長白山北坡云冷杉闊葉混交林的物種多度格局*
2021-07-16 03:12:26郭躍東張會儒王卓暉
林業科學 2021年5期
郭躍東 張會儒 盧 軍 王卓暉
(1.中國林業科學研究院資源信息研究所 北京 100091; 2.山西農業大學 太谷 030801; 3.國家林業和草原局森林經營與生長 模擬重點實驗室 北京 100091; 4.吉林省汪清林業局 汪清 133200)
群落格局與動態過程的關系是群落構建研究的關鍵內容(Vellend, 2010; Roughgarden, 2009),森林植物群落的空間格局往往是過去生態過程作用的結果,由格局反演過程已成為當前群落生態學研究的主要手段(McGilletal., 2007; Matthewsetal., 2014)。但群落裝配的生態學格局和過程并不是一一對應的,相同的格局可能由不同的生態過程導致(Huetal., 2006; Stanleyetal., 2006), 且這些過程也并非是相互排斥的(周淑榮等, 2006; 牛克昌等, 2009)。物種多度格局是群落裝配理論的重要結構指標(McGilletal., 2007),通過物種多度關系模型的比較,可以指示群落的演替狀態(施建敏等, 2015)、檢測生態系統的干擾與破壞(Hilletal., 2002)、解釋物種的資源分配和種間關聯(Chase, 2005; Sugiharaetal., 2003)、闡明物種分布的環境篩選機制(石培禮等, 2000; 韓大勇等, 2014; 張姍等, 2015)以及充實生態位模型、群落中性模型及其連續體理論體系(Hubbell, 2006),為生物多樣性和穩定性評價(張立敏等, 2010)及能流、進化、生產力等功能特性關系(Legovic, 1991)研究提供思路。物種多度關系模型主要包括經驗統計模型、生態位模型和群落中性理論模型(Pootetal., 2003; 馬克平等, 1997),其中Hubbell(2001)提出的中性理論模型引發廣泛爭議(Chisholmetal., 2010; 周淑榮等, 2006; Volkovetal., 2003),爭議的焦點是對經驗數據格局的模擬哪種模型最佳(Alonsoetal., 2006; McGilletal., 2006),且在爭議中不斷繁衍出新的模型(McGilletal., 2007),朝著聯合理論的方向發展(王緒高等, 2008)。目前,國內群落多度格局的報道較多,森林生態系統因其復雜性成為群落多度格局研究的重要方面(馮云等, 2007),且有關長白山典型森林的群落多度格局研究也較多(王緒高等, 2008; 張姍等, 2015; 閆琰等, 2012),但大多數研究仍是經驗統計模型、生態位模型和群落中性理論模型的對比分析,復合模型以及過伐林群落多度格局的研究尚未見報道。
鑒于此,本研究以長白山北坡云冷杉闊葉混交過伐林為研究對象,通過經驗統計模型、生態位模型和群落中性理論模型的對比分析,構建過伐林喬木層物種多度格局生態位復合模型,闡明過伐林群落冠層結構維持的生態過程,探討生境異質性、密度依賴、種間關聯、擴散限制、更新限制和物種分化在針闊混交林群落裝配中的作用,以期為揭示過伐林恢復過程中的物種多度分布機制提供理論指導。
1 研究區概況
研究區位于吉林省汪清林業局金溝嶺林場,屬長白山系老爺嶺山脈雪嶺支脈,地理坐標為130°05′—130°20′E,43°17′—43°25′N。林區地貌屬低山丘陵,海拔 300~1 200 m,坡度主要集中在5°~25°,個別陡坡在35°以上。研究區屬季風性氣候,1月最低平均氣溫-32 ℃,7月最高平均氣溫32 ℃,全年平均氣溫4 ℃; 年降水量 600~700 mm,多集中在7月。土壤多為灰棕壤土。主要樹種有魚鱗云杉(Piceajezoensisvar.microsperma)、紅皮云杉(P.koraiensis)、臭冷杉(Abiesnephrolepis)、沙松(A.holophylla)、黃花落葉松(Larixolgensis)、紅松(Pinuskoraiensis)、楓樺(Betulacostata)、白樺(B.platyphylla)、紫椴(Tiliaamurensis)、白榆 (Ulmuspumila)、裂葉榆(U.laciniata)、大青楊(Populusussuriensis)、水曲柳 (Fraxinusmandshurica)、東北紅豆杉(Taxuscuspidata)、色木槭(Acermono)、青楷槭(A.tegmentosum)、花楷槭(A.ukurunduense)、朝鮮槐(Maackiaamurensis)等。
2 研究方法
2.1 樣地設置與調查
2013年,在研究區設置 12 塊面積為1 hm2(100 m×100 m)的固定樣地,檢尺所有胸徑 1 cm 以上林木的特征值,包括樹種、胸徑、樹高、枝下高和冠幅等。按相鄰格子法將樣地劃分成 100 個 10 m×10 m 的小樣方,用皮尺定位每株林木的橫縱坐標,并于2016和2018年進行復測調查。本研究采用2013年樣地初始調查數據,根據主林層優勢樹種(組)林木生長情況及對應樹種的生長過程得知,研究區樣地內林分處于中齡林到近熟林階段,樣地基本概況見表 1。研究區林分在中華人民共和國成立前后經過多次采伐干擾,森林蓄積量明顯下降, 1998年禁伐后進入次生林恢復經營階段,此后未采取任何經營措施, 2016年開展目標樹撫育間伐,撫育強度為5%~20%。

表1 樣地基本概況①Tab.1 Basic characteristics of the twelve sample plots
2.2 物種多度模型擬合與檢驗
對12塊樣地喬木層樹種多度數據分別構建對數級數模型、對數正態模型、泊松對數正態模型等經驗統計模型,幾何級數模型、斷棍模型、Zipf模型、Zipf-Mandelbrot模型、生態位重疊模型、優勢優先模型、隨機分配模型等生態位模型,集合群落零和多項式模型和Volkov模型等群落中性理論模型以及復合模型, 擬合物種多度格局。
2.2.1 經驗統計模型擬合 采用對數級數模型(logarithm series distribution,ls)(馬克平等, 1997)、對數正態模型(logarithm normal distribution,ln)(石培禮等, 2000)和泊松對數正態模型(Poission logarithm normal distribution, pl)對12塊樣地物種多度分布進行擬合,泊松對數正態模型認為集合(或樣本)中1個種包含r個個體的概率為:
(1)
式中: 假設lnλ遵從均值和方差分別為M和V的正態分布;r=1,2,…;λ為泊松分布均值,即物種出現的平均多度(Bulmer, 1974)。
2.2.2 生態位模型擬合 采用幾何級數模型(geometrical series model,GS)、斷棍模型(random broken sticks model, RBS)、Zipf模型(Zipf)、Zipf-Mandelbrot模型(Mand)、生態位重疊模型(overlapping niche model,ONM)、優勢優先模型(dominance preemption model, DPM)和隨機分配模型(random assortment model, RAM)對12塊樣地物種多度分布進行擬合,具體公式見參考文獻(馬克平等, 1997; 任萍等, 2009; 趙天啟等, 2017)。
2.2.3 中性理論模型擬合 1) 集合群落零和多項式模型(metacommunity zero sum polynomial model,MZSM) 該模型假定某一樣點的物種多度分布來自于中性復合群落的隨機漂變(Alonsoetal., 2010),復合群落中任一取樣點內多度為n的物種數S可表示為:
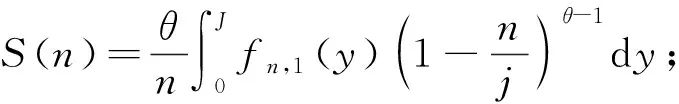
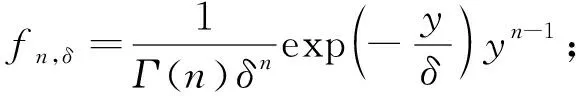
(2)

式中:J為取樣點內的個體數;θ為基本生物多樣性指數。對數級數模型是該模型的一個特例(Hubbell, 2001),二者擬合效果非常相似。
2) Volkov模型 該模型在擬合群落物種多度分布時,相較于MZSM模型增加了遷移系數(immigration rate,m)(Volkovetal., 2003),并假定此遷移系數在物種從復合群落到局域群落的過程中是不變的(張姍等, 2015)。根據該模型,局域群落內多度為n的物種數S可表示為:

(3)
式中:γ為遷移到局域群落的個體數(Volkovetal., 2003; 張姍等, 2015);m為遷移系數。
2.2.4 復合模型擬合 群落是指特定生境中共同生活的許多生物種群的集合,復合模型源于1個群落存在2種或多種構建規律,可能最合理的情形是,優勢度最前面幾個豐富種(常見種)符合生態位模型,后面幾個稀有種符合隨機分配模型,需要組合生態位分割或重疊和隨機生態位分配等模型。Tokeshi(1990)在進行搖蚊(Polypedilum)群落的多度分析時就采用了復合模型。
2.2.5 斷面積廣義多度模型擬合 由于多度格局僅代表物種多寡的分布,而與森林測定密切相關的胸高斷面積更能反映出物種生態位或資源占據狀況(Picardetal., 2002),因此本研究以胸高斷面積作為廣義多度指標進行物種廣義多度格局分析。目前,物種多度格局研究主要限于以物種個體數作為物種多度指標而忽略了物種個體特征(劉夢雪等, 2010),如生物量、高度、蓋度等,已有研究證明生物量作為物種多度指標的分布不同于以群落物種個體數為指標的分布(Morlonetal., 2009)。本研究通過上述幾種多度模型擬合對比斷面積廣義多度與傳統多度指標的差異。
2.2.6 物種多度模型檢驗 1) 統計檢驗 McGill(2003)認為,采用1種模型擬合度檢驗不能很好判定模型擬合效果好壞,并提出了幾種在統計上證明曲線擬合優于零假設的檢驗方法。本研究采用 Kolmogorov-Smirnov(K-S)檢驗統計量(D)和卡方檢驗統計量(χ2)進行物種多度模型檢驗。
2) 赤池信息準則 赤池信息準則(Akaike information criterion,AIC)是在假設模型誤差服從獨立正態分布的情況下,為避免過擬合,在模型對數據擬合的優良性與模型的復雜性之間進行權衡,選擇AIC最小的模型為最優模型的一種信息標準(Kennethetal., 2002)。公式如下:
(4)
式中:L為估計模型的似然函數最大值; RSS為剩余平方和;n為樣本數;p為參數個數。AIC兼顧模型的簡約性和預測的最優性,可避免模型參數冗余。
3) 包絡線檢驗 群落中性理論模型采用隨機生成的多次模擬數據進行包絡線檢驗。具體步驟如下: 首先,運用R語言sads包獲得群落中性理論模型的基本參數; 然后,運用untb包進行600次多度隨機模擬預測分布,計算各排序物種的多度期望值和95%置信區間,取第 2.5%(第 15個)和第 97.5%(585個)作為 95%置信區間的上下限; 最后,通過對比實際觀察多度與模擬多度包絡線的關系,分析群落中性理論模型的擬合狀況(Walkeretal., 2007)。
2.2.7 數據處理 本研究所有統計分析與模型擬合、檢驗均采用R-3.4.1中的sads和untb包完成。Shannon-Wiener多樣性指數、Marglef豐富度指數、Simpson優勢度指數和Pielou均勻度指數計算采用vegan包完成,計算公式見參考文獻(馬克平等, 1997)。
3 結果與分析
3.1 經驗統計模型擬合
采用對數級數模型(ls)、對數正態模型(ln)和泊松對數正態模型(pl)對12塊云冷杉闊葉混交過伐林樣地喬木層物種多度分布進行擬合,其擬合檢驗統計量見表2。

表2 物種多度格局擬合的經驗統計模型檢驗統計量①Tab.2 Empirical statistical model test of species abundance pattern
由表2可知,從擬合檢驗統計量χ2通過比例看,對數級數模型好于對數正態模型和泊松對數正態模型,而從D和AIC指標看,對數正態模型和泊松對數正態模型好于對數級數模型。樣地1、3、4未通過χ2顯著性檢驗,樣地2未通過K-S檢驗,表明這些群落采用經驗統計模型擬合效果不佳。圖1中樣地7和1物種多度分布的經驗統計模型擬合對比也得到相似結論,樣地7擬合效果較好,樣地1較樣地7擬合效果差一些。表2還表明,D和AIC統計量在群落多度格局擬合檢驗中表現一致,而χ2檢驗相對較差。
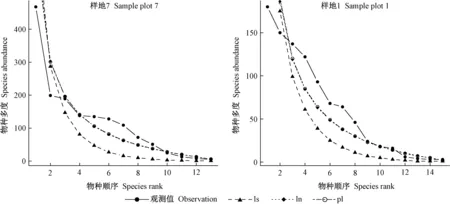
圖1 物種多度格局的經驗統計模型擬合Fig. 1 Fitting curve of empirical statistical model of species abundance pattern
3.2 群落多樣性參數
對數級數模型參數T和對數正態模型參數λ可作為表征群落多樣性的重要參數,以個體數和胸高斷面積為指標的Shannon-Wiener多樣性指數、Marglef豐富度指數、Simpson優勢度指數和Pielou均勻度指數計算結果見表3,經驗統計模型參數與群落多樣性參數之間的相關性分析見表4。

表3 物種多度分布擬合的經驗統計模型多樣性參數與群落α多樣性參數Tab.3 Biodiversity parameters of fitting species abundance model and alpha biodiversity parameters
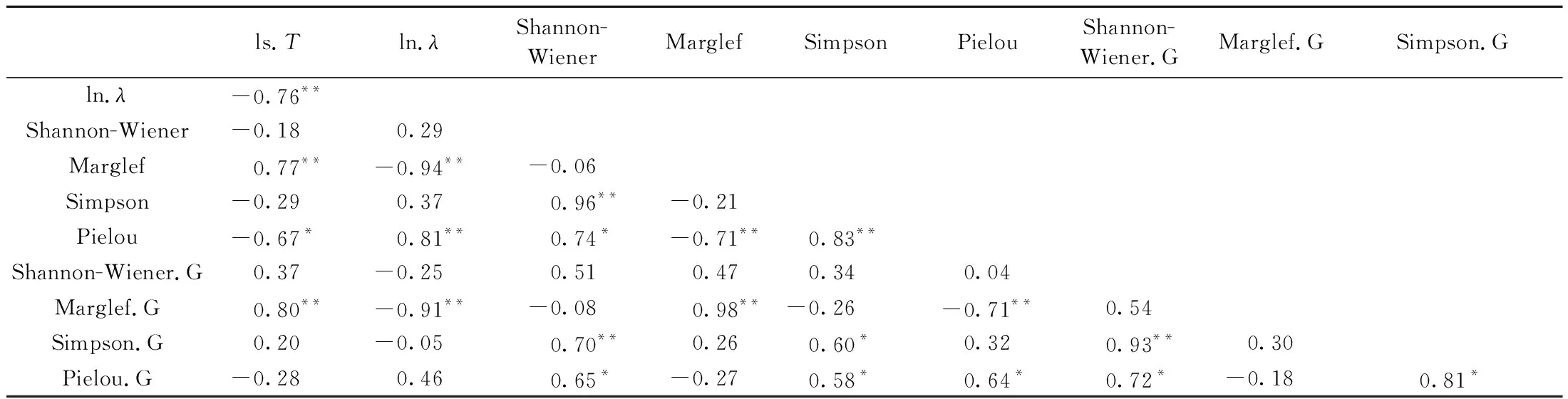
表4 物種多度分布擬合的經驗統計模型多樣性參數與群落多樣性參數相關性分析①Tab.4 Correlation analysis between biodiversity parameters and fitting parameters from empirical statistical models
由表3可知,除樣地12外,以胸高斷面積為指標的群落多樣性參數明顯高于以個體數為指標的群落多樣性參數,表明群落中稀有種胸徑較大,且大多為優勢林冠層的重要伴生種,也可能是過伐林殘留林冠層的組分。由表4可知,參數T與λ呈顯著負相關,參數T與Marglef豐富度指數呈顯著正相關、與Pielou均勻度指數呈顯著負相關、與Shannon-Wiener多樣性指數和Simpson優勢度指數無顯著相關性。參數λ與群落多樣性參數的關系類似于參數T,但符號相反。Shannon-Wiener多樣性指數與Simpson優勢度指數、Pielou均勻度指數呈顯著正相關,Marglef豐富度指數與多度的Pielou均勻度指數呈顯著負相關、與斷面積的Pielou均勻度指數不相關,Simpson優勢度指數與Pielou均勻度指數呈顯著正相關。
3.3 生態位模型擬合
采用幾何級數模型(GS)、斷棍模型(RBS)、Zipf模型(Zipf)、Zipf-Mandelbrot模型(Mand)、生態位重疊模型(ONM)、優勢優先模型(DPM)和隨機分配模型(RAM)對12塊云冷杉闊葉混交過伐林樣地喬木層物種多度分布進行擬合,其擬合檢驗統計量見表5。各模型對群落物種多度分布的擬合及實際觀測數據見圖2。
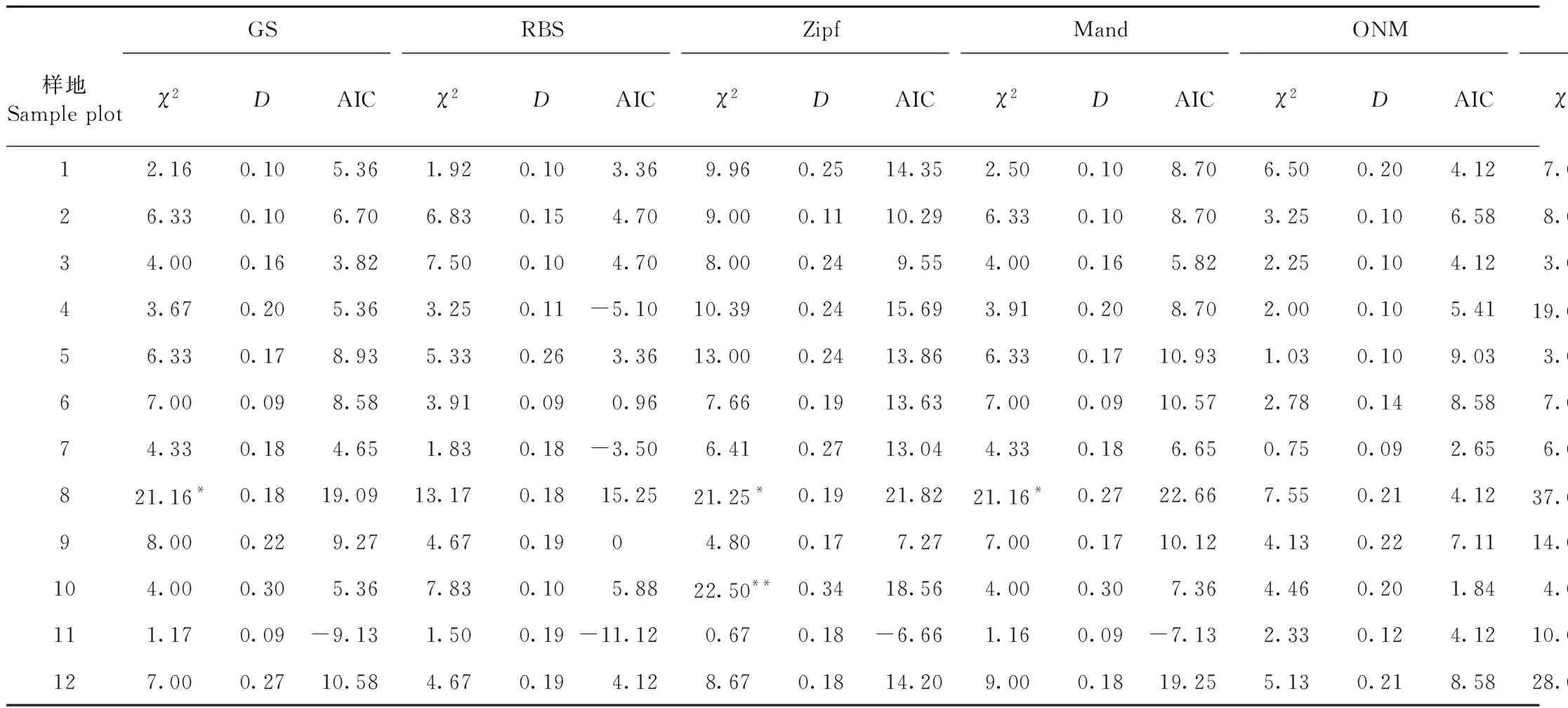
表5 物種多度格局的生態位模型檢驗統計量Tab.5 Niche model test of species abundance pattern

圖2 物種多度格局的生態位模型擬合Fig. 2 Fitting curve of niche model of species abundance pattern
由表5可知,不同生態位模型對群落物種多度分布的擬合存在較大差異,按照擬合優良性表現指標,斷棍模型擬合效果最佳,其次是生態位重疊模型和幾何級數模型,再次為Zipf模型和Zipf-Mandelbrot模型,最后為隨機分配模型和優勢優先模型(圖2)。采用斷棍模型、生態位重疊模型和幾何級數模型,所有群落優勢度多度曲線均具有一個明顯的擬合陡變點,在重要性排序軸的前半段,表征群落多度均勻性的曲線陡度較緩,3種模型的擬合效果均較好,尤其是生態位重疊模型,而在重要性排序軸的后半段,曲線急劇下降,3種模型的擬合效果均較差(圖2),表明群落是由5~8個明顯的優勢種(豐富種)組成的群落,群落物種多度格局可能存在2種規律,豐富種和稀有種多度呈2個明顯的競爭等級,采用復合模型可能具有更高的擬合優良性。
3.4 中性理論模型擬合
采用零和多項式模型(MZSM)和Volkov模型(Volkov)對12塊云冷杉闊葉混交過伐林樣地喬木層物種多度分布進行擬合,其擬合檢驗統計量見表6。群落物種多度分析的Volkov模型包絡線檢驗見圖3。
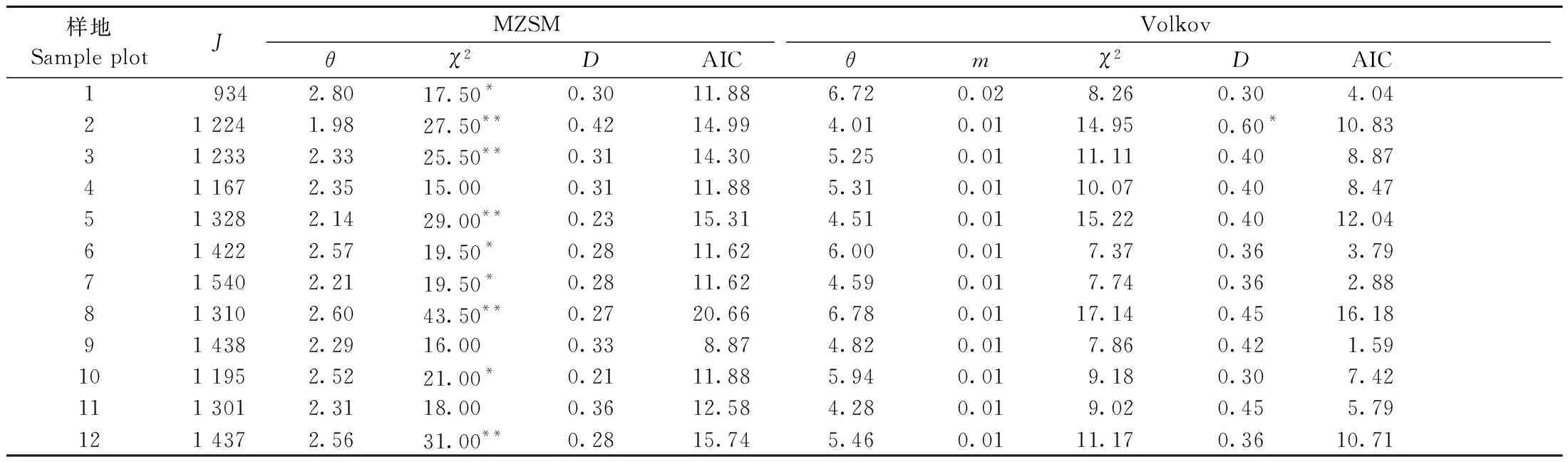
表6 物種多度格局的中性理論模型檢驗統計量①Tab.6 Neutral theory model test of species abundance pattern
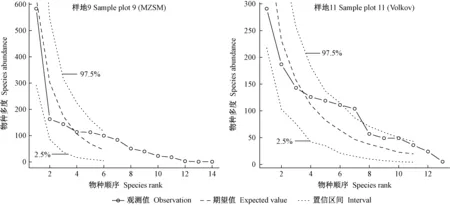
圖3 物種多度格局的中性理論模型包絡線檢驗Fig. 3 Envelop test of synthetic data from different neutral theory model
由表6 可知,所有樣地零和多項式模型的χ2統計量均大于15,除樣地4、9、11外,其他樣地的零和多項式模型均未通過顯著性檢驗,而所有樣地的Volkov模型均通過統計檢驗,表明Volkov模型可用于擬合群落的物種多度格局,但其χ2明顯高于生態位模型,即Volkov模型擬合并不是最佳的。圖3群落中性理論模型包絡線檢驗結果也表明Volkov模型優于MZSM模型。樣地9和11的大多數物種多度觀測值明顯偏離中性理論期望值,重要性排序居中的物種多度甚至達到或超出600次隨機模擬的97.5%包絡線(群落11表現尤甚),少數物種的多度也達到或超出2.5%置信區間(圖3),且少數稀有種或伴生種的多度預測為0,無法體現群落生物多樣性狀況,尤其是MZSM模型。這表明群落中性理論模型用于物種優勢度多樣性曲線擬合相對較差,換言之,群落中各物種的多度分布并不是獨立、隨機的,而是受某種特定的生態位過程機制制約。
3.5 復合模型擬合
由上述分析可知,經驗統計模型中對數正態模型和泊松對數正態模型較對數級數模型擬合效果好,可能與群落中中間倍程的物種數較多、優勢種或稀有種的物種數較少有關(圖1)。生態位模型擬合效果較經驗統計模型好,可能是因為群落中存在明確的生態位過程,制約群落資源分配,影響物種多度,但觀測值具有明顯的拐點特征,表明群落物種多度格局存在2種或2種以上規律。本研究采取以下步驟進行復合模型擬合: 首先,從物種多度觀測曲線上(圖2)找出陡度明顯變化陡變點的物種序號,將多度數據分為前后2個數據集進行分段擬合,所有群落陡變點前物種數據趨向于各物種多度分配均衡狀態,采用斷棍模型和生態位重疊模型可能更能反映該情況,而陡變點后物種數據趨向于多度分配明顯失衡狀態,采用隨機分配模型、優勢優先模型和斷棍模型可能更能反映該情況; 其次,對2個數據集分別采用前述參試模型組成6種復合模型進行群落物種分布擬合,得到最佳復合模型擬合方法。在分段擬合時,涉及最優陡變點的問題,觀察記錄明顯陡變點的物種序號,前后推移各3個序號,采用Lai等(2013)的最大似然法確定最佳臨界值。本研究利用AIC最小值獲得復合模型擬合陡變點最佳序號,以方便比較各復合模型的表現。表7所示為物種多度分布數據的復合模型擬合結果,圖4所示為群落物種序列-多度格局實例。

圖4 6種復合模型擬合物種多度格局實例Fig. 4 Examples from the six kinds of composite model fitting to species abundance pattern
由表7可知,大多數群落陡變點的物種序號為8~10,表明云冷杉闊葉混交林是由7~10個物種組成的多優群落,這從表1中樣地樹種組成也可明顯看到,優勢種多度占整個群落多度的87%~99%,且多度分配較為均衡。不同數據集都以生態位重疊模型、隨機分配模型、斷棍模型和優勢優先模型組合的復合模型擬合效果最佳,可見生態位重疊模型、隨機分配模型、斷棍模型和優勢優先模型組合可能是云冷杉闊葉混交過伐林物種多度分布擬合的最佳復合模型。另外,復合模型檢驗指標除樣地8外均通過擬合優度卡方和K-S檢驗,但χ2和D較生態位模型沒有明顯變化,AIC明顯下降,表明復合模型擬合效果更好。由圖4可知,復合模型的預測值與觀測值更加接近,模型擬合的優良性有較大提高。2種模型的銜接部位可能會出現違背物種序列-多度關系曲線的現象(圖4右圖),但由于陡變點前物種多度占總多度的87%~99%,因此這種擬合不會嚴重違背多度分布曲線。

表7 物種多度分布數據的復合模型擬合①Tab.7 Composite model fitting of species abundance distribution
3.6 斷面積廣義多度模型擬合
以胸高斷面積作為廣義多度指標進行群落物種多度格局擬合的結果如圖5所示(以樣地1為例)。
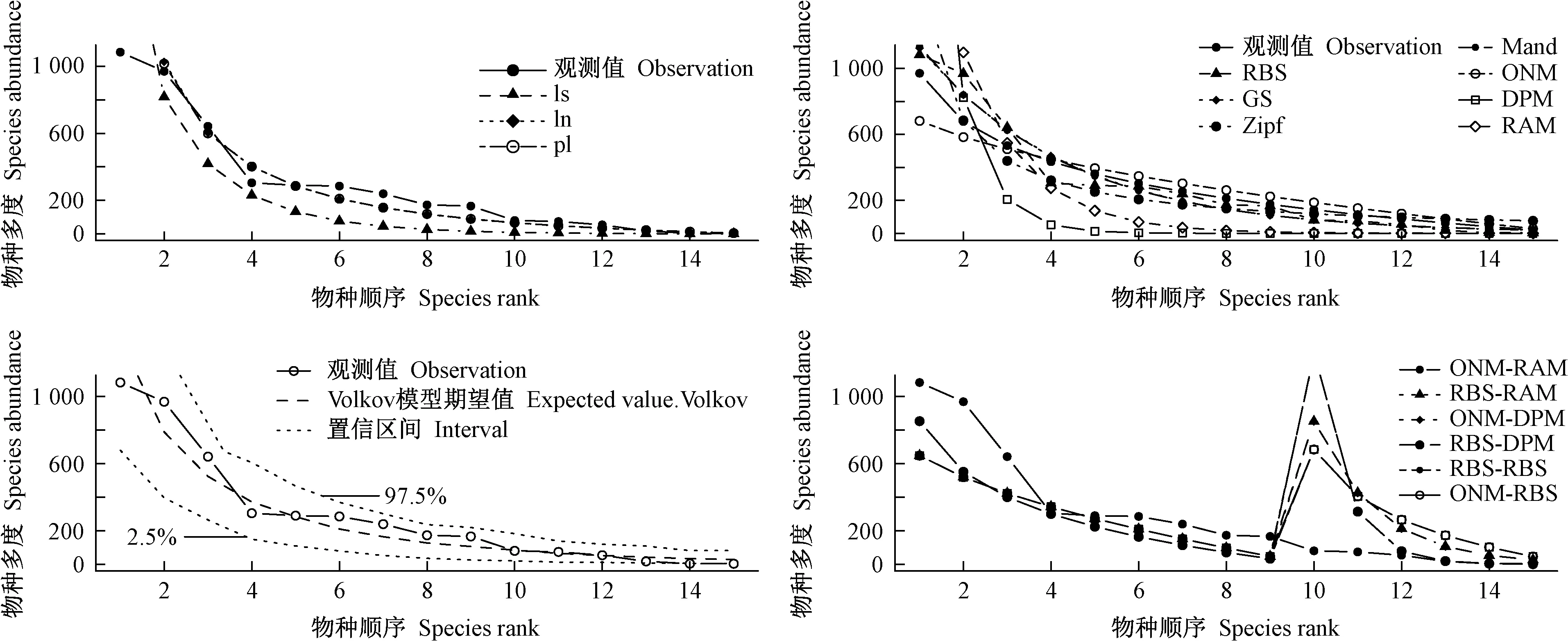
圖5 斷面積廣義多度物種多度格局的經驗統計模型、生態位模型、中性理論模型包絡線檢驗和復合模型擬合(樣地1)Fig. 5 Fitting of species abundance pattern for sample plot 1 basal area as abundance indices of different models
由圖5可知,采用胸高斷面積廣義多度指標與采用數量多度指標進行群落物種多度格局擬合的經驗統計模型、生態位模型擬合結果基本一致,但中性理論模型Volkov包絡線檢驗擬合、復合模型表現與采用數量多度指標的相應模型截然不同,群落胸高斷面積的多度分布更多表現為隨機中性理論模型,復合模型的表現較單一模型差,尤其在陡變點附近,這可能是因為稀有種的數量多度所占比例雖然較低,但稀有種多數情況下在林分中為優勢木或亞優勢木,胸高斷面積所占比例較高,胸高斷面積的分配在各物種之間更加均勻,因此采用胸高斷面積作為廣義多度指標的復合模型擬合不能嚴格區分豐富種和稀有種,會導致競爭等級層次的模糊化或缺失化。
4 討論
4.1 經驗統計模型擬合情況
研究區云冷杉闊葉混交過伐林生境優越,種源豐富,林分異質性和多樣性較高,為物種的抽彩式競爭奠定了基礎,各物種多度分配較為均衡,優勢種和稀有種數量均較少,大多數物種多度處于平均水平,采用對數正態模型擬合效果最佳,與趙志模等(1990)的研究結論一致。在擬合物種多度格局方面,泊松對數正態模型和對數正態模型擬合度一致,可采用對數正態分布去近似泊松對數正態分布(Blissetal., 1953)。在參數相關性方面,對數級數模型參數T和對數正態模型參數λ與群落Marglef豐富度指數和Pielou均勻度指數具有明顯相關性,表明這2個參數能較好反映群落物種豐富度和物種多樣性(馬克平等, 1997; 任萍等, 2009)。
4.2 機制模型擬合情況
相較于經驗統計模型,生態位模型更能反映生境優越度(高利霞等, 2011)、異質性(劉夢雪等, 2010)、群落演化歷史、物種組成的豐富程度、物種資源占據能力(彭少麟等, 2003)、生態位空間動態及其等級多度分布(Tokeshi, 1990),可能是反映群落多度與資源分布正、負或無相關的更好模型(韓大勇等, 2014)。復合生態位模型認為群落物種多度格局不止遵循1種規律,最可能出現的情況是幾個多度最大的種符合生態位分割模型,而其余多度較小的種符合隨機分配模型(Tokeshi, 1990)。長白山云冷杉闊葉混交林資源豐富,豐富種多度配置較為均衡,采用斷棍模型和生態位重疊模型擬合效果最佳,復合模型中生態位重疊模型在豐富種多度擬合時也表現最佳,稀有種所占比例較少,且多度分配是隨機的,采用隨機分配模型、斷棍模型和優勢優先模型擬合效果較好,其中尤以隨機分配模型最佳,與Tokeshi(1990)預測基本一致。
復合生態位模型對豐富種(數據集1)和稀有種(數據集2)2個多度等級的分段擬合效果較單一模型好,表明豐富種和稀有種2個競爭等級在群落結構構建機制方面存在差異。前者更加符合重疊生態位理論,后者更加符合隨機生態位分配理論,說明豐富種和稀有種在群落裝配中屬于2種機制,前者體現了生態位模型的確定性機制,后者體現了隨機性機制,2種機制在過伐林群落構建中是共存的。物種相對頻次分布曲線由豐富種和稀有種2部分組成(Frontier, 1985),Magurran等(2003)在研究魚類群落時也發現豐富種和稀有種(文中稱為持久種或核心種和機會種)的群落構建經驗統計模型存在差異,證明了群落構建的等級機制。隨機分配模型認為物種生態位分配是非等級物種多度分布格局的動態變化(Tokeshi, 1990),這可能是其在群落整體擬合中表現較差(圖2)、在稀有種擬合中表現較佳(圖4)的原因之一。另外,采用群落中性理論Volkov模型對過伐林物種多度格局進行擬合雖然通過檢驗,但其擬合表現稍遜于生態位模型,可見生態位過程在群落構建中的重要作用,而稀有種的生態位隨機分配可能成為群落中性理論的弱化表現,中性理論被認為是當種間適應性無差異且無穩定生態位分化時的特例(Bar-Massadaetal., 2014; Matthewsetal., 2014),Hubbell(2001)采用數學理論證明對數級數分布是零和多項式模型的一個特例,這些研究表明經驗統計模型和機制模型并不是孤立的,任何群落都是群落連續體理論的一個“點”,采用復合模型分析可能闡釋群落構建的生態位和中性理論機制過程在群落多度格局方面的體現,成為揭示各模型連續性的數學途徑。
胸高斷面積廣義多度擬合實際上是傳統物種多度格局的加權擬合方法,其權重為各物種平均胸徑的平方,即將林木大小指標作為權重實現對物種多度格局的量化分析,擬合結果與傳統物種多度格局擬合雖存在差異,但未能總體改變物種多度格局的分配情況。胸高斷面積作為森林生產的功能指標(Picardetal., 2002),可能更加有利于進行森林功能多樣性分析,擴展物種多度格局的功能特性闡釋。
本研究的復合模型組成機制可能略有違背物種序列-多度原則,尤其是陡變點之后1~2個物種的多度可能超過陡變點物種的多度(圖5),這可能與隨機分配、優勢優先和斷棍3種模型在稀有種擬合方面給第一物種優先分配更多多度有關,其改善可能需要提出更為恰當的機制模型。
4.3 過伐林物種多度分布成因分析
干擾是塑造群落結構、影響群落裝配過程的重要因子(Podlaski, 2017),林分過伐干擾后形成斷層,先鋒物種隨機侵入,萌生物種原位更新,二者很快進入主林層,并與殘留林冠層形成共優群落,隨機干擾采伐事件的空間多尺度性和時間離散性增加了群落異質性和多樣性(Podlaski, 2010),種源擴散、物種競爭、森林采伐、隨機干擾等多種機制共同構建了過伐林群落。研究區云冷杉闊葉混交過伐林目前處于恢復演替階段,林分異質性為多物種侵入和生態位占據能力奠定了基礎,各物種多度均衡的生態位模型如生態位重疊模型更能較好擬合當前群落狀態,而到達演替頂極階段,群落物種組成和數量比例趨于穩定,種間競爭等生態位過程并不明顯(閆琰等, 2012; 張姍等, 2015),物種多度差異較大的隨機生態位模型或中性隨機理論模型則可能占據主導(施建敏等, 2015; 任萍等, 2009; 徐國瑞等, 2017),有利于群落定向演替的發生發展。姚蘭等(2016)研究發現,干擾機制下萌生物種是影響群落物種多度格局的重要因素,過伐狀態下萌生樹種的大量出現(如紫椴、色木槭等)可能也是導致群落物種多度分布均衡的機制。先鋒物種迅速侵占采伐跡地和快速生長進入主林層的本領(張會儒等, 2008),是當前林分物種多度分配較為均衡的重要原因。殘留林冠層的衰老、枯立、風倒機制和更新層的替代占位、競爭機制也是決定過伐林群落物種裝配狀態的重要生態過程(Kraftetal., 2008),2種以上的生態位過程機制在決定群落裝配中發揮重要作用(Hollingsworthetal., 2013)。
關于長白山溫帶森林群落的多數研究認為,群落生境優良,同質性高,環境因子的過濾篩選作用較低,種間作用并不強烈,表現為隨機生態過程占據主導,擴散限制作用較高(Zhangetal., 2009; Yuanetal., 2011; Wangetal., 2011),而生態位作用較低(張姍等, 2015)。本研究對過伐林群落多度格局過程的分析表明,云冷杉闊葉混交過伐林群落處于恢復演替階段,高生境異質性和干擾事件隨機性分別決定了生態位理論和中性理論2種機制在過伐林群落構建中的作用,同時受擴散機制、生態位競爭、干擾過程等影響,過伐林多優群落處于均衡多度分布格局中,演替后期云杉、冷杉和頂極伴生種的物種比例與演替前期闊葉樹種的比例相當,演替頂極針葉樹種紅松受種源限制很少出現,東北紅豆杉、黃蘗(Phellodendronamurense)、胡桃楸(Juglansmandshurica)、花楸(Sorbuspohuashanensis)、朝鮮槐等稀有種的物種多度較少,這可能是由于過伐林與長白山溫帶森林處于2個不同的發育階段,過伐林處于演替恢復期,生態位過程占據主導,稀有種的多度和頻度均較少,而長白山溫帶森林處于原始演替頂極狀態,中性隨機理論模型占據主導(Wangetal., 2011),演替后期的稀有種和伴生種比例較高。在過伐林群落次生演替中,喜寒涼氣候的暗針葉樹種與喜溫涼濕潤氣候的闊葉樹種因資源利用、環境適應、干擾應變等方面的差異而在群落中同時存在,且表現出普遍生態位重疊(伏捷等, 2018),但重疊程度較低(鄒春靜等, 2001),這也可能是塑造過伐林物種多度均衡的重要機制。
多數研究表明,物種多度格局受尺度(張忠華等, 2012; 程佳佳等, 2011)、物種空間分布格局(王緒高等, 2008)等的影響。Wang等(2018)對吉林蛟河溫帶林的研究發現,大樹周圍總是被異種的大樹所圍繞,這也可能是制約物種多度格局均衡分配的主要因素。中性理論模型在大尺度(程佳佳等, 2011; 閆琰等, 2012; Fisheretal., 2014; Chengetal., 2012)和物種多樣性較高群落(Chisholmetal., 2010)擬合方面往往較生態位模型表現更好,本研究并未發現該規律。下一步將開展物種空間分布格局與物種多度格局的尺度關聯研究,建立空間明確的多尺度物種多度格局模型,闡明不同尺度上控制格局的生態過程和機制,揭示物種空間分布與多度格局之間的關系。
4.4 過伐林經營意見
上述群落物種多度格局分析表明,長白山云冷杉闊葉混交過伐林群落處于恢復演替階段,通過經營干預可有效減少其演替發育時間。針對林分特點,在過伐林經營中要注意以下幾點: 1) 減少先鋒軟木闊葉樹種比例,增加伴生硬木闊葉樹種比例; 2) 通過林下補植提高紅松比例,間伐林冠上層減少云冷杉比例,提高目標樹單木生長量,減弱風倒危害,影響云冷杉更新層,降低云冷杉聚集規模; 3) 保留部分上層霸王木、老狼木,保留和促進珍貴樹種如東北紅豆杉和三大硬闊的更新; 4) 采用近自然經營方法(陸元昌等, 2011)、森林生態系統采伐技術體系(張會儒等, 2008)、可變綠樹保持法(Maríaetal., 2011; Laurenetal., 2013),使云冷杉闊葉混交林林分逐步向紅松云冷杉林過渡,促進結構發育的多層性、異質性和復雜性,穩定和提高林分多樣性和生產力,提升林分的健康穩定性。
5 結論
1) 經驗統計模型中對數正態模型和泊松對數正態模型擬合效果最佳,對數級數模型擬合效果較差,對數級數模型參數T和對數正態模型參數λ可作為表征群落多樣性的重要參數。
2) 生態位模型中斷棍模型擬合效果最佳,其次是生態位重疊模型和幾何級數模型,再次為Zipf模型和Zipf-Mandelbrot模型,最后為隨機分配模型和優勢優先模型。
3) 復合生態位模型在物種多度分析中擬合效果較傳統生態位模型有較大提高,復合模型可能在群落豐富種和稀有種呈現競爭等級多度分布時擬合效果最佳。
4) 復合模型可能成為連接經驗模型、生態位模型和中性理論模型的紐帶,為研究物種多度格局提供數學途徑和思路。
5) 以胸高斷面積為指標的物種多度分布擬合與以個體數為指標的經驗統計模型和生態位模型物種多度分布擬合表現基本一致,而在群落中性理論模型和復合模型擬合表現方面有一定差異,可能與稀有種斷面積比重較大有關。
6) 云冷杉闊葉混交過伐林群落物種等級多度分布某種程度上是生態位過程和隨機干擾生態過程共同作用的結果。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
英語世界(2023年10期)2023-11-17 09:18:18
保健醫苑(2021年7期)2021-08-13 08:48:02
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
學生天地(2020年36期)2020-06-09 03:12:30
數學物理學報(2020年2期)2020-06-02 11:29:24
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
光學精密工程(2016年6期)2016-11-07 09:07:19
