中國南方雙斑長跗螢葉甲地理種群遺傳結構及Wolbachia感染
2021-07-15 12:34:08張小飛徐玲玲申圓圓李肖肖王振營
昆蟲學報 2021年6期
李 菁, 張小飛, 徐玲玲, 申圓圓, 李肖肖, 王振營
(1. 西安文理學院生物與環境工程學院, 西安 710065; 2. 西安市農業技術推廣中心, 西安 710061;3. 中國農業科學院植物保護研究所, 北京 100193)
雙斑長跗螢葉甲Monoleptahieroglyphica,亦稱雙斑螢葉甲,屬鞘翅目(Coleoptera)葉甲科(Chrysomelidae)螢葉甲亞科(Galerucinae),是一種在我國分布范圍極廣的多食性害蟲,除取食玉米、高粱、棉花、向日葵、豆類等多種作物外,還取食多種林木和雜草,且能在不同寄主植物之間轉移為害(陳靜等, 2007; 張聰, 2012; 劉景皓, 2018)。該害蟲對玉米等多種農作物的為害近年來呈現加重趨勢,為害區域和面積逐年擴張,已成為多地區玉米等作物上的重要害蟲之一(Zhengetal., 2020)。
遺傳多樣性豐富、遺傳結構復雜的昆蟲種群往往更能適應生存環境的變化,在化學藥劑的連續篩選下也更易演化出抗藥性(Xinetal., 2014)。因此,在農業生產中要對害蟲進行有效防治,應先對其田間種群的遺傳多樣性及遺傳結構進行調查和評估。目前DNA測序中最常用的就是線粒體DNA(mtDNA),由于在演化過程中昆蟲的線粒體基因具有選擇壓力小,突變易穩定遺傳,在群體中變異率較高等特點,常被用作系統發育、種群遺傳結構及遺傳分化研究的分子標記(Lynchetal., 2006; Nabholzetal., 2008; 李菁等, 2018b)。在線粒體基因中,細胞色素氧化酶COI和COII基因具有較豐富的變異,在種下階元系統發育及遺傳結構研究中應用尤為廣泛(Liu and Beckenbach, 1992)。目前,國內外關于雙斑長跗螢葉甲及其近緣種屬的種群遺傳學研究較少,分子系統學研究多集中在目、科、亞科和屬的水平上,包括亞科內不同族或屬之間的系統發育關系與螢葉甲部分屬種與寄主植物間的進化關系等(Huntetal., 2007; 陳光輝等, 2016)。通過葉甲科中跳甲亞科、螢葉甲亞科和葉甲亞科共11屬15種昆蟲間的系統進化分析,不同屬均表現為明顯的單系性,證明COII基因是分析葉甲亞科及種屬階元分類與進化的有效分子標記(張高峰, 2006)。而在種內種群水平上,中國北方雙斑長跗螢葉甲不同地理種群之間已產生明顯的遺傳分化,不同種群間基因流水平低,且推測在近期未出現種群擴張現象(梁日霞等, 2011)。雙斑長跗螢葉甲在中國的分布遍及南北方多省區,然而迄今國內外對該甲蟲的種群遺傳學研究僅限于中國北方種群(梁日霞等, 2011)。
本研究以線粒體COII基因作為分子標記,對中國南方地區的雙斑長跗螢葉甲地理種群的遺傳結構及遺傳多樣性進行研究,作為該害蟲南方種群研究數據的補充,并與先前報道的北方種群進行比較分析,同時對南方種群中共生菌Wolbachia進行感染率檢測和感染類群分析,從種群遺傳學水平上探討該害蟲能適應多種寄主植物和氣候環境的內在遺傳因素,為日后對其制定科學有效的防治策略提供理論依據。
1 材料與方法
1.1 供試蟲源
本研究供試的雙斑長跗螢葉甲14個地理種群于2016-2018年間采集自中國南方9個省區(其中,包括陜西南部的兩個種群),采集地均為玉米田。每個種群采集不少于25頭(為便于田間識別,采集蟲態均為成蟲)。采回后單頭成蟲置于離心管中,于-20℃凍存。各種群采樣信息詳見表1。
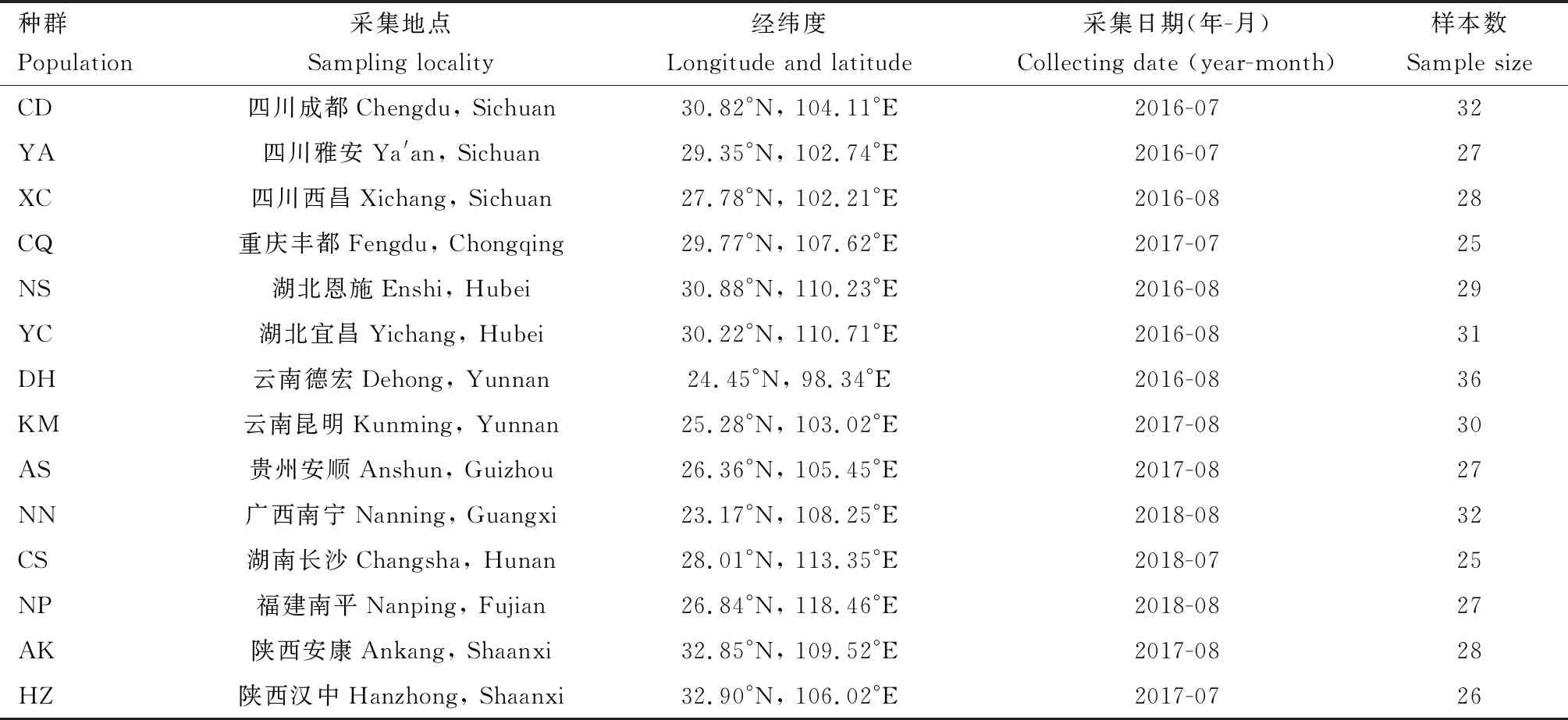
表1 供試中國南方雙斑長跗螢葉甲地理種群樣本采集信息
1.2 基因組DNA提取及PCR擴增
單頭雙斑長跗螢葉甲在1×TE緩沖液(Tris-EDTA,pH 8.0)中冰浴勻漿,應用血液/細胞/組織基因組DNA提取試劑盒(型號DP304,TIANGEN)提取供試甲蟲基因組DNA。提取完畢后通過1.0%瓊脂糖凝膠電泳檢測提取質量,并利用NanoDrop微量核酸/蛋白測定儀(Thermo)測定提取DNA的濃度及純度,檢測合格后保存于-20℃冰箱備用。
雙斑長跗螢葉甲COII基因擴增特異性引物序列參考梁日霞等(2011),PCR反應體系(20 μL): 2×Taq PCR MasterMix 10 μL(TIANGEN),正/反向引物(10 mmol/L)各1 μL, 模板DNA 1 μL, ddH2O 7 μL。PCR反應條件: 95℃ 3 min; 95℃30 s, 53℃ 30 s, 72℃ 1 min, 35個循環;最后72℃延伸10 min。反應結束后每個樣品各取3 μL PCR產物進行電泳檢測(1.5%瓊脂糖凝膠),確認擴增出預期大小的單一片段后,將剩余PCR產物送交生工生物工程有限公司進行純化及雙向測序。
1.3 Wolbachia感染率檢測
通過PCR擴增Wolbachia的wsp基因對供試雙斑長跗螢葉甲種群進行Wolbachia感染率檢測,以實驗室中確定感染Wolbachia的亞洲玉米螟DNA樣本作為陽性對照,以ddH2O作為陰性對照。檢測所用引物為wsp81F(5′-TGGTCCAATAAGTGATGAA GAAAC-3′)和wsp691R(5′-AAAAATTAAACGCTA CTCCA-3′),供試引物由生工生物工程有限公司合成。在每個種群的感染個體中隨機選取10頭進行wsp基因片段PCR擴增和亞克隆,PCR體系、PCR反應程序及亞克隆具體方法參見李菁等(2018a)。委托生工生物工程有限公司進行雙向測序,計算Wolbachia在各供試雙斑長跗螢葉甲種群中的感染率(%)。
1.4 生物信息學分析
通過Chromas軟件讀取1.2節測序數據,并將正反向序列拼接后在NCBI網站中進行BLAST比對,確認為雙斑長跗螢葉甲COII基因再經DNAMAN軟件進行多序列同源比對,為便于與已發表的中國北方雙斑長跗螢葉甲種群COII基因數據進行比較分析(梁日霞等, 2011),最終所有供試個體的COII基因測序樣本均統一截取484 bp序列長度進行后續分析。應用DnaSP6軟件(Rozasetal., 2017)計算單倍型數(h)、單倍型多樣性(Hd)、核苷酸多樣度(Pi)、種群間遺傳分化指數(Fst)和基因流(Nm)等,并進行Tajima’sD和Fu’sFs中性檢驗。結合雙斑長跗螢葉甲中國北方種群的COII數據結果(梁日霞等, 2011),應用Network10.1軟件基于median-joining算法構建單倍型中介網絡圖,并基于鄰接法中的Kimura 2-parameter模型應用MEGA6軟件(Tamuraetal., 2013)構建單倍型系統發育樹,系統樹各分支進行1 000次置信度重復檢驗。應用Arlequin v3.5軟件(Excoffier and Lischer, 2010)進行種群變異的分子方差分析(AMOVA)。通過Genepop在線工具(http:∥www.genepop.curtin.edu.au/genepop_op6.html),對種群間地理距離與遺傳距離進行相關性檢驗。分析Wolbachia感染類群及株系,在PubMLST數據庫中選取具有代表性的Wolbachia株系,與本研究中所測得雙斑長跗螢葉甲種群中感染Wolbachia的wsp基因序列共同構建系統發育樹(最大似然法),建樹時采用MEGA6軟件中的Kimura2-parameter模型,檢驗支持率Bootstrap設置為1 000次重復檢驗。
2 結果
2.1 COII基因變異位點及單倍型
對供試14個地理種群共403頭雙斑長跗螢葉甲個體COII基因片段序列進行整理和多重比對,將兩端存在測序誤差的區段截去,并參考梁日霞等(2011)在GenBank中登錄的中國北方種群COII基因單倍型序列,最終在所有供試個體中均截取484 bp長度序列片段,該COII基因片段中共發現27個變異位點,約占分析區段位點總數的5.6%,并包含2個單變異位點(singleton variable sites)和25個簡約信息位點(parsimony informative sites)。在測序序列中不存在堿基插入/缺失現象,并表現出昆蟲線粒體基因A/T堿基偏倚性的特點(A+T平均含量占76.0%)。
本研究在供試14個地理種群中總共發現23種單倍型,將23種COII單倍型序列分別在NCBI網站進行在線BLAST,發現其中有10種單倍型與梁日霞等(2011)登錄的單倍型序列完全一致,分別命名為MHap14-MHap23(GenBank登錄號分別為HQ909339, HQ909343, HQ909353, HQ909346, HQ909340, HQ909342, HQ909348, HQ909349, HQ909347及HQ909341)。為避免造成數據庫冗余重復信息,將新發現的13種單倍型命名為MHap1-MHap13(GenBank登錄號: MT861137-MT861149);沒有全部種群均有共享的單倍型,至少在2個種群中出現的單倍型有11種,而有12種單倍型僅在1個種群中出現(表2)。
2.2 種群遺傳多樣性及中性檢驗
供試14個雙斑長跗螢葉甲種群的總種群Hd為0.748,Pi為0.00769,核苷酸平均差異數(K)為3.7232。各種群包含的單倍型數(h)在2~6之間,變異位點數(S)在2~20之間,Hd在0.394~0.782,Pi在0.00125~0.01512(表2)。總種群的Tajima’sD和Fu’sFs檢驗均為負值且不顯著,在不同種群中這兩項中性檢驗值既有正值也有負值,且只有NS種群的Tajima’sD檢驗結果為顯著正值(D值為2.0704,P<0.05)(表2)。

表2 基于中國南方雙斑長跗螢葉甲地理種群COII基因序列的遺傳多樣性及中性檢驗
2.3 COII單倍型系統進化
雙斑長跗螢葉甲中國北方種群數據(梁日霞等, 2011)與本研究中中國南方種群的數據相結合,構建COII單倍型網絡關系圖(圖1)。在28種單倍型中,有13種單倍型(MHap1~MHap13)為南方種群所獨有且個體總和數量較大,有5種單倍型為北方種群獨有且個體總和數量較小,有10種單倍型(MHap14~MHap23)為南、北方種群所共有。在所有供試個體中出現頻率最高的3種單倍型依次為MHap14, MHap1和MHap3,其中在Hap14單倍型的組成個體中采自北方種群的個體數占比70.9%,南方種群的個體數占比29.1%。除上述3種單倍型外, MHap17, MHap15和MHap19也是出現頻率較高的單倍型種類,且這3種單倍型在南方和北方種群中均存在,所占數量比例不同。整個單倍型網絡結構可分成兩個部分:主干部分以MHap14為中心,有21種單倍型圍繞其呈輻射狀分布,共占全部個體數(含南方種群和北方種群)的96.1%;另一部分距離主干較遠(圖1中方框內所示),由7種單倍型組成且在種群中分布頻率很低,僅占到全部個體數的3.9%。兩部分內部的單倍型彼此間變異位點數較少,而兩部分單倍型之間的變異位點數較多。由COII單倍型序列構建的系統發生樹分支結構來看(圖2),在南方種群和北方種群中共檢測到的28種單倍型聚為兩大分支,第一大支共包含21種單倍型,所包括的單倍型涵蓋全部供試種群,其中南方種群和北方種群所共有的MHap15和MHap19兩種單倍型又與其他15種單倍型獨立分成一個分支;第二大支共包含7種單倍型,其中有4種單倍型為南方種群特有,分布于5個南方種群的少數個體中,另有1種單倍型(HQ909352)僅在北方的1個種群中出現(梁日霞等, 2011)。由單倍型網絡圖的拓撲結構與單倍型系統發生樹所展現的親緣關系可以看出,總體而言南方種群的單倍型多樣性更高且在種群中的分布更加分散,而北方種群的單倍型多樣性較低且在種群中的分布相對集中。

圖1 中國南方和北方雙斑長跗螢葉甲地理種群COII基因單倍型中介網絡圖

圖2 鄰接法構建的基于COII基因序列的中國南方和北方雙斑長跗螢葉甲地理種群單倍型系統發生樹(1 000次重復)
2.4 種群遺傳分化與基因流
14個雙斑長跗螢葉甲地理種群間總體Fst為0.2481,總體Nm為0.76,說明種群間遺傳分化程度普遍較高,種群間基因流弱。從各種群間這兩項參數來看,種群間遺傳分化指數Fst數值在0.0102~0.8239之間,其中有11對種群間無明顯的遺傳分化(Fst<0.05),占比12.09%;30對種群間存在中等水平的遺傳分化(0.05
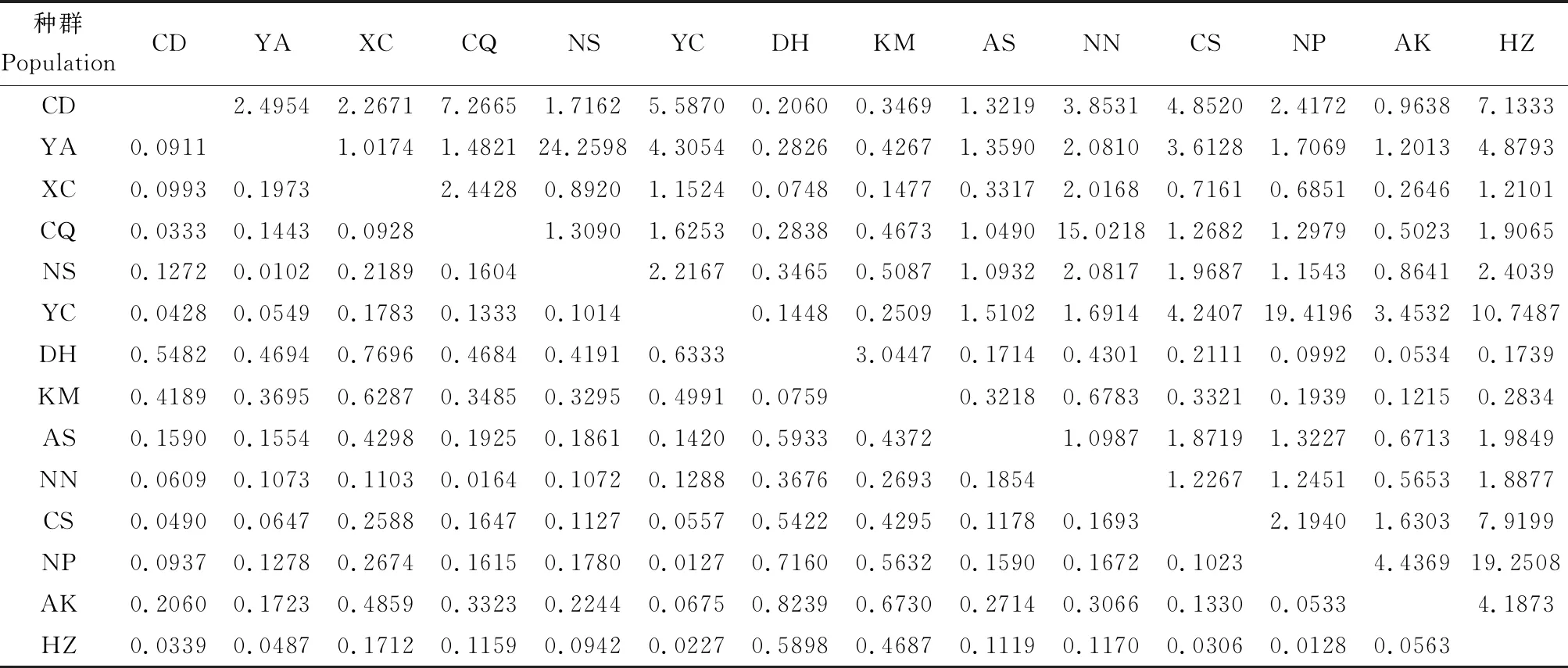
表3 基于COII基因序列的中國南方雙斑長跗螢葉甲地理種群間遺傳分化指數(Fst)(下三角)與基因流(Nm)(上三角)
AMOVA結果顯示COII基因遺傳變異種群間方差組分為0.4978,所占方差比率為26.25%;種群內方差組分為1.3986,所占方差比率為73.75%,二者均為極顯著水平(表4),表明雙斑長跗螢葉甲種群的遺傳分化主要來自種群內部個體間的變異,其次才是來自種群間的分化。
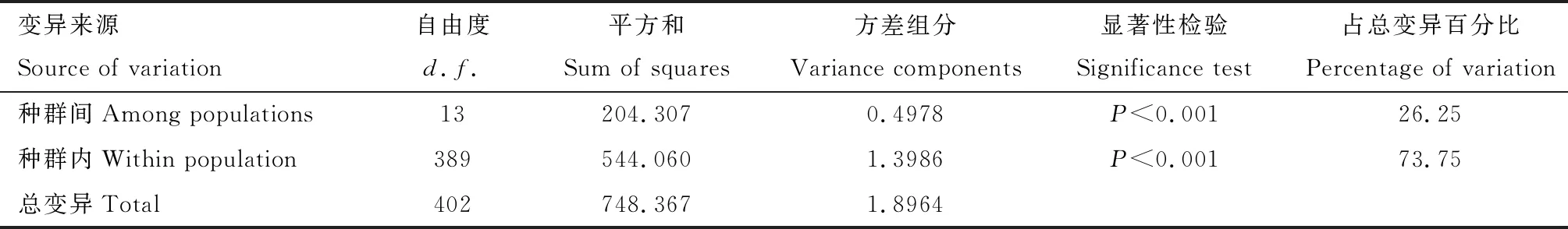
表4 基于COII基因序列的中國南方雙斑長跗螢葉甲地理種群變異分子方差分析
為了檢測雙斑長跗螢葉甲不同地理種群之間的遺傳分化程度是否與地理隔離相關,對兩兩種群間的遺傳距離與地理距離(取自然對數)之間的相關性進行Mantel檢驗,得到遺傳距離與地理距離之間的線性回歸方程為y=0.103x-0.435,相關系數R為0.2898(P=0.0640>0.05,1 000次隨機抽樣),表明雙斑長跗螢葉甲種群間的遺傳分化與地理隔離之間無顯著相關性(圖3)。
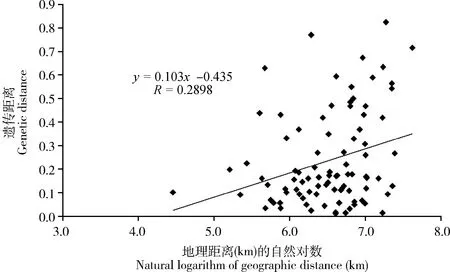
圖3 中國南方雙斑長跗螢葉甲地理種群間遺傳距離與地理距離對數相關性分析
2.5 Wolbachia感染率及感染類型
對14個地理種群403頭雙斑長跗螢葉甲個體進行Wolbachia共生菌感染檢測,結果發現共有396頭感染,各種群內感染率在92.59%~100%之間不等,平均感染率為97.60%,其中有7個種群感染率高達100%(表5)。可見Wolbachia在雙斑長跗螢葉甲內的感染相當普遍,在種群中均具有很高的感染率。
wsp基因序列分析發現供試雙斑長跗螢葉甲種群中共感染了6種Wolbachia株系(strain),根據通用命名法則將其命名為wMhie1~wMhie6,6個株系克隆獲得的wsp片段序列長度分別為624, 621, 651, 606, 594和636 bp(GenBank登錄號: JF747223-JF747228),wsp基因遺傳相似度在78%~93%之間,遺傳距離在0.025~0.150之間(表6)。在感染株系中,wMhie1株系在雙斑長跗螢葉甲種群中的感染率最高,占檢測個體的62.86%;其次是wMhie2,占檢測個體的26.43%;其他4種株系在種群中的感染率均較低,wMhie3僅在5個種群中檢測到(占檢測個體的5.00%),wMhie4和wMhie5僅在3個種群中檢測到,而wMhie6僅在2個種群中檢測到(表5)。
系統發育分析結果顯示,雙斑長跗螢葉甲中感染的6種Wolbachia株系(即wMhie1~wMhie6)全部屬于A大組,但在系統聚類上與A組中已知的代表性株系分開而單獨聚為一個分支,在這個分支內部又表現為wMhie4與wMhie6聚為一支,另外4個株系聚為另一支,且wMhie株系聚類分支結構的置信度分值均高于90,可見此聚類結果具有較高可信度(圖4)。基于以上結果,認為雙斑長跗螢葉甲中感染的6種株系均屬于不同于其他昆蟲物種中感染的Wolbachia類群。
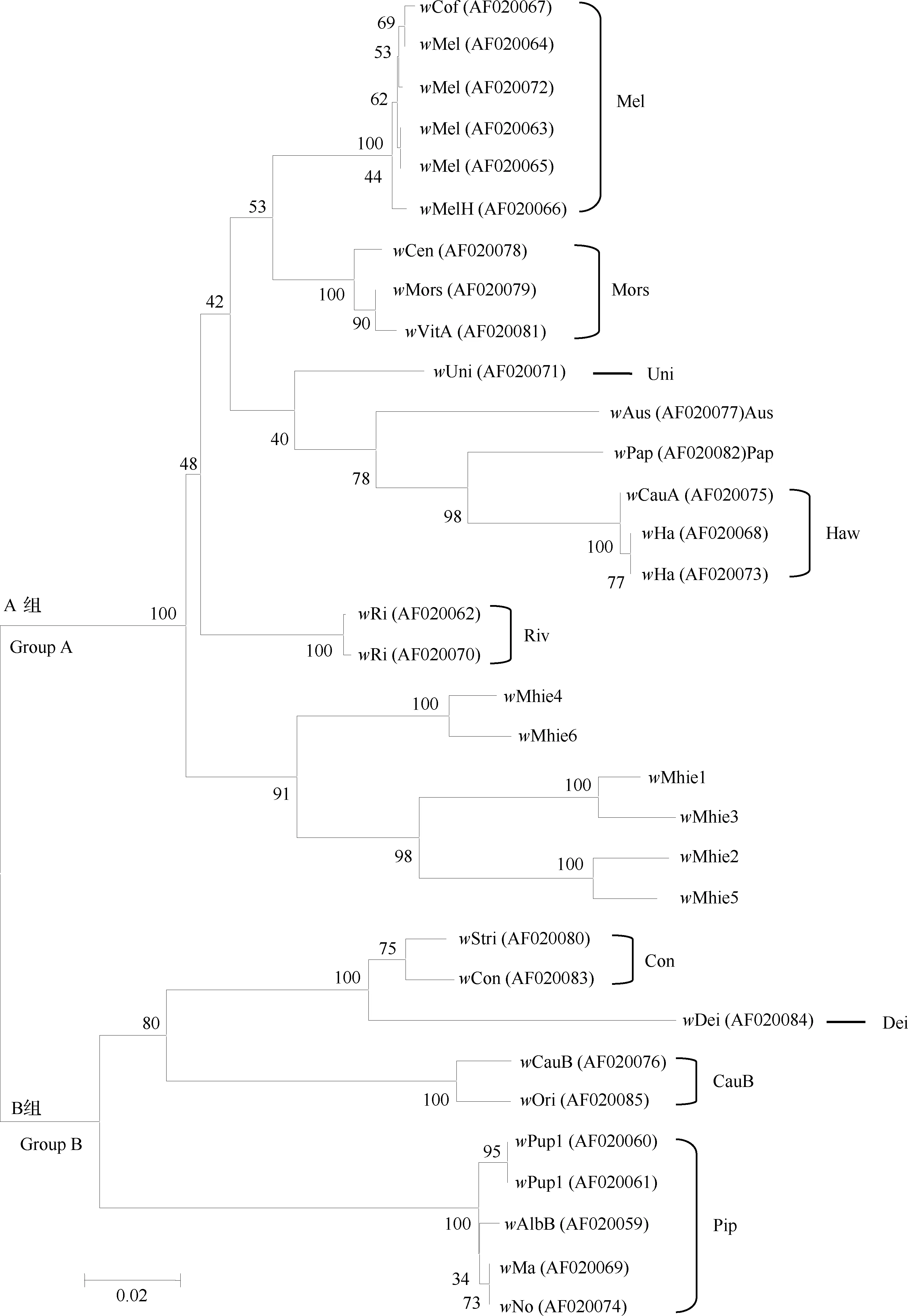
圖4 鄰接法構建的基于wsp序列的Wolbachia系統發育樹(1 000次重復)
3 討論
本研究以線粒體COII基因作為分子標記,對中國南方多省區分布的雙斑長跗螢葉甲地理種群的遺傳結構、遺傳多樣性及種群間遺傳分化程度進行了綜合分析。在先前報道的北方種群數據結果基礎上,又檢測到13種新的COII基因單倍型。與梁日霞等(2011)報道的北方種群相比,無論是總群體還是各種群間的平均值,南方種群在單倍型數、單倍型多樣性、核苷酸多樣度和核苷酸平均差異數等幾項參數指標上均明顯高于北方種群,說明雙斑長跗螢葉甲南方種群的遺傳多樣性普遍高于北方種群,即南方種群具有更豐富的遺傳多樣性。造成南北方種群遺傳多樣性存在差異的原因可能與環境溫度相關,已有研究發現雙斑長跗螢葉甲的發育歷期與溫度顯著相關,其卵、幼蟲和蛹的發育速率隨溫度升高而加快(李廣偉等, 2010; 張聰, 2012)。雙斑長跗螢葉甲在我國北方為一年發生1代,以卵在土壤中越冬(張聰等, 2013, 2014),而其在我國南方地區的發生代數迄今尚未有研究報道,推測該種昆蟲與鞘翅目其他甲蟲相似,即隨緯度降低年發生代數增加,因此較發生代數少、發育歷期長的北方種群,南方種群內部發生遺傳變異的幾率增加,種群內具有更豐富的遺傳多樣性。
遺傳分化指數Fst可反映出種群間的遺傳分化程度,通常Fst值越大說明群體間遺傳分化程度越高。普遍認為當Fst<0.05時群體間遺傳分化程度很低,當0.05
對比雙斑長跗螢葉甲北方種群的數據,本研究在南方種群中檢測到更加豐富的線粒體基因單倍型,從單倍型網絡關系圖(圖1)和系統發生樹的結構(圖2)來看,MHap14, MHap1和MHap3這3種單倍型在種群中出現頻率最高,且均位于單倍型的主干分支中,說明這3種單倍型作為該昆蟲種群中的優勢單倍型在群體中較穩定的遺傳下來。而有7種單倍型構成了一個與主干分支距離較遠的叢支,其中的單倍型與多數單倍型之間存在較多的變異位點。這些稀有單倍型的存在豐富了種群的遺傳多樣性,在一定程度上體現了昆蟲種群具備應對環境變化的進化潛力。在主干單倍型與稀有單倍型之間尚存在若干“過渡”單倍型在供試個體中未檢測到,可能這些缺失的單倍型(變異位點)由于存在個體稀少已逐漸被淘汰或由于采樣量的限制未被檢測到。
本研究供試雙斑長跗螢葉甲種群中性檢驗結果大多為負值且不顯著(表2),因此不支持種群數量消減或增長、經歷瓶頸效應或發生大規模遷移事件的假設,在進化上遵循中性模型,表明種群在較近的歷史時期內沒有經歷明顯的群體擴張事件,種群大小保持相對穩定(Korneliussenetal., 2013),此推論與北方種群的檢驗結果(梁日霞等, 2011)一致。雙斑長跗螢葉甲為多食性昆蟲,寄主植物種類豐富,因此能應對環境的變化而保持種群的穩定,另外該種昆蟲飛行能力弱,在一定程度上制約了種群的遷移和擴張,這也是維持種群相對穩定的另一個原因。
有研究提出Wolbachia在宿主中的感染率常遵循“或多或少(most or few)模式”,即一個物種內的Wolbachia感染率通常會很高(>90%)或很低(<10%)(Hilgenboeckeretal., 2008)。對葉甲科昆蟲Wolbachia感染已有一些研究,Ali等(2018)通過檢測發現椰心葉甲Brontispalongissima5個東南亞種群中Wolbachia感染率高達100%,且感染類群十分豐富。本研究發現,中國南方雙斑長跗螢葉甲所有供試種群中的Wolbachia感染率均高于90%,其中有一半種群100%感染,整體平均感染率高達97.60%(表5),證明Wolbachia在雙斑長跗螢葉甲宿主中的感染率符合上述規律。通過wsp基因序列共檢測到6種Wolbachia株系,表明在雙斑長跗螢葉甲宿主中感染著豐富的Wolbachia類群。系統進化分析顯示雙斑長跗螢葉甲中感染的6種株系均與目前已知感染昆蟲的代表性株系在系統進化上關系較遠(圖4),尚無法在系統發生上對其準確劃分類群,且這6種Wolbachia株系是否具有對雙斑長跗螢葉甲宿主具有生殖調控作用仍然未知。
近年來越來越多的研究發現,Wolbachia對其昆蟲宿主的調控作用可能在宿主種群發生遺傳分化過程中起到了重要的作用,其中一個潛在機制是Wolbachia誘導的胞質不親和能夠有效阻礙感染不同株系的宿主種群之間進行基因交流,從而加速了宿主種群間的遺傳分化,甚至可能促進新物種的形成(J?ckeletal., 2013; Zhangetal., 2013; Lisetal., 2015)。本研究發現雙斑長跗螢葉甲種群間普遍存在遺傳分化且基因交流不頻繁,這種現象是否與Wolbachia的生殖調控作用相關仍是未來研究中需要解答的一個問題。在今后的研究中,還需要通過MLST(多位點序列分型)系統對雙斑長跗螢葉甲中感染的Wolbachia類群進行更加系統的鑒定,并將Wolbachia感染類群與宿主線粒體單倍型進行結合分析,進一步探究Wolbachia感染是否在雙斑長跗螢葉甲種群間產生遺傳分化過程中發揮了作用。
