溫度和稃殼去除對甘肅野生草地早熟禾發芽特性和多胚苗頻率的影響
2021-07-26 05:18:32張金青賈新鳳李凡李玉珠馬暉玲
草原與草坪 2021年3期
張金青,賈新鳳,李凡,李玉珠,馬暉玲
(甘肅農業大學草業學院,草業生態系統教育部重點實驗室,甘肅省草業工程實驗室,中-美草地畜牧業可持續發展研究中心,甘肅 蘭州 730070)
草地早熟禾(Poapratensis)主要分布在寒帶和北溫帶冷涼濕潤地區,是草原與草甸植被組成中的重要成分[1],其不僅是很多天然草地中的優良牧草,同時也被廣泛應用于草坪建植[2]。此外,它還可用于穩定侵蝕土壤、改善土壤結構和肥力[3]。由于草地早熟禾品種選育工作滯后,我國草坪建植中所用的草地早熟禾種子全是國外引進品種,因此培育我國的優良草地早熟禾新品種極其重要,這有賴于對我國草地早熟禾野生種或生態型的搜集。同時Wieners等[3]也指出若要改善草地早熟禾,如減少其在生產上的管理投入、提高其抗病蟲性、耐低修剪或其他重要農藝性狀,則需要收集和鑒定不同的種質。甘肅地處青藏高原、黃土高原、蒙新高原和西秦嶺山地的交匯地帶,境內地形、氣候復雜多樣,早熟禾屬植物資源十分豐富,這為搜集不同的草地早熟禾野生種質材料提供了便利[5]。近幾年,甘肅野生草地早熟禾的研究主要集中在抗旱性[6]、抗寒性[7]、耐踐踏性[8]、根莖擴展性[9]以及生產性能[10]等方面,而有關其多胚苗的研究較少。
草地早熟禾具有多樣的生殖系統,主要是無孢子生殖和有性雜交或自交的兼性無融合生殖體系[11]。無融合生殖具有雙胚珠、雙胚囊、雙胚雙苗或多苗等特點,這種由一粒多胚種子同時萌發出兩株或兩株以上的幼苗稱為多胚苗[12]。母錫金等[13]和田晨霞等[14]的研究均表明草地早熟禾產生雙胚苗和三胚苗是常見的。已有研究表明,植物中多胚與其無融合生殖之間具有正相關性,因而許多學者試圖通過尋找具有多胚現象的植物材料,進而篩選出一些可能具有無融合生殖特性的種質資源。植物多胚現象的發生涉及多胚的形成和表現兩大階段,前者主要由遺傳背景控制,而后者則受到外界條件的影響[14],例如黃宇[16]研究發現閩楠(Phoebebournei)種子在不同貯藏時間和培養條件下發芽率和多胚率都有所不同;黃群策等[17]也發現水稻(Oryzasativa)多胚苗發生頻率因材料種類或發芽溫度不同而表現出差異,此外也報道了種子去稃殼后可使其雙苗率提高32.4%~36.0%。同時,黃雪梅等[18]也發現黃花風鈴木(Tabebuiachrysantha)種子貯藏溫度從15℃降為10℃時,其多胚率也由63.3%降為50.0%;王巨媛[19]通過對38個韭菜(Alliumtuberosum)的多胚苗研究,指出其多胚苗發生頻率與溫度之間存在“低溫效應”,即溫度越低多胚苗發生頻率越高。可見,被子植物種子的萌發環境和萌發狀態對其多胚苗頻率具有顯著影響,但這些外界條件對草地早熟禾的多胚苗頻率是否有影響,其影響規律如何依然不清楚。因此,本研究以采集于甘肅不同地區的7個野生草地早熟禾種質為材料,探索不同萌發溫度和種子萌發時的不同狀態(除稃殼)下草地早熟禾多胚苗頻率的變化規律,以期為揭示草地早熟禾無融合生殖機理,選育我國本土的優良草地早熟禾新品種提供基礎數據。
1 材料和方法
1.1 試驗材料
供試材料為2012年8-9月采集于甘肅隴南市、甘南州夏河縣、張掖市肅南縣、定西市、天水市甘谷縣、清水縣、秦州區7個地區的野生草地早熟禾,種子經人工清選、自然干燥后,長期于25℃保存。用采集地的地名分別表示該地采集的野生草地早熟禾種質材料。
1.2 試驗方法
試驗前,對種子進行消毒處理,具體步驟為先用自來水浸泡12 h,然后用70%乙醇消毒2 min,再用20%次氯酸鈉浸泡15 min,最后用無菌水沖洗5~6次,直到去除次氯酸鈉的刺鼻味,晾干待用。
供試材料每種取8 000粒晾干后的飽滿種子,其中4 000粒置于體式顯微鏡下剝除稃殼,然后帶稃殼種子和除稃殼種子各取1 000粒為1組,每種4組,根據《牧草種子檢驗規程—發芽試驗》[20]進行發芽試驗,4組種子對應的萌發溫度分別為15℃、20℃、25℃和30℃。其他條件均相同,即在每個鋪有2層濾紙的9 cm塑料培養皿中放置100粒種子,分別加入4 mL無菌水,每處理10個重復。培養條件為光照14 h、黑暗10 h,每天采用稱重補水法補充蒸發的無菌水,25 d后結束試驗,統計并記錄各培養皿的發芽種子和雙胚苗和三胚苗及多胚苗的數量,然后計算發芽勢、發芽率和雙胚苗、三胚苗及多胚苗的頻率。計算方法參考田晨霞等[14]的方法:
發芽勢(%)=第13 d 發芽種子總數/供試種子數×100%
發芽率(%)=第25 d 發芽種子總數/供試種子數×100%
雙胚苗率(%)=雙胚苗數量/第25 d 發芽種子數×100%
三胚苗率(%)=三胚苗數量/第25 d 發芽種子數×100%
多苗率(%)=(雙胚苗數量+三胚苗數量)/第25 d 發芽種子數×100%
1.3 數據統計與分析方法
運用Microsoft Excel 2010軟件進行數據整理,數據用平均值±標準誤表示;運用SPSS 19.0統計軟件對數據進行Duncan法比較各處理間的差異顯著性。
2 結果與分析
2.1 溫度和稃殼對甘肅野生草地早熟禾發芽特性的影響
2.1.1 溫度和稃殼對甘肅野生草地早熟禾發芽勢的影響 在萌發溫度和種子狀態不同時,7個地區采集的種子發芽勢不同。相較于帶稃殼,除稃殼后隴南種子的發芽勢在相同溫度下均顯著降低,且不管稃殼是否存在,均在15℃時顯著低于其余溫度,其他溫度間沒有差異。甘南種子的發芽勢在除稃殼后均高于帶稃殼,僅15℃差異不顯著;溫度對其發芽勢的影響因種子的狀態而異,帶稃殼時,隨著溫度的上升而逐漸升高,除稃殼后,在25℃時達到最大值,30℃時顯著降低。清水種子的發芽勢變化較單一,15℃時顯著低于其余溫度,是否除稃殼的區別在于:除稃殼后,其發芽勢在相同溫度下略有升高。除稃殼后,相同溫度下肅南種子的發芽勢均顯著高于帶稃殼,且同一種子狀態時其隨著溫度的變化趨勢相同,均表現為隨著溫度升高而逐漸上升,25℃達到最大值,且與30℃無顯著差異。無論是否帶稃殼,定西種子的發芽勢均在15℃時顯著低于其他溫度,其余溫度間無顯著差異,稃殼的影響較小,除15℃時除稃殼顯著高于帶稃殼外,其余溫度下無顯著差異。秦州種子發芽勢隨溫度與稃殼的變化與清水相同,即除稃殼后略有升高;且不管有無稃殼,均在15℃時顯著低于其他溫度,且其余溫度間無顯著差異。稃殼的有無對甘谷種子發芽勢的影響為:除15℃差異不顯著外,其余溫度下除稃殼均顯著高于帶稃殼;帶稃殼時,30℃顯著高于15℃,其余溫度間無顯著差異,除稃殼后,30℃顯著高于20℃和15℃,與25℃差異不顯著(表1)。
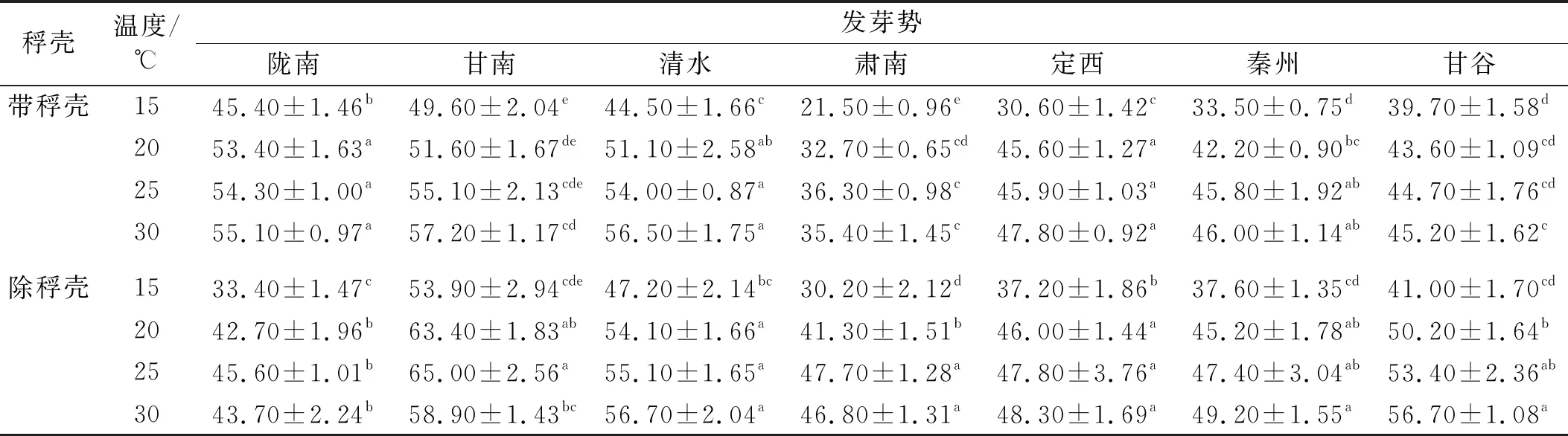
表1 溫度和稃殼對甘肅野生草地早熟禾發芽勢的影響
2.1.2 溫度和稃殼對甘肅野生草地早熟禾發芽率的影響 不同溫度與是否除稃殼對不同的野生草地早熟禾種質材料的發芽率影響不同,其中秦州種子的表現較特別,所有的處理間均無顯著差異(表2),當溫度從15℃升高到30℃時,帶稃殼種子發芽率變化幅度最大,降低了2.54%;除稃殼后,發芽率在25℃變化最大,升高了0.57%。此外,肅南的變化也不同于其余材料,不管是否除去稃殼,其發芽率均在25℃時最高,不同的是帶稃殼時25℃與其他溫度間均差異顯著,除稃殼時25℃與20℃間無顯著差異,但顯著高于15℃和30℃;稃殼對肅南發芽率的影響與溫度有關,僅20℃時除稃殼顯著高于帶稃殼,其他溫度下無顯著差異。其他材料在種子狀態相同時,其發芽率均隨著萌發溫度的升高而逐漸降低,不同的材料間其降低程度存在差異,當溫度從15℃升高到30℃時,隴南在除稃殼后降低程度最大,為25.44%。比較稃殼對其余野生草地早熟禾發芽率的影響,發現清水和甘谷在溫度不變時,其發芽率均在除去稃殼后升高,不同材料升高程度不同;但是隴南的表現正好相反:在溫度保持不變時,其發芽率在除稃殼后顯著下降,最高可下降30.17%,推測其原因可能是隴南種子的稃殼對種子的包裹比較緊密,在去除稃殼時對種子造成了一定的傷害,進而降低了其發芽率。
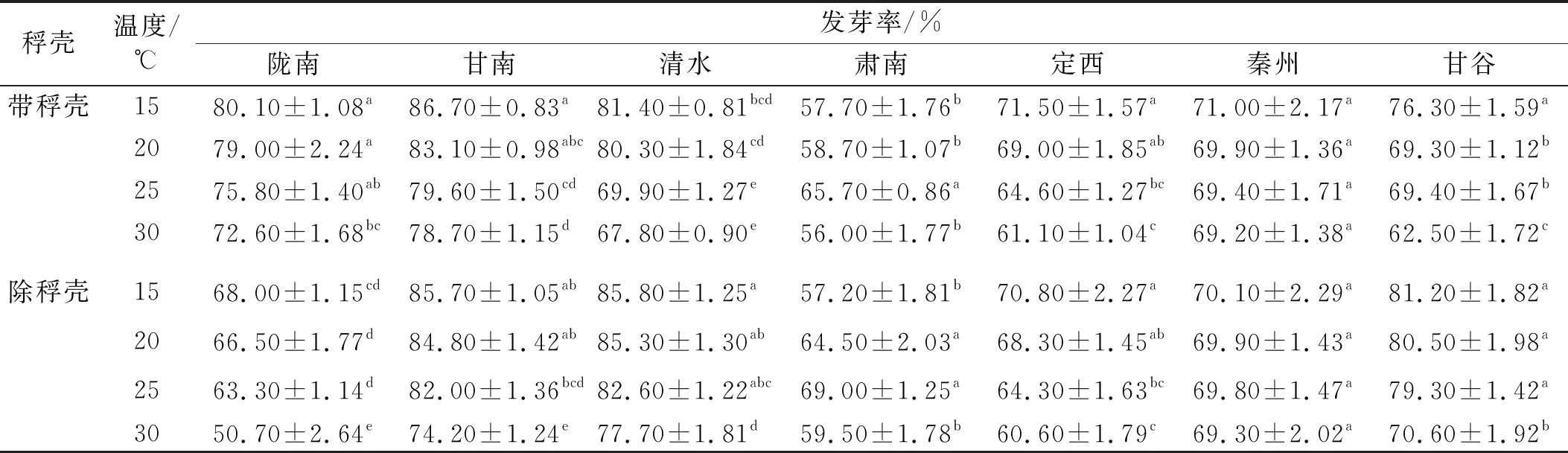
表2 溫度和稃殼對甘肅野生草地早熟禾發芽率的影響
2.2 溫度和稃殼對甘肅野生草地早熟禾多胚苗頻率的影響
2.2.1 溫度和稃殼對甘肅野生草地早熟禾二胚率的影響 不同萌發溫度下以及種子除去稃殼后隴南和甘谷采集的種子所有處理間均無顯著差異,這表明它們的二胚率不受外界條件溫度以及種子狀態的影響(表2)。甘南種子的二胚率變化最明顯,帶稃殼時,4個溫度下其二胚率均為0,除稃殼后,其二胚率隨著溫度的升高而增大。清水種子的二胚率隨著溫度的升高先上升再降低,帶稃殼種子在20℃發芽率達到最大,且與其余溫度相比差異顯著,除稃殼時25℃略高于其他溫度,但與其余溫度相比均無顯著差異。肅南種子的二胚率在種子狀態不同時,隨溫度的變化不同:帶稃殼時隨溫度升高而升高,但各溫度之間差異均不顯著;除稃殼時隨溫度升高先升高再降低,在25℃達到最大,但與其余溫度相比均無顯著差異。定西種子二胚率的變化比較規律,首先隨著溫度的升高而增大,且在30℃時顯著高于其他溫度,其余溫度間差異不顯著;其次溫度不變時,相較于帶稃殼,其二胚率在除稃殼后均略有升高,但差異不顯著。秦州種子的二胚率隨著溫度的升高先升高再降低,在20℃達到最大值,稃殼的影響在于帶稃殼時,最大值與其余溫度間均具有顯著差異,除稃殼后,最大值與其余溫度間均無顯著差異,且在溫度相同時,除稃殼后其二胚率均顯著高于帶稃殼。
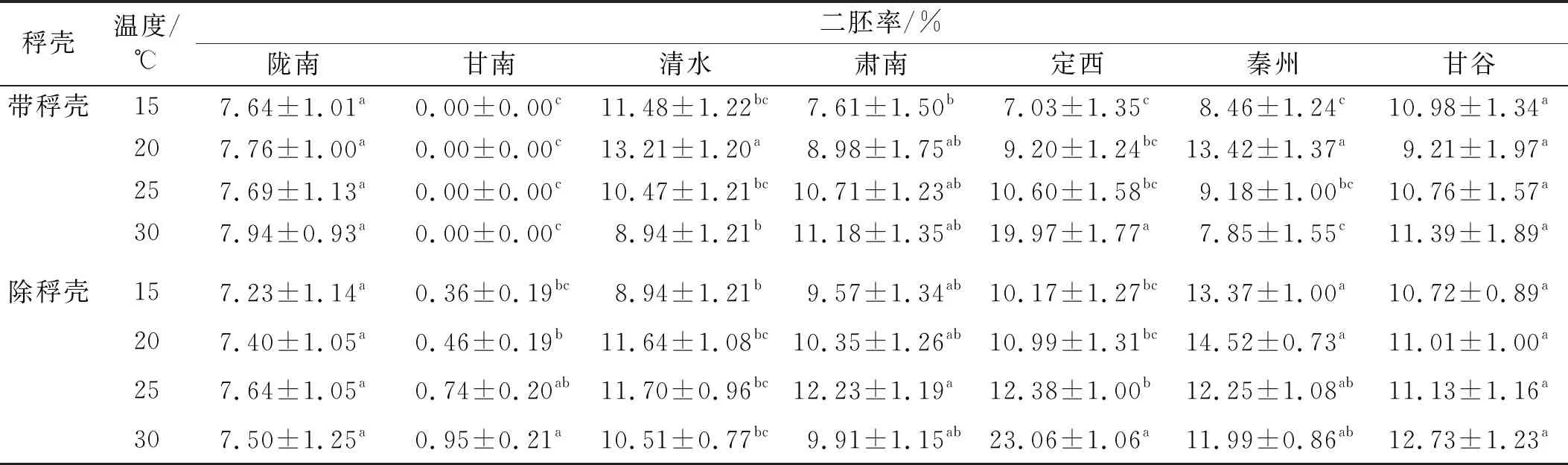
表3 溫度和稃殼對甘肅野生草地早熟禾二胚率的影響
2.2.2 溫度和稃殼對甘肅野生草地早熟禾三胚率的影響 甘肅野生草地早熟禾在不同溫度以及除稃殼處理后,各材料的三胚率變化不明顯(表4),這可能是由于草地早熟禾的三胚苗頻率偏低,導致在不同條件時變化不明顯。在所有材料中,定西種子的三胚率在任何溫度以及除稃殼后均為0,推測定西可能并不具有三胚苗的特性。甘南和清水地區采集種子的三胚率變化比較一致,相較于溫度的變化,其對于是否除稃殼的變化更敏感,在帶稃殼時,三胚率均為0,除稃殼后,不同溫度下均有三胚苗的萌發,但各溫度下差異不顯著;二者所不同的是,甘南種子的三胚率除稃殼后與帶稃殼時相比無顯著差異,而清水種子除稃殼后顯著高于帶稃殼。隴南、肅南和秦州地區采集種子的三胚率在所有處理下均無差異,表明這三個材料的三胚率不受溫度和稃殼的影響。在帶稃殼20℃和30℃時天水種子的三胚率顯著高于其他處理,其余處理下不受溫度和稃殼的影響。
2.2.3 溫度和稃殼對甘肅野生草地早熟禾多胚率的影響 隴南和甘谷種子的多胚率變化一致,在所有處理下均無差異,表明它們的多胚率不受萌發溫度和種子稃殼的影響(表5)。甘南種子的多胚率在帶稃殼時,不受溫度的影響,各溫度下均為0;除稃殼后,其多胚率隨著溫度的升高而逐漸增大。清水多胚率隨著溫度的升高先上升再降低,稃殼存在與否的差異在于前者在20℃達到最大值,與其余溫度間有差異,但不顯著,除稃殼后20℃和25℃間無顯著差異,且均高于15℃和30℃。肅南的多胚率在除稃殼25℃時達到最大值,顯著高于帶稃殼15℃,與其余溫度有差異但不顯著。定西由于其三胚率均為0,故其多胚率的變化與二胚率相同,均表現為隨著溫度的升高而增大,且在30℃時顯著高于其他溫度,其余溫度間差異不顯著;溫度不變時,相較于帶稃殼,定西的多胚率在除稃殼后均略有升高,差異不顯著。秦州的多胚率隨著溫度的升高先升高再降低,20℃時達到最大值,且帶稃殼時顯著高于其余溫度。

表5 溫度和稃殼對甘肅野生草地早熟禾多胚率的影響
3 討論
3.1 溫度及稃殼對草地早熟禾發芽特性的影響
3.1.1 溫度對草地早熟禾發芽特性的影響 溫度影響種子發芽是通過影響其代謝反應來實現的[22],當環境溫度過高或過低時,會導致細胞中的酶失活,致使細胞不能進行正常的生理代謝而死亡[23]。發芽勢和發芽率可反映種子發芽的速度和整齊度,一般來說,發芽率高、發芽勢強,表示出苗快而齊,苗壯,反之則表示出苗不齊、弱苗多。本研究發現,不管是否除稃殼,甘肅野生草地早熟禾的發芽勢均隨著溫度的升高而上升。發芽率的變化在除稃殼后以及不同的材料之間均不相同,除肅南種子外,總體趨勢為隨著溫度的上升而下降。筆者前期預實驗發現,當溫度升高到35℃時,草地早熟禾材料均不發芽,這表明草地早熟禾作為冷季型草坪草,對低溫耐性較強,而對高溫則比較敏感。然而,溫度較低時,雖然其發芽率較高,但發芽勢過低,存在出苗不齊、弱苗多的現象。此外,不同材料發芽狀況受溫度的影響并不相同,這可能與種子原產地的氣候條件有關,因此在實際生產中應根據具體的草地早熟禾材料篩選其適宜的發芽溫度。
3.1.2 稃殼對草地早熟禾發芽特性的影響 稃殼是指禾本科植物小穗基部的苞片,具有保護稃果的作用[25]。然而,有些稃殼厚而堅硬,使種子不透水、不透氣或對胚具有機械阻礙作用,而抑制種子萌發[25]。稃殼對不同植物發芽率的影響差異較大,黃群策[17]報道,水稻種子去殼可使其發芽率提高3%左右;趙昕等[27]研究發現,除稃殼后結縷草(Zoysiajaponica)種子發芽率明顯高于對照;王禹等[28]發現湘雜芒2號(Miscanthussinensis×lutarioripari)種子除稃殼后,發芽率由66%提高到86%。也有相關研究報道除稃殼降低了種子發芽率,如譚志軍等[26]的研究;任萬軍等[29]也報道去除稃殼處理可使水稻種子的成苗率降低26.1%,且不同雜交稻組合間差異較大。本研究發現,不同來源的草地早熟禾在除稃殼后,發芽率的變化不同。這表明稃殼對種子發芽的影響因植物品種和來源而異。推測除稃殼處理促進發芽率的原因可能是去殼消除了稃殼上的不透氣性、不透水性和稃殼的機械約束作用[25],也可能是稃殼中具有發芽抑制物,去除稃殼意味著發芽抑制物的消除,因而提高了發芽率。降低種子發芽率可能是種子在除稃過程中,傷害了稃果的果皮和種皮,導致營養外滲,進而引起污染[26]。因此,稃殼對發芽的影響可能是多個因素平衡后的表現,故是否能用除稃殼的方式提高植物的發芽率,則需要根據具體的材料做相關的試驗后才能應用于實踐。
3.2 溫度及稃殼對草地早熟禾多胚苗頻率的影響
3.2.1 溫度對草地早熟禾多胚苗頻率的影響 植物多胚現象的發生涉及多胚的形成和表現兩大階段,前者受到遺傳背景的控制,其表現又受到外界條件的影響,故其發生頻率受環境因素如溫度的影響[14]。本研究發現草地早熟禾的多胚苗率因材料種類或萌發溫度不同而表現出差異,這與前人的研究結果相似[16]。此外,隴南、甘南和甘谷種子的多胚率不隨溫度改變,這表明不同的材料多胚率對溫度的敏感性及反應度都不同。譚志軍等[26]認為植物多胚率是由基因型所決定的,而基因表達受外界環境的影響,故適宜的萌發溫度是植物多胚充分表達的因素之一。這可能是不同溫度時,草地早熟禾多胚苗頻率不同的原因。前人研究報道適當的高溫處理有利于水稻多胚苗頻率的提高,因為適宜的高溫可提高種子胚乳貯藏物質的轉化效率,利于多胚的萌發[26]。然而,本試驗結果不同于該報道,僅定西種子在30℃時多胚率最高,而清水和秦州種子在20℃時,其多胚率高于其他溫度,這可能是由于研究材料不同而造成的差異,也可能與實驗中所設置的具體溫度范圍有關。
3.2.2 稃殼對草地早熟禾多胚苗頻率的影響 種子稃殼的有無影響植物的多胚苗頻率,如黃群策[17]等報道,在ASDOR05-01和ASDOR05-02水稻群體內,種子去稃殼后使其雙苗頻率分別提高了32.4%和36.0%。本研究發現,不同的材料除稃殼后,多胚率變化不同,除極個別外,大多數材料在同一溫度下除稃殼后,多胚苗頻率會有所升高。這可能是由于合子胚是有性生殖,具有較強的生長優勢,且都較靠近稃殼上唯一的萌發孔,故容易萌發。然而,另一個胚是無性生殖胚,與有性胚相比其生長狀態處于劣勢,且一般都遠離萌發孔,故稃殼對其萌發具有明顯的阻礙作用,因此,稃殼的存在直接影響了多胚的發生[30]。
3.3 提高草地早熟禾多胚苗頻率的措施
草地早熟禾不僅是草坪草,還可以作為牧草利用,多胚苗的萌發及成苗會增加其地上生物量,增大裸地的覆蓋率,提高種子的產量。此外,康慶華[31]認為多胚苗可作為篩選無融合生殖種質的標志性狀,可以將具有雙胚性的材料作為誘導無融合生殖后代的遺傳基礎,同時多胚現象在胚胎發育研究、雜種優勢固定、單倍體育種及自交系選育等方面均有重要價值[32]。因此,提高多胚苗頻率在實際生產中具有重要意義。研究報道,對于具有潛在能力的多胚種子,萌發條件對多胚苗的表達起決定性作用,故優化萌發條件對于多胚苗的獲得極為重要。因此,可通過控制種子萌發條件提高其多胚率,如種子去殼、預處理溫度、Zn2+或H2O2處理、Co60-Y射線輻射、萌發溫度以及外源激素(IAA、KT、GA3、2,4-D、6-BA)處理[33]。另外,馮輝等[34]也研究報道,用0.6%二甲基亞砜(DMSO)或失活花粉處理韭菜,其多胚苗發生頻率明顯提高。
本研究結果表明,草地早熟禾主要有單胚苗、二胚苗和三胚苗3種種苗,其頻率大小為單胚苗>二胚苗>三胚苗,且不同野生材料的多胚頻率存在差異,萌發溫度和去除稃殼對其影響也不同,這表明草地早熟禾的多胚現象存在材料特異性,同時該結果也表明了草地早熟禾的種子萌發環境對其多胚苗頻率具有一定的影響。因此,前人所報道的Zn2+、H2O2、DMSO、Co60-Y以及外源激素是否能提高草地早熟禾的多胚率,適宜的濃度和處理方式等問題依然需要進一步的試驗后才能確定。
4 結論
通過對去除稃殼的7個甘肅野生草地早熟禾材料的發芽特性和多胚苗率分別于15℃、20℃、25℃和30℃條件下的統計分析表明,草地早熟禾的發芽特性和多胚苗頻率隨著材料來源、萌發溫度和種子狀態的不同而異。其中隴南和甘谷種子在所有處理下其多胚率均無顯著差異,以省時、省力、經濟的原則篩選出其余材料多胚率最高的萌發條件分別為:甘南和定西種子為30℃,且前者需要去除稃殼;清水和肅南種子為25℃,且后者需要去除稃殼;秦州種子為20℃。具體的草地早熟禾材料多胚率最高的萌發條件不盡相同,需提前進行試驗才能確定其最適萌發條件。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
