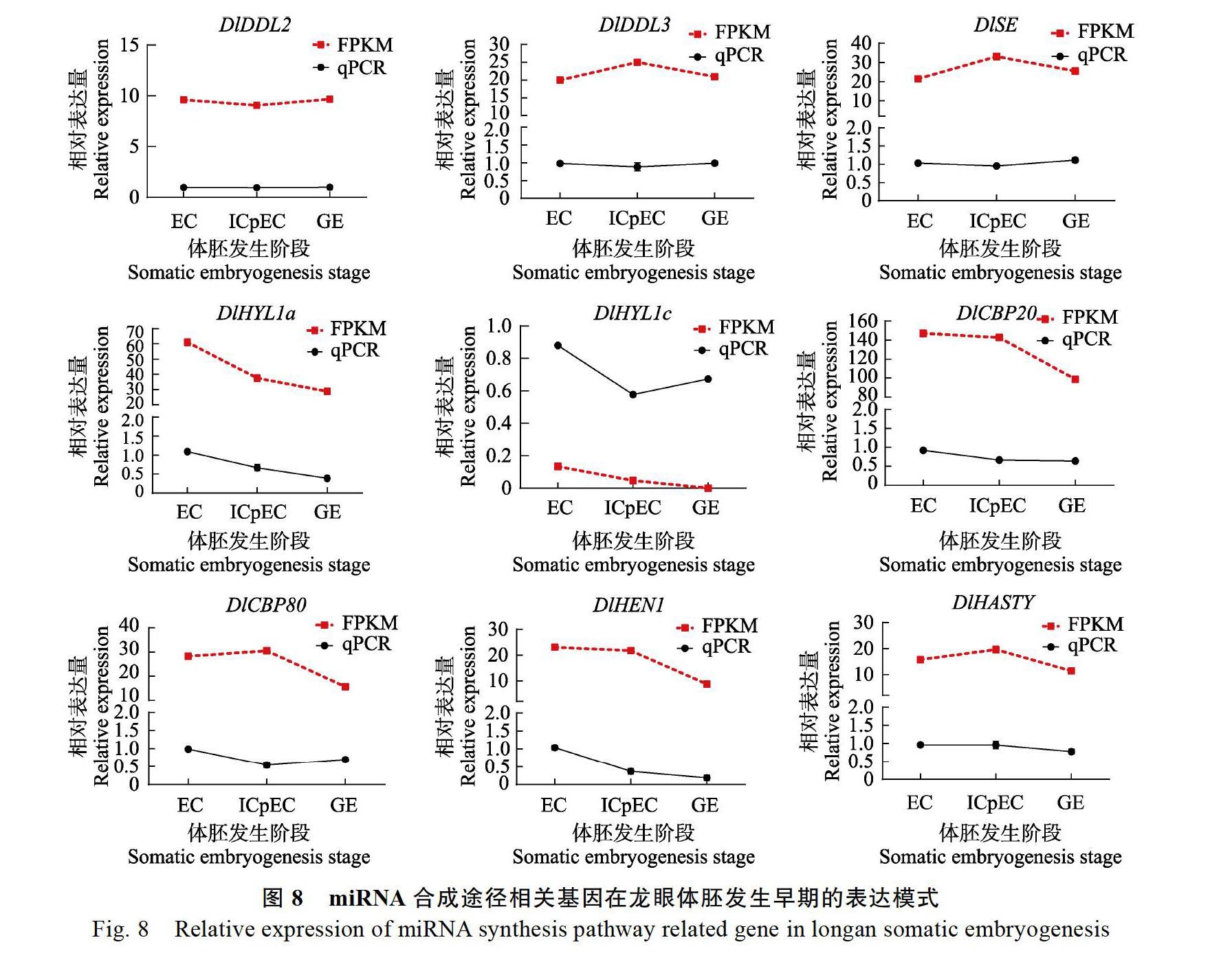
通過(guò)實(shí)時(shí)熒光定量PCR結(jié)果發(fā)現(xiàn)(圖8),DlDDL1與DlHYL1b幾乎不表達(dá),DlDDL2、DlDDL3在體胚早期3個(gè)階段表達(dá)量較平穩(wěn);DlSE在體胚早期3個(gè)階段的表達(dá)量也較平穩(wěn);DlHYL1a在3個(gè)階段的表達(dá)量呈現(xiàn)明顯的下降趨勢(shì),DlHYL1c在3個(gè)階段的相對(duì)表達(dá)量呈現(xiàn)先下降后上升的趨勢(shì),但總體上仍是呈現(xiàn)下降趨勢(shì);DlCBP20與DlCBP80的相對(duì)表達(dá)量總體上也是呈現(xiàn)下降趨勢(shì),DlHEN1與DlHASTY的相對(duì)表達(dá)量趨勢(shì)與其FPKM值的趨勢(shì)大致相同均呈現(xiàn)下降趨勢(shì)。由此可見(jiàn),miRNA合成途徑基因在龍眼體胚發(fā)生早期的表達(dá)程度有所不同,總體上在EC階段的表達(dá)量較高。
3? 討論
3.1? miRNA合成途徑相關(guān)基因在植物生長(zhǎng)發(fā)育中的作用
miRNA合成途徑涉及到很多基因的參與,這些基因已經(jīng)在模式植物尤其是擬南芥中展開(kāi)廣泛研究。AtDDLs在miRNA的生物合成和內(nèi)源性的siRNA中發(fā)揮作用,其突變體普遍呈現(xiàn)出發(fā)育過(guò)程延遲、生長(zhǎng)分化出不完全的根、幼苗和花、結(jié)實(shí)率降低等現(xiàn)象,充分表明了DDL蛋白在植物早期生長(zhǎng)發(fā)育過(guò)程中具有重要作用[17];HYL1又被稱為DRB1,是擬南芥中發(fā)現(xiàn)最早、研究最透徹的雙鏈RNA結(jié)合蛋白,擬南芥hyl1的突變體會(huì)出現(xiàn)如植株矮小、晚花、根系生長(zhǎng)慢、根向地性減弱等發(fā)育缺陷的癥狀,同時(shí)還會(huì)出現(xiàn)對(duì)IAA不敏感并且抑制生長(zhǎng)素運(yùn)輸?shù)默F(xiàn)象,因此HYL1在植物的生長(zhǎng)發(fā)育過(guò)程發(fā)揮著重要作用[18];有研究表明當(dāng)在體外共孵育DCL1、HYL1、SE蛋白及pri-miR167時(shí),能較精準(zhǔn)地剪切出成熟miR167序列,若只加HYL1或SE蛋白其中一種時(shí),剪切精確度會(huì)下降,若2種蛋白都不加時(shí),剪切精確度下降更明顯,這說(shuō)明HYL1和SE在DCL1識(shí)別pri-miRNA并剪切的過(guò)程中發(fā)揮重要作用[19];在水稻中發(fā)現(xiàn)CBP20蛋白可提高大腸桿菌重組對(duì)高鹽度、高熱和脫水的耐受性,提示OSCBP20蛋白在應(yīng)激條件下可能起保護(hù)作用[20],擬南芥中有研究發(fā)現(xiàn)CBP20和CBP80突變體中miRNA含量減少,pri-miRNA含量增多,免疫共沉淀實(shí)驗(yàn)表明,pri-miRNAs 159、166、168和172可能與CBP20和CBP80有關(guān),還發(fā)現(xiàn)ABA處理后的CBP20/80突變體種子中miR159積累減少,MYB33和MYB101轉(zhuǎn)錄水平的升高,推測(cè)CBP20和CBP80是ABA在種子萌發(fā)過(guò)程中誘導(dǎo)miR159所必需的[21];在擬南芥發(fā)育的大多數(shù)階段,HEN1的單個(gè)突變體表現(xiàn)出多種表型,如器官大小減少、蓮座葉形狀改變和花序數(shù)量增加[22];HASTY蛋白的缺失也會(huì)影響擬南芥發(fā)育中的許多過(guò)程,除減少莖根和側(cè)器官的大小外,還會(huì)影響莖尖分生組織的大小,延遲短時(shí)間內(nèi)的花誘導(dǎo),葉片和心皮的正面化,破壞花序的節(jié)性,降低生育能力等[23]。
3.2? miRNA合成途徑相關(guān)基因可能參與激素應(yīng)答和響應(yīng)非生物脅迫
miRNA合成途徑基因的表達(dá)不僅受到不同激素的誘導(dǎo)同樣也受到許多環(huán)境因子的調(diào)節(jié),不同的基因?qū)Ψ巧锩{迫的應(yīng)答模式也有所不同。對(duì)龍眼miRNA合成途徑相關(guān)基因啟動(dòng)子序列進(jìn)行生物信息學(xué)分析發(fā)現(xiàn),這些基因含有脫落酸、乙烯、赤霉素、生長(zhǎng)素、茉莉酸甲酯、水楊酸的激素響應(yīng)元件,脫落酸有調(diào)節(jié)胚的發(fā)育、加速脫落休眠及抑制生長(zhǎng)的生理功能[24],其中DlDCL1、DlCBP80、DlDRM2還有不止一個(gè)脫落酸激素元件;赤霉素不僅能促進(jìn)體胚發(fā)生相關(guān)的轉(zhuǎn)錄因子表達(dá)影響早期胚性組織細(xì)胞向球形胚階段的分化[25],還能影響胚軸的發(fā)育及種子的萌發(fā)[26];這些基因中還包含了水楊酸和茉莉酸甲酯的響應(yīng)元件,這2種激素在植物抗病機(jī)制中發(fā)揮重要作用;生長(zhǎng)素和乙烯對(duì)植物同樣有著很重要的作用,有研究發(fā)現(xiàn)這2種激素在棉花的體胚發(fā)育中有重要作用[27],因此推測(cè)其中一些基因在相應(yīng)的激素信號(hào)傳導(dǎo)中發(fā)揮一定作用。
3.3? miRNA及合成途徑相關(guān)基因參與植物體胚發(fā)育進(jìn)程
植物體胚發(fā)生過(guò)程中需要很多基因的共同參與,其中miRNA及相關(guān)基因?qū)χ参矬w細(xì)胞胚胎的形成也有影響。在擬南芥中通過(guò)原位雜交實(shí)驗(yàn)表明,HEN1可以通過(guò)miR167負(fù)調(diào)控ARF6和ARF8的表達(dá)進(jìn)而調(diào)控胚珠發(fā)育[28]。
本研究通過(guò)對(duì)miRNA合成途徑相關(guān)基因在龍眼體胚發(fā)生早期的FPKM值與基因表達(dá)模式分析發(fā)現(xiàn),DlDDL1與DlHYL1b在3個(gè)階段均不表達(dá),而DlHYL1c在3個(gè)階段均有表達(dá)。DlDDL2、DlHYL1a、DlCBP20、DlHEN1、DlHASTY的表達(dá)模式與其FPKM值趨勢(shì)一致,其中DlHYL1a、DlCBP20、DlHEN1均呈現(xiàn)出下調(diào)的模式,在EC階段的表達(dá)量最高,提示其可能在龍眼胚性愈傷組織建成中發(fā)揮重要作用,DlDDL2、DlHASTY在3個(gè)階段的表達(dá)量相差不大;DlHYL1c、DlCBP80、DlHEN1的表達(dá)模式與其FPKM值相比雖略有差異,但總體均呈現(xiàn)出下降趨勢(shì),即在EC階段的表達(dá)量最高;DlDDL3、DlSE的表達(dá)模式與其FPKM值相比也有差異,但在3個(gè)階段的表達(dá)趨勢(shì)均相對(duì)平穩(wěn)。DlDCL1在3個(gè)階段的表達(dá)量呈現(xiàn)明顯的下降趨勢(shì)[29]。龍眼miRNA合成途徑相關(guān)基因在體胚發(fā)生早期3個(gè)階段的表達(dá)模式有差異,提示這些基因可能在龍眼體胚發(fā)生早期過(guò)程中發(fā)揮著不同的作用。
參考文獻(xiàn)
[1]? Hutvagner G. A microRNA in a multiple-turnover RNAi enzyme complex[J]. Science, 297(5589): 2056-2060.
[2]? Zeng Y, Yi R, Cullen B R. MicroRNAs and small interfering RNAs can inhibit mRNA expression by similar mechanisms[J]. Proceedings of the National Academy of Sciences of the United States of America, 2003, 100(17): 9779-9784.
[3]? Doench J G, Petersen C P, Sharp P A. siRNAs can function as miRNAs[J]. Genes and Development, 2003, 17(4): 438-442.
[4]? 付? 蓉. 玉米自交系鹽脅迫響應(yīng)miRNA及其靶基因的鑒定研究[D]. 南通: 南通大學(xué), 2016.
[5]? 劉元龍. 參與柑橘果實(shí)發(fā)育的miRNA和靶基因的挖掘與分析[D]. 武漢: 華中農(nóng)業(yè)大學(xué), 2015.
[6]? 林玉玲, 賴鐘雄. 龍眼胚性愈傷組織miR398b前體克隆及其在龍眼體胚發(fā)生過(guò)程中的表達(dá)分析[J]. 熱帶作物學(xué)報(bào), 2012, 33(11): 2012-2017.
[7]? Lee Y, Kim M, Han J, et al. MicroRNA genes are transcribed by RNA polymerase II[J]. Embo Journal, 2004, 23 (20): 4051-4060.
[8]? Kurihara Y, Watanabe Y. Arabidopsis micro-RNA biogenesis through Dicer-like 1 protein functions[J]. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(34): 12753-12758.
[9] Yu B, Yang Z, Li J, et al. Methylation as a crucial step in plant microRNA biogenesis[J]. Science, 2005, 307(5711): 932-935.
[10]????? Yu B, Bi L, Zheng B, et al. The FHA domain proteins DAWDLE in Arabidopsis and SNIP1 in humans act in small RNA biogenesis[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(29): 10073-10078.
[11]????? 李冬梅. 龍眼體細(xì)胞胚胎成熟機(jī)理的研究[D]. 福州: 福建農(nóng)林大學(xué), 2003: 3-5.
[12]????? 林玉玲, 賴鐘雄. 利用Solexa測(cè)序技術(shù)鑒定龍眼體胚發(fā)生過(guò)程中的miRNAs[EB/OL].(2011-02-10) [2020-05-25]. http://www.paper.edu.cn/releasepaper/content/201102-64.
[13]????? Lin Y, Lai Z, Lin L, et al. Endogenous target mimics, microRNA167, and its targets ARF6 and ARF8 during somatic embryo development in Dimocarpus longan Lour[J]. Molecular Breeding, 2015, 35(12): 1-15.
[14]????? 賴鐘雄. 龍眼生物技術(shù)研究[M]. 福州: 福建科學(xué)技術(shù)出版社, 2003: 18-22.
[15]????? Chen C J, Xia R, Chen H, et al. TBtools, a toolkit for biologists integrating various biological data handling tools with a user-friendly interface[J/OL]. BioRxiv, 2018: 289660 [2020- 05-25]. http://dx.doi.org/10.1101/289660.
[16]????? Lin Y L, Lai Z X. Reference gene selection for qPCR analysis during somatic embryogenesis in longan tree[J]. Plant Science, 2010, 178(4): 359-365.
[17]????? Morris E R, Chevalier D, Walker J C. DAWDLE, a forkhead- associated domain gene, regulates multiple aspects of plant development[J]. Plant Physiology, 2006, 141(3): 932-941.
[18]????? Hiraguri A, Itoh R, Kondo N, et al. Specific interactions between Dicer-like proteins and HYL1/DRB-family dsRNA- binding proteins in Arabidopsis thaliana[J]. Plant Molecular Biology, 2005, 57(2): 173-188.
[19]????? Dong Z, Han M H, Fedoroff N. The RNA-binding proteins HYL1 and SE promote accurate in vitro processing of pri-miRNA by DCL1[J]. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(29): 9970-9975.
[20]????? Zhou C, Chen R J, Gao X L, et al. Heterologous expression of a rice RNA-recognition motif gene OsCBP20 in escherichia coli confers abiotic stress tolerance[J]. Plant Omics, 2014, 7(1): 28-34.
[21]????? Kim S, Yang J Y, Xu J, et al. Two cap-binding proteins CBP20 and CBP80 are involved in processing primary MicroRNAs[J]. Plant Cell Physiology, 2008, 49(11): 1634-1644.
[22]????? Chen X, Liu J, Cheng Y, et al. HEN1 functions pleiotropically in Arabidopsis development and Acts in C function in the flower[J]. Development, 2002, 129(5): 1085-1094.
[23]????? Bollman M K. HASTY, the Arabidopsis ortholog of exportin 5/MSN5, regulates phase change and morphogenesis[J]. Development, 2003, 130(8): 1493-1504.
[24]????? Finkelstein R. Abscisic acid synthesis and response[J]. Arabidopsis Book, 2013, 11: e0166.
[25]????? Gómez-Cadenas A, Zentella R, Walker-Simmons M K, et al. Gibberellin/abscisic acid antagonism in barley aleurone cells: site of action of the protein kinase PKABA1 in relation to gibberellin signaling molecules[J]. Plant Cell, 2001, 13(3): 667-679.
[26]????? Chen M, Du X, Zhu Y, et al. Seed Fatty Acid Reducer Acts downstream of gibberellin signalling pathway to lower seed fatty acid storage in Arabidopsis[J]. Plant Cell Environment, 2012, 35(12): 2155-2169.
[27]????? 朱華國(guó), 張獻(xiàn)龍, 金雙俠, 等. 兩種常用激素組合下棉花體細(xì)胞胚胎發(fā)生過(guò)程的組織學(xué)觀察[J]. 棉花學(xué)報(bào), 2012, 24(2): 159-166.
[28]????? 魏帥杰. 擬南芥HEN1調(diào)控胚珠發(fā)育的分子機(jī)理研究[D]. 泰安: 山東農(nóng)業(yè)大學(xué), 2019.
[29]????? 陳曉慧. 龍眼體胚發(fā)生過(guò)程中DlDCLs的克隆與功能分析[D]. 福州: 福建農(nóng)林大學(xué), 2018.
責(zé)任編輯:黃東杰
猜你喜歡
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
電子制作(2018年18期)2018-11-14 01:48:24
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
中國(guó)衛(wèi)生(2016年3期)2016-11-12 13:23:26
中國(guó)衛(wèi)生(2014年12期)2014-11-12 13:12:52
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
