北極新奧爾松地區(qū)藻類可培養(yǎng)附生細(xì)菌多樣性研究
2021-07-23 01:10:40孫曉夢樊守金
極地研究 2021年2期
關(guān)鍵詞:變形
孫曉夢 樊守金
(1山東師范大學(xué)生命科學(xué)學(xué)院, 山東 濟(jì)南 250014;2山東大學(xué)微生物技術(shù)國家重點(diǎn)實(shí)驗(yàn)室, 山東 青島 266237)
提要 為研究北極地區(qū)海洋藻類可培養(yǎng)附生細(xì)菌的多樣性, 利用稀釋平板法自2019年7月采集的北極新奧爾松地區(qū)的海洋紅藻多管藻(Polysiphonia sp.)樣品中分離純化得到60株可培養(yǎng)的藻類附生細(xì)菌。16S rRNA基因序列測定分析表明所分離的藻類附生細(xì)菌主要隸屬于γ-變形菌綱(39株, 65%), 其次隸屬于放線菌綱(10株, 17%)和黃桿菌綱(8株, 13%), 僅有少量隸屬于α-變形菌綱(2株, 3%)和芽孢桿菌綱(1株, 2%), 其中的優(yōu)勢細(xì)菌屬為Psychrobacter屬(22株, 37%)和Pseudoalteromonas屬(12株, 20%)。此次獲得了較多極地藻類多管藻的附生細(xì)菌菌株, 初步揭示了多管藻可培養(yǎng)附生細(xì)菌的多樣性, 為多管藻自身以及其附生細(xì)菌生態(tài)功能及應(yīng)用開發(fā)研究奠定了基礎(chǔ)。
0 引言
藻類是海洋環(huán)境中極為重要的資源, 它們種類繁多, 作為生產(chǎn)者在海洋生態(tài)系統(tǒng)中起著重要作用。藻類在生長過程中向環(huán)境釋放糖類、氨基酸、脂類、酶和毒素等成分, 使藻體周圍形成獨(dú)特“藻際微環(huán)境”, 簡稱“藻際”(phycosphere)[1-2]。“藻際”間存在相同的或特異性的藻類附生和浮游細(xì)菌群落[3-4]。浮游細(xì)菌被認(rèn)為是藻類附生細(xì)菌的主要來源, 對藻類附生細(xì)菌群落組成有重要影響[5-6], 附生細(xì)菌也可能擴(kuò)散到浮游細(xì)菌中[7-8];而藻類自身特性、不同生長階段的分泌物及藻類生長的環(huán)境條件(如pH值、水流、光照、溫度和養(yǎng)分)也影響附生細(xì)菌群落的組成[9-10]。這些藻類附生菌與藻類間存在著復(fù)雜的相互作用關(guān)系, 主要表現(xiàn)為互利共生、拮抗和競爭等作用, 研究藻類附生細(xì)菌多樣性可以促進(jìn)對藻菌藻際相互作用關(guān)系及其機(jī)制的了解。Lachnit等[11]利用DGGE與16S rRNA文庫構(gòu)建相結(jié)合的技術(shù)分別于2007—2008年連續(xù)兩年的春季和秋季, 對基爾峽灣的褐藻(Fucus vesiculosus)、紅藻(Gracilaria vermiculophylla)、綠藻(Ulva intestinalis)的附生菌群落結(jié)構(gòu)進(jìn)行了分析。結(jié)果發(fā)現(xiàn)海洋大型藻類藻體表面具有物種特異性和時(shí)間適應(yīng)性的附生細(xì)菌生物膜, 其中寄主特異性細(xì)菌組合要么避免了真核生物幼蟲的定殖, 要么表現(xiàn)出強(qiáng)大的抗菌活性,在藻類生存中發(fā)揮重要作用。近年來, 海洋大藻類已成為微生物多樣性和生物活性次生代謝物的豐富來源[12], 多管藻(Polysiphoniasp.)隸屬于紅藻門(Rhodophyta)、紅藻綱(Rhodophyceae)、仙菜目(Ceramiales)、松節(jié)藻科(Rhodomelaceae)、多管藻屬(Polysiphonia), 有豐富的生物活性物質(zhì), 而且具有一定的抗菌、抗氧化、抗腫瘤等多種生理活性。以往的研究多針對于溫帶地區(qū)多管藻附生細(xì)菌的應(yīng)用研究, 對于極地多管藻的附生細(xì)菌的應(yīng)用研究較少: Romanenko等[13]從日本海濱紅藻樣品(PolysiphoniaGreville, 1823, family Rhodomelaceae)中分離出隸屬于3個(gè)細(xì)菌門(變形菌門、放線菌門和厚壁菌門)12個(gè)細(xì)菌屬的80株附生細(xì)菌, 其中變形菌門占優(yōu)勢。北極地區(qū)由于其獨(dú)特的地理環(huán)境與氣候特征, 蘊(yùn)含豐富獨(dú)特的極地藻類和微生物資源, 藻類附生細(xì)菌群落組成具有特殊性。本文采集了北極新奧爾松地區(qū)的多管藻樣品, 進(jìn)一步研究了這類極地藻類樣品中可培養(yǎng)附生細(xì)菌的多樣性, 為北極藻類附生微生物的生態(tài)作用及應(yīng)用開發(fā)研究奠定了基礎(chǔ)。
1 實(shí)驗(yàn)方法
1.1 材料
1.1.1 樣品的采集
于2019年7月利用無菌剪刀和容器采集北極王灣新奧爾松附近的潮間帶多管藻(Polysiphoniasp.)樣品(11°42′3.24″, 78°57′36.72″)。所采集藻類樣品經(jīng)適當(dāng)分裝后置于無菌容器中于4℃保存,空運(yùn)(24小時(shí))至實(shí)驗(yàn)室進(jìn)一步處理。
1.1.2 2216E培養(yǎng)基(g·L-1)
蛋白胨5.0, 酵母粉1.0, 檸檬酸鐵0.1, 氯化鈉19.45, 氯化鎂5.98, 硫酸鈉3.24, 氯化鈣1.8,氯化鉀0.55, 碳酸鈉0.16, 溴化鉀0.08, 氯化鍶0.034, 硼酸0.022, 硅酸鈉0.004, 氟化鈉0.0024,硝酸鈉0.0016, 磷酸氫二鈉0.008, pH值7.4~7.8。2216E固體培養(yǎng)基配制時(shí)添加15.0瓊脂。
1.1.3 瓊膠唯一碳源培養(yǎng)基(g·L-1)
氯化銨0.5, 氯化鈉30.0, 六水氯化鎂3.0, 硫酸鉀2.0, 磷酸氫二鉀0.2, 氯化鈣0.01, 六水氯化鐵0.006, 七水鉬酸鈉0.005, 二水氯化銅0.004,Tris 6.0, 瓊膠10.0, pH值7.5~8.0。
1.2 方法
1.2.1 菌株分離和保藏
取多管藻樣品少許, 用滅菌的天然海水對其進(jìn)行三次沖洗, 去除表面雜物后將樣品懸浮在20~30 mL無菌海水中, 劇烈震蕩1 min, 取震蕩后的溶液與無菌海水以1∶9的比例混合, 梯度稀釋至濃度為10-1、10-2、10-3、10-4、10-5和10-6的菌液, 每個(gè)稀釋度的菌液100μL菌液涂布在含2216E固體培養(yǎng)基的平板上, 每個(gè)梯度做三個(gè)重復(fù)。將涂布后的平板置于在15℃下培養(yǎng), 培養(yǎng)3周后挑取平板上不同形態(tài)的菌落在含新鮮培養(yǎng)基的平板上進(jìn)行多次劃線分離純化。獲得純培養(yǎng)的菌株接種于含5 mL 2216E液體培養(yǎng)基試管內(nèi),15℃振蕩培養(yǎng)3~5 d, 獲得的培養(yǎng)液添加20%甘油(v·v-1)后于-80 ℃低溫保存。
1.2.2 基因組DNA的提取
菌株接種于含5 ml 2216E液體培養(yǎng)基試管內(nèi),15℃振蕩培養(yǎng)3~5 d, 培養(yǎng)液10000 rpm離心獲得菌體, 利用BioTeKe的細(xì)菌基因組提取試劑盒(BioTeKe Corporation公司, 北京)按照操作說明提取。
1.2.3 PCR擴(kuò)增及產(chǎn)物測序
擴(kuò)增分離菌株16S rRNA基因所用引物為27F(5′-AGAGTTTGATCCTGGCTCAG-3′)和 1492R(5′-GGTTACCTTGTTACGACTTC-3′)[14]。
PCR 反應(yīng)體系: Nuclease-free Water 9.4 μL,2×TransTaq? HiFi PCR SuperMix 10μL, 27F 0.2 μL,1492R 0.2 μL, 細(xì)菌基因組DNA(模板)0.5 μL。
PCR反應(yīng)條件: 94℃, 5 min; 94℃, 30 s, 55℃,30 s, 72℃, 90 s, 30個(gè)循環(huán); 72℃, 10 min。
PCR產(chǎn)物直接送至青島擎科測序公司經(jīng)純化后利用ABI 3730XL DNA測序儀進(jìn)行兩端測序,去除不確定堿基后, 利用ChromasPro軟件進(jìn)行序列拼接, 獲得不同附生細(xì)菌的16S rRNA基因序列。
1.2.4 系統(tǒng)發(fā)育樹的構(gòu)建
所有可培養(yǎng)附生菌的16S rRNA基因序列利用Blastn程序在GenBank數(shù)據(jù)庫進(jìn)行序列相似性搜索, 獲取序列相似性較高的菌株信息, 若兩種細(xì)菌的16S rDNA序列同源性低于98%, 可以認(rèn)為屬于不同種[15]。采用軟件MEGA 7.0[16]利用Neighbor-Joining法和Kimura 2-parameter model構(gòu)建系統(tǒng)進(jìn)化樹。系統(tǒng)樹各分支的自舉檢驗(yàn)值(bootstrap values)由1000次重復(fù)檢驗(yàn)獲得。
2 結(jié)果與分析
2.1 多管藻附生細(xì)菌分類結(jié)果
從多管藻的表面分離可培養(yǎng)附生細(xì)菌60株。基于16S rRNA基因序列相似性對比, 發(fā)現(xiàn)它們隸屬于4個(gè)細(xì)菌門 [變形菌門(Proteobacteria), 放線菌門(Actinobacteria), 擬桿菌門(Bacteroidetes)和厚壁菌門(Firmicutes)]中的12個(gè)屬。這些菌株大部分(65%)屬于變形菌門, 隸屬于γ-變形菌綱(γ-Proteobacteria)中的Pseudoalteromonas(12株)、Halomonas(1株)、Psychrobacter(22株)、Cobetia(2株)、Shewanella(1株)和Marinomonas(1株)6個(gè)屬以及α-變形菌綱(α-Proteobacteria)中的Paracoccus(1株)和Yoonia(1株)2個(gè)屬; 少部分菌株(30%)屬于放線菌門和擬桿菌門, 隸屬于放線菌綱(Actinobacteria)的Salinibacterium屬(10株)和黃桿菌綱(Flavobacteriia)的Winogradskyella(4株)、Brumimicrobium(4株)2個(gè)屬。僅有1株菌株屬于厚壁菌門桿菌綱(Bacilli)的Planomicrobium屬。將所有菌株的16S rRNA基因序列在NCBI的BLAST中進(jìn)行兩兩比對, 序列相似度高于98.5%的菌株視為同種菌株, 各菌株分類隸屬關(guān)系見表1。

表1 多管藻各可培養(yǎng)附生細(xì)菌分類隸屬關(guān)系Table 1.Taxonomic affiliation of epiphytic bacterial strains isolated from the Polysiphonia sp.
有56.6%的分離菌株隸屬于γ-變形菌綱(γ-Proteobacteria)的Psychrobacter(22株)和Pseudoalteromonas(12株)兩個(gè)細(xì)菌屬, 這兩屬的成員在海洋低溫環(huán)境中分布廣泛; 有16.6%(10株)的分離菌株隸屬于放線菌綱(Actinobacteria)的Salinibacterium屬, 這些分離菌株均與該屬來自海水細(xì)菌種Salinibacterium amurskyense親緣關(guān)系接近。隸屬于其他9屬的分離菌株均少于5株,說明其相對豐度較低。
將多管藻的各稀釋梯度分離出的附生細(xì)菌種屬分布相對比, 結(jié)果如圖1所示, 從各稀釋梯度分離得到的附生細(xì)菌存在較大差異性: γ-變形菌綱(γ-Proteobacteria)中Psychrobacter屬在7個(gè)梯度中均存在, 且在大部分梯度中占比較大,Pseu-doalteromonas屬在100~10-3梯度中存在, 且占比較大; 放線菌綱(Actinobacteria)的Salinibacterium屬和黃桿菌綱(Flavobacteriia)的Winogradskyella屬在較低的稀釋梯度存在, 但占比較小; 其他8個(gè)屬均僅在某一梯度存在, 不占優(yōu)勢。進(jìn)一步證明了γ-變形菌綱(γ-Proteobacteria)在所分離細(xì)菌中的優(yōu)勢地位。

圖1 多管藻的可培養(yǎng)附生細(xì)菌菌群屬水平上比較Fig.1.Comparison of community compositions of the cultivable epiphytic bacteria of the Polysiphonia sp.on the genus level
2.2 16S rRNA基因序列分析
基于16S rRNA基因序列構(gòu)建的分離菌株的系統(tǒng)進(jìn)化樹見圖2。其中分離菌株H-3-4-2、H-3-5-1、H-3-5-5、H-3-5-8、H-3-5-10和H-3-5-11與數(shù)據(jù)庫中已知細(xì)菌種模式菌株的16S rRNA 基因序列的相似性均小于98.0%, 可能代表細(xì)菌新的種(屬)。其中H-3-4-2與Marinomonas屬的Marinomonas ushuaiensis模式菌株16S rRNA 基因序列的相似性為97.46%, H-3-5-1、H-3-5-8、H-3-5-10和H-3-5-11與Brumimicrobium屬的Brumimicrobium mesophilum模式菌株16S rRNA基因序列的相似性分別為97.84%、97.75%、97.34%和97.70%, 而菌株H-3-5-5與Yoonia屬的Yoonia sediminilitoris模式菌株16S rRNA 基因序列的相似性為95.46%, 并且在進(jìn)化樹中均形成獨(dú)立分支。這些說明北極多管藻中蘊(yùn)含新型菌株資源。
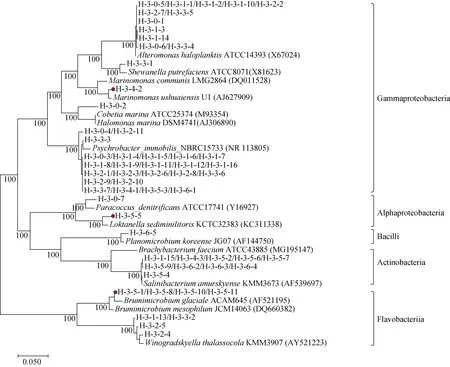
圖2 利用NJ法基于16S rRNA基因序列構(gòu)建的北極多管藻附生細(xì)菌的系統(tǒng)進(jìn)化樹Fig.2.A neighbor-joining phylogenetic tree based on the 16S rRNA gene sequences of 60 Epiphytic bacteria from the Arctic Polysiphonia algae (accession numbers are given in parentheses)
2.3 降解瓊膠的附生細(xì)菌篩選
由于多管藻的多糖組分與瓊膠較為相似[17],并且瓊膠酶在分子生物學(xué)、水產(chǎn)養(yǎng)殖以及原生質(zhì)體的制備中有較大的應(yīng)用前景[18], 因此對篩選出的附生菌株進(jìn)行瓊膠酶的初步篩選, 挑取純化的單菌落涂布在瓊膠的唯一碳源培養(yǎng)基上, 培養(yǎng)一段時(shí)間后觀察生長狀況, 初步判斷菌株的瓊膠降解能力, 結(jié)果如表2所示:Cobetia crustatorum、Marinomonas ushuaiensis、Paracoccus sediminilitoris、Pseudoalteromonas distincta、Pseudoalteromonas fuliginea、Pseudoalteromonas nigrifaciens、Pseudoalteromonas translucida、Psychrobacter fozii、Psychrobacter nivimaris、Psychrobacter okhotskensis和Winogradskyella undariae11種菌株可以在瓊膠的唯一碳源培養(yǎng)基上生長, 其中Pseudoalteromonas和Psychrobacter兩個(gè)細(xì)菌屬占比較大, 可以初步判定它們有瓊膠降解能力。

表2 多管藻各可培養(yǎng)附生細(xì)菌瓊膠降解能力Table 2.Agar-degrading ability of epiphytic bacterial strains isolated from the Polysiphonia sp.
3 討論
利用培養(yǎng)的方法研究微生物多樣性具有一定局限性[19-20], 但目前依然是調(diào)查藻類附生細(xì)菌常用手段。本文利用稀釋平板法從北極潮間帶采集的1種海洋紅藻多管藻中獲得60株附生細(xì)菌菌株,它們隸屬于放線菌綱、α-變形菌綱、芽孢桿菌綱、黃桿菌綱和γ-變形菌綱等五綱中12個(gè)細(xì)菌屬, 所獲得細(xì)菌菌株主要集中分布在 γ-變形菌綱Pseudoalteromonas屬和Psychrobacter屬及放線菌綱Salinibacterium屬中。苗禎等[21]對5株北極微藻的不同生長時(shí)期的附生細(xì)菌和游離細(xì)菌的16S rRNA基因進(jìn)行PCR以及DGGE 分析, 研究發(fā)現(xiàn)5株微藻具有不同的藻際微生物群落結(jié)構(gòu)組成, 每個(gè)藻株的主要附生微生物群落組成差異顯著, 說明藻類附生細(xì)菌群落組成與藻類的種類密切相關(guān)。Bidle等[22]通過對海洋環(huán)境中的硅藻的附生細(xì)菌分析也發(fā)現(xiàn)它們主要由γ-變形菌綱組成,僅有極少數(shù)α-變形菌綱, 同時(shí)發(fā)現(xiàn)γ-變形菌綱的菌株在藻類生長的延滯期、指數(shù)期和穩(wěn)定期都作為優(yōu)勢定殖者持續(xù)存在于硅藻表面, 這與本文多管藻附生細(xì)菌群落中γ-變形菌綱成員為優(yōu)勢類群的結(jié)論一致, 說明了γ-變形菌綱的成員可能具有較強(qiáng)的藻類表面定殖和生存能力。并且有研究表明γ-變形菌綱不僅在藻類附生菌中占優(yōu)勢地位, 在北極其他海洋環(huán)境中也廣泛存在且占優(yōu)勢地位, γ-變形菌綱可以在多種不同濃度的碳源上快速生長的[23], 進(jìn)一步說明了γ-變形菌綱具有較強(qiáng)生存能力。
王子峰等[24]利用TCBS培養(yǎng)基從多管藻(Polysiphonia urceolata)表面分離得到6株細(xì)菌。生理生化試驗(yàn)和gyrB基因序列分析結(jié)果表明6株細(xì)菌都屬于變形菌門Vibrio屬。Lyudmila等[25]從日本海濱紅藻樣品(PolysiphoniaGreville, 1823,family Rhodomelaceae)中分離到5株革蘭氏陰性需氧嗜鹽菌, 并進(jìn)行了分類研究。根據(jù)16S rRNA基因序列分析, 5株分離株隸屬于放線菌門Labrenzia屬。Romanenko等[13]從日本海濱紅藻樣品(PolysiphoniaGreville, 1823, family Rhodomelaceae)中分離到的細(xì)菌隸屬于Pseudovibrio、Labrenzia、Sphingomonas(α-變形菌綱);Cobetia、Shewanella、Vibrio(γ-變形菌綱);Microbacterium、Dietzia、Agrococcus、Streptomyces(放線菌門);Bacillus、Paenibacillus(擬桿菌門)。Goecke等[26]認(rèn)為在大型藻類的附生細(xì)菌中通常存在變形菌門和擬桿菌門的細(xì)菌。Bakunina等[27]從鄂霍次克海的紅藻樣品Polysiphoniasp.分離研究擬桿菌門菌株的糖苷酶活性, 發(fā)現(xiàn)Winogradskyella細(xì)菌屬在數(shù)量上占重要地位, 但缺少糖苷酶活性, 可能對藻類生長起其他重要作用。將本文結(jié)果與以往研究對比發(fā)現(xiàn), 多管藻的附生細(xì)菌多集中在4個(gè)細(xì)菌門 (變形菌門、放線菌門、擬桿菌門和厚壁菌門)中, 但是在細(xì)菌種屬分類甚至是細(xì)菌科的分類上差異較大。研究表明革蘭氏陰性菌在海洋中占據(jù)主要地位, 與藻類相關(guān)的革蘭氏陽性異養(yǎng)菌的占比較少[28], 例如在紅藻Delesseria sanguineawere和囊藻Fucus vesiculosus中分離的166株附生細(xì)菌中僅有1株細(xì)菌屬于Salinibacterium屬[29], 在以往的關(guān)于多管藻的研究中也少有革蘭氏陽性菌的報(bào)道, 但是本次采集的多管藻附生細(xì)菌中Salinibacterium屬占比較大(16.7%),這可能與北極地區(qū)多管藻自身的性質(zhì)有關(guān)。
藻類附生細(xì)菌常產(chǎn)生多種生物活性物質(zhì)[30-34],包括酶類及小分子的抗菌物質(zhì)等, 以助其在藻類表面定殖及生存[33]。交替單胞菌屬、芽孢桿菌屬、假交替單胞菌屬、假單胞菌屬、鏈霉菌屬和弧菌屬細(xì)菌是常見的可產(chǎn)生抗菌物質(zhì)的的藻類附生菌[31-32]。根據(jù)以往的報(bào)道, 多管藻本身具有較好的抗菌能力[35-36], 由于藻際菌藻相互作用的關(guān)系, 多管藻的附生細(xì)菌也可能有較好的抗菌能力。本文所分離附生細(xì)菌中Psychrobacter、Pseudoalteromonas和Salinibacterium屬菌株占有優(yōu)勢, 這3個(gè)屬菌株均具有較強(qiáng)的低溫酶及小分子抗菌物質(zhì)產(chǎn)生能力[29,37-39], 對可能有瓊膠降解能力的菌株進(jìn)行了初步篩選, 通過表2我們可以發(fā)現(xiàn)降解菌株主要集中在Pseudoalteromonas和Psychrobacter兩個(gè)細(xì)菌屬中, 與前人研究一致, 但是對于本文所獲得藻類附生細(xì)菌其他低溫酶和抑菌活性物質(zhì)的進(jìn)一步篩選研究工作正在進(jìn)行中。
本研究對北極新奧爾松地區(qū)海洋藻類多管藻可培養(yǎng)附生細(xì)菌多樣性做了初步研究, 獲得了較多的藻類附生細(xì)菌菌株, 這為研究極地藻類附生細(xì)菌的生態(tài)作用及其開發(fā)應(yīng)用奠定了基礎(chǔ)。
猜你喜歡
智慧少年·故事叮當(dāng)(2020年10期)2020-11-06 06:19:00
中華詩詞(2020年1期)2020-09-21 09:24:52
河北理科教學(xué)研究(2020年1期)2020-07-24 08:14:34
作文成功之路·小學(xué)版(2020年11期)2020-02-01 06:26:58
作文周刊·小學(xué)二年級(jí)版(2018年29期)2018-11-26 11:20:28
小學(xué)生作文(中高年級(jí)適用)(2018年5期)2018-06-11 01:22:56
童話世界(2018年14期)2018-05-29 00:48:08
數(shù)學(xué)小靈通·3-4年級(jí)(2017年10期)2017-11-08 08:42:59
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2017年11期)2017-04-23 07:18:00
數(shù)學(xué)大王·中高年級(jí)(2016年12期)2016-12-26 21:37:36
