歐拉型藏羊IL21基因序列、同源性及蛋白特性分析
2021-07-21 05:58:14陳曉慧冶貴生
家畜生態學報 2021年6期
關鍵詞:分析
陳曉慧,李 清,冶貴生*
(1.青海大學 農牧學院,青海 西寧,810016;2.青海大學 省部共建三江源生態與高原農牧業國家重點實驗室,青海 西寧,810016)
歐拉型藏羊主要分布在青藏高原,由于受到地理環境等因素的影響,歐拉型藏羊擁有較高的抗寒性與抗病性[1]。為了提高藏羊對疾病的抵抗力,發掘藏羊抗病免疫基因對其種質資源的開發具有重要意義。早在20世紀末,Parrish-Novak等[2]提出白細胞介素-21(IL21)。IL21主要是由濾泡輔助T 細胞(Tfh)、輔助性T細胞17(Th17)和自然殺傷細胞(NK)產生[3-4],可增強CD+8 T細胞的擴增和CD+4 T細胞向效應T細胞的分化[5-6]。IL21受體(IL21Rα)可有助于B淋巴細胞、T淋巴細胞和NK細胞等細胞的增殖與分化,IL21與其受體IL21Rα連接可促進B細胞依賴性Ig G產生,從而增強細胞毒性和提高殺傷能力[7-8]。IL21作為一種多效性細胞因子,在先天性免疫和適應性免疫應答中發揮著重要作用,與多種自身免疫性疾病有關。IL21能抑制腫瘤的生長繁殖,通過促使合成IFN-γ在抗腫瘤中有著良好效果[9]。IL21最開始對Th1細胞進行誘導分化,使得機體細胞對病毒有一定的免疫效果,合成大量IFN-γ,這對NK細胞的增殖和CTL效應有著一定的推動作用;IL21還可以通過聯合其他細胞因子活化STAT1和STAT3等信號通路[10],不但對細胞的毒活性有所提升,更是可以良好的促進IFN-γ分泌,使得效應T細胞和NK細胞可以更好地工作,實現抑制腫瘤生長的效果[11-12]。另外,在HIV感染的患者中,IL21提高了CD+8 T細胞和NK細胞分泌穿孔素以此來抑制HAVD的復制[13],控制HIV感染。由于IL21對HIV患者的T細胞、NK細胞和B細胞可以起到一個較好的改善效果,所以在對HIV患者的診斷與該病毒的研究途中,IL21可作為重要的生物標志物[14]。劉秀煥等[8]發現加強IL21的功能有利于癌癥、HIV感染等疾病的治療,但在一些免疫系統疾病如SLE和RA中,降低IL21的作用可提高臨床療效。目前關于藏羊IL21的研究未見報道,本研究使用PCR方法擴增了藏羊IL21基因,并進行了生物信息學分析,為歐拉型藏羊IL21免疫調控機理的研究提供基礎數據。
1 材料與方法
1.1 引物設計與合成
根據NCBI登錄的綿羊IL21基因序列,使用oligo引物設計軟件設計歐拉型藏羊IL21基因擴增引物,由TaKaRa公司(大連)合成。引物序列如下:IL21-F:5'-CACAAGTCAAGCTTCCAA-3',IL21-R: 5'-GGACAGATGCTGATGAATC-3',目的基因長度387 bp。
1.2 試驗材料
試驗樣品歐拉型藏羊的脾臟組織采自青海省河南縣屠宰場,由青藏高原動物疾病研究室采集、液氮速凍,-80 ℃冷凍保存。
1.3 試驗試劑
RNA 提取試劑盒由 Aidlab Biotechnologies Co. Ltd提供;Prime Script 反轉錄試劑盒、預混液由 TaKaRa 公司(大連)提供。
1.4 提取總 RNA
取歐拉型藏羊的脾臟組織通過液氮研磨成末狀,根據試劑盒說明書提取總 RNA。
1.5 反轉錄 cDNA
提取的歐拉型藏羊脾臟總 RNA通過反轉錄為 cDNA (操作依說明書)。
1.6 PCR 擴增
IL21基因的 PCR 合成體系:預混液 Premix Taq 25 μL、ddH2O 18 μL、模板 cDNA 5 μL、IL21 基因上、下游引物各1 μL。
反應條件:94 ℃ 1 min、50 ℃ 50 s、72 ℃ 1 min,35個循環;72 ℃ 10 min 延伸,4 ℃ 保存。
1.7 IL21基因序列分析
將擴增產物送至測序公司進行測序,然后進行IL21基因核苷酸序列分析和同源性比對分析。
1.8 IL21 蛋白結構分析
使用 DNAStar、PredictProtein、I-Tasser 在線服務器對 IL21 蛋白結構進行預測分析。
2 結果與分析
2.1 IL21基因 PCR 擴增
經過凝膠電泳檢測,應用IL21基因特異引物擴增出的產物大小約為 387 bp,與預期片段大小相符(如圖1),條帶特異性強。

圖1 IL21基因 PCR 擴增瓊脂糖凝膠電泳M. DNA Marker;1.陰性對照;2. PCR產物Fig. 1 Agarose gel electrophoresis of IL21 gene PCRM. DNA Marker;1. Negative control;2. PCR product
2.2 IL21基因核苷酸序列分析
序列分析結果表明(表1),IL21基因片段長度為 387 bp,在堿基組成中腺嘌呤含量占 35.92%,且AT含量高于GC含量。
2.3 IL21基因同源性比對分析
本研究將歐拉型藏羊IL21基因與不同種類羊和不同種類牛的IL21基因核苷酸序列、氨基酸序列同源比對。用來做對比分析羊的種類包括山羊、綿羊和藏羚羊等;用來作對比分析牛的種類包括水牛、牦牛、瘤牛和美洲草原野牛等。結果表明歐拉型藏羊IL21基因與參考綿羊(登錄號:NM_001256817.1)核苷酸序列同源性最高,為100%;與參考藏羚羊(登錄號: XM_005971185.1)核苷酸序列同源性為99.5%;與參考山羊(登錄號: XM_005691285.2)核苷酸序列同源性為99.0%,僅次于藏羚羊;與參考美洲草原野牛(登錄號:XM_010860276.1)核苷酸序列同源性為98.4%;與參考牦牛(登錄號: XM_005899735.1)核苷酸序列同源性為98.4%;與參考水牛(登錄: XM_006078950.2)核苷酸序列同源性為98.2%;與參考瘤牛(登錄號: XM_019977155.1)核苷酸序列同源性為98.2%。另外,氨基酸序列同源性依次為100%、100%、100%、98.4%、98.4%、98.4% 和97.7%,如圖 2 和圖 3 所示。

表1 IL21基因核苷酸序列分析Table 1 Nucleotide sequence analysis of IL21 gene
2.4 IL21蛋白結構分析
經分析發現,IL21 蛋白中 α 螺旋占 53.5%,其次是 β 轉角占 27.1%,β 折疊最少,為 2.33%(圖4);親水性分析結果顯示,IL21 蛋白質的主要親水區是1~13、26~35、37~50、57~71、77~99、101~115、121~129(如圖5);表面可及性分析表明,該蛋白在 1~8、24~28、39~43、61~70、77~94、102~116 位氨基酸呈現表面可及性較大(圖6);IL21 蛋白三級結構模型見圖7。
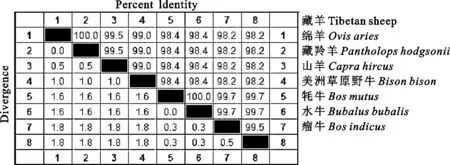
圖2 IL21基因核苷酸序列同源比對Fig. 2 The nucleotide sequence homology of IL21 gene

圖3 IL21蛋白氨基酸序列同源比對Fig.3 The amino acid homology of IL21 protein
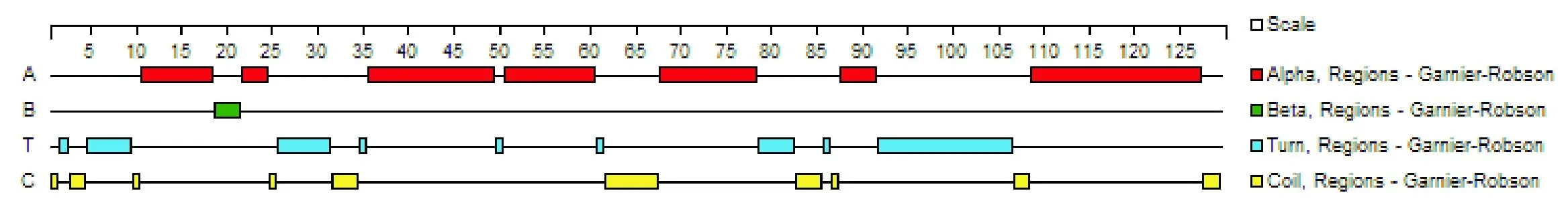
圖4 IL21 蛋白二級結構Fig. 4 Secondary structure of IL21 protein

圖5 IL21 蛋白親水性Fig. 5 Hydrophilicty of IL21 protein
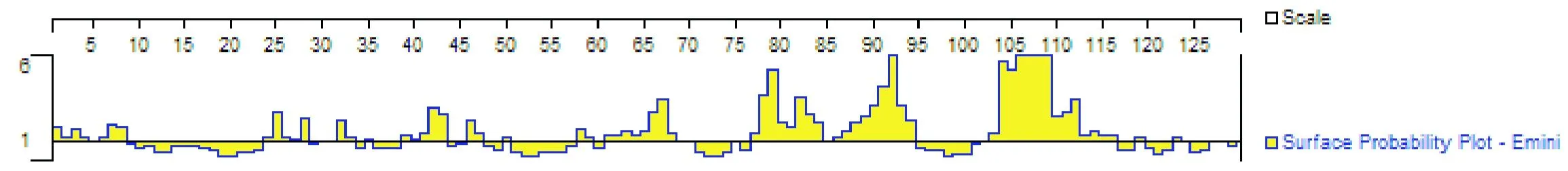
圖6 IL21 蛋白表面可及性Fig. 6 Surface accessibility of IL21 protein
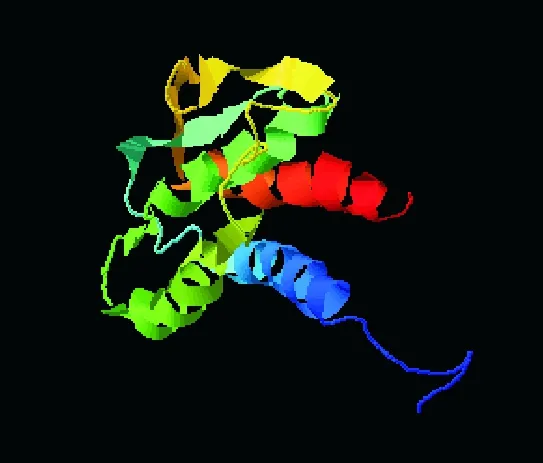
圖7 IL21 蛋白三級結構模型Fig. 7 Tertiary structure model of IL21 protein
2.5 蛋白功能位點預測
通過對歐拉型藏羊IL21蛋白功能位點分析發現,該蛋白含有 1 個酰胺化位點,為88~91 位的AGRR;1個亮氨酸拉鏈模式,為11~33位LFIRLRQLIDIVDQLKNYVNDL。
3 討 論
藏羊作為青藏高原的優良品種,具有抗病性強的種質特征,因此發掘歐拉型藏羊抗病免疫基因,對其種質資源的開發具有重要研究意義。IL21作為免疫機制輔助因子,在調控機體免疫應答機制中發揮重要作用,研究發現IL21可視為B細胞的免疫抑制因子,用于阻止免疫球蛋白合成[15]。IL21R是由人類幼稚B細胞、記憶B細胞和生發中心B(GCB)細胞以及漿細胞分泌[16-17],分泌的IL21R又能反向調節B細胞的增殖分化及其功能,可提高B細胞等免疫細胞的免疫功能和活性[18]。IL21與IL21R的結合通過酪氨酸激酶-信號轉導和轉錄激活因子(JAK-STAT)信號通路進行信號傳導[19]。此通路經過激活JAK1與JAK3激酶的催化活化導致IL21R上的酪氨酸殘基被磷酸化,從而為STAT蛋白和其他信號分子提供結合位點[20]。被磷酸化的STAT與同源二聚體或異二聚體發生二聚化后,轉移到細胞核中并通過參與調節靶基因來調控B細胞[21-22]。目前,關于藏羊IL21基因方面研究報道很少,本研究使用PCR方法成功擴增出歐拉型藏羊IL21基因,對其進行了生物信息學分析,此研究結果可為藏羊IL21基因抗病功能研究提供參考依據。
本研究通過分析歐拉型藏羊IL21基因序列和同源性比對結果,發現歐拉型藏羊IL21基因與羊亞科類動物如綿羊、藏羚羊和山羊IL21基因的核苷酸序列同源性較高,均達到99.0% 以上,而與牛亞科動物如美洲草原野牛、牦牛、水牛和瘤牛IL21基因的核苷酸序列同源性低于羊亞科類動物,結果均在98.2%~98.4%之間,說明IL21基因具有一定的種屬特異性,種族越相近,其同源性越高,并且兩者相差僅有1%左右,說明IL21基因具有一定的保守性。胡春榮等[23]發現在小鼠和人類中表達螺旋A和D區時,IL21保守性很高。
蛋白質的卷曲螺旋具有多元化的作用,包括細胞骨架、寡聚化結構域及跨膜信號的調節等。由于肽鏈中的全部肽鍵都可形成氫鍵,因此蛋白質的α螺旋十分穩定,可牢固的維持蛋白的高級結構。β轉角是較為松散的結構,穩定性低。本研究結果表明,在歐拉型藏羊IL21蛋白主要由α螺旋組成,占53.5%,表明 IL21蛋白穩定很好;且歐拉型藏羊IL21蛋白的77~115 區域親水性指數高和蛋白表面可及性較大,這為研究藏羊IL21蛋白的免疫功能提供了理論依據。
蛋白質的翻譯后修飾在生命活動中起著重要作用,可以讓蛋白質的構成更豐富,使其功能具有多樣性和完整性。經三級結構預測顯示IL21蛋白含有酰胺化修飾位點和亮氨酸拉鏈區域。其中酰胺化修飾在蛋白質翻譯中比較常見,是蛋白質翻譯后修飾種類中的第4大類型,由于其發生于肽的C末端,所以也被叫做C-末端酰胺化。酰胺化修飾在動物神經系統和內分泌系統中普遍存在,超過一半的活性肽都存在此過程,在很多神經肽的生物活性中起著重要作用[24]。酰胺化不但可以作為生物活性肽與受體識別和信號轉導過程中的重要因子,可能在提高多肽與受體親和力方面具有顯著作用,而且還與多種疾病如癌癥和高血壓等有關[25-26]。亮氨酸拉鏈 (BZIP) 蛋白是真核生物的轉錄因子和阻遏蛋白中最大且最保守的家族之一,在哺乳動物等真核生物中都發揮重要作用[27-28],如在哺乳動物中具有抑制或者促進基因的翻譯的作用。亮氨酸拉鏈越短其靈活可變性就越小,穩定性越高[29];然而較長的拉鏈是為了能與特定配體結合,有著更好的二聚體特異性,但是穩定性較差。有些亮氨酸拉鏈的N端包含一些額外的DNA結合分子,這樣可以增加DNA的穩定性[30]。蛋白質翻譯后修飾對其蛋白的結構與功能具有重要意義,所有蛋白質都需要經過翻譯后修飾,其結構更加復雜功能更加完整,才可表現出正常的生物學活性。
4 結 論
歐拉型藏羊IL21基因長度為387 bp(無終止密碼子),與參考綿羊、藏羚羊、山羊、美洲草原野牛、牦牛、水牛和瘤牛的IL21基因核苷酸序列同源性依次為100%、99.5%、99.0%、98.4%、98.4%、98.2%和98.2%;氨基酸序列同源性依次為100%、100%、100%、98.4%、98.4%、98.4% 和97.7%,藏羊與綿羊的親緣性最高。IL21蛋白主要由α螺旋組成,是一種親水性蛋白,含有酰胺化位點和亮氨酸拉鏈模式兩種蛋白功能修飾位點。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
財經界(學術版)(2015年20期)2015-12-23 09:20:13
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
