云南切梢小蠹幼蟲表皮碳氫化合物與齡數的相關性*
2021-07-13 07:54:16張夢蝶錢路兵澤桑梓李宗波
林業科學 2021年5期
張夢蝶 錢路兵 澤桑梓 楊 斌 李宗波
(1.西南林業大學生物多樣性保護學院 云南省森林災害預警與控制重點實驗室 昆明 650224; 2.云南省林業和草原有害生物防治檢疫局 昆明 650224)
昆蟲表皮碳氫化合物(Cuticular hydrocarbons, CHCs)是指廣泛存在于昆蟲上表皮中一類由長鏈烷烴和烯烴所組成的混合物,其性質穩定,加之成分和含量在種間種內的差異性,即使是復合種、近緣種、隱形種、同種個體的不同發育階段或社會品級等也有明顯差異,對于昆蟲化學分類、物種演化、信息通訊與調控、胚后發育、農林害蟲治理及其益蟲利用等研究均具有重要的啟示作用(Pageetal., 1997; Blomquistetal., 2010; Yoonetal., 2012; Derstineetal., 2018; Bersonetal., 2019; 李群臣等, 2019)。云南切梢小蠹(Tomicusyunnanensis)隸屬于鞘翅目象蟲總科(Curculionoidea)小蠹科(Scolytinae)切梢小蠹屬(Tomicus),又稱為縱坑切梢小蠹或云南松縱坑切梢小蠹,是一種為害云南松的蛀干害蟲,自20世紀80年代暴發危害以來,累積致死的活立木蓄積量超過20萬hm2,嚴重影響了云南松林的可持續發展和生態安全(李麗莎 等, 1997; Kirkendalletal., 2008; 沈紹偉等, 2018)。當前,雖然已有一套關于云南切梢小蠹的綜合治理措施,例如噴灑殺蟲劑和生物制劑、釋放天敵昆蟲、信息素和寄主氣味或餌木誘殺、非寄主氣味干擾、營造混交林等(李浩然等, 2000; 王海林等, 2002; 李雙成等, 2003; 葉輝等, 2006; 岳鋒等, 2011; 王大偉等, 2015; Wuetal., 2019),但仍無法控制該蟲擴散蔓延的趨勢,這說明在云南切梢小蠹治理體系中存在著缺失環節,特別是關于云南切梢小蠹的發育生物學特征。但對于蛀干、潛葉、土壤中等營隱蔽式生活和危害的昆蟲,其發育生物學難以直接觀察,特別是幼蟲的齡數不易確定; 而通過人工室內飼養導致幼蟲生活環境的改變,常常引起齡數出現較大變化,從而無法精確了解寄主幼蟲發育齡數及發育進度。由于這種基礎研究的缺失,會極大地降低害蟲科學預測預報的準確性,隨之降低有害生物的綜合治理效能(陳輝, 2002; Castexetal., 2018),如遷飛高峰措置(Peak shift displacement)使得噴灑的真菌制劑殺蟲效果不佳,或者釋放的天敵昆蟲無法及時找到合適寄主(王海林等, 2002; 葉輝等, 2006)。同時,隨著全球化進程的加快,國際交往日益頻繁,并考慮到全球氣候變化的大背景(Castexetal., 2018; Liuetal., 2019),該蟲的擴散及危害潛能會被放大,產生的生態破壞性將難以估量。本研究旨在通過林間大量抽樣調查,運用形態數據測定自然條件下云南切梢小蠹幼蟲的齡數,結合不同齡級幼蟲的表皮碳氫化合物進行定性定量分析,為深入了解云南切梢小蠹生物學特征及其科學治理該蟲危害提供依據。
1 材料與方法
1.1 幼蟲的獲取與判定
云南切梢小蠹幼蟲采集于曲靖市沾益區九龍山林場(103°7′15″ E,25°0′35″ N),為該區域內林木害蟲的優勢種群,主要在樹干中上部蛀干繁殖(沈紹偉等, 2018)。于2017年11月至2019年5月,分批次帶回新蛀入或已在樹干內發育的云南切梢小蠹幼蟲。為了更準確地區分齡數,將位于坑道兩側的卵(60粒)置于培養皿中的濕潤濾紙上,室溫條件下保濕培養,待其孵化后作為1齡幼蟲(圖1A),而老齡幼蟲以蛹室形成且有木屑封堵為判斷依據(圖1B)。將每次采集的云南切梢小蠹幼蟲分別放入75%的乙醇溶液中保存,共收集到各齡幼蟲1 426頭。參考象蟲總科幼蟲齡數的劃分標準(Mizelletal., 1979; Castaeda-Vildzolaetal., 2016),用蔡氏體式鏡(V20,German)對各個幼蟲進行拍照,用Image J(Version 1.62,https:∥imagej.nih.gov)測量其頭殼寬和體長,并用掃描電鏡(Zeiss sigma 300, German)觀察不同齡數幼蟲的微觀形態,測其上顎長與寬。

圖1 云南切梢小蠹初孵幼蟲和末齡幼蟲的判定Fig. 1 Determination of larval instars in Tomicus yunnanensis using the freshly hatched larvae from eggs and last larvae blocked in the pupal cellsA1: 坑道兩側的卵 Eggs on the both sides of gallery ; A2: 初孵幼蟲 The freshly hatched larvae from eggs; B1: 封堵在蛹室(虛線)內的老齡幼蟲 The last instar larvae are blocked by the wood chips (the arrow) in the pupal cell (the dashed line); B2: 末齡幼蟲 Morphology of the last instar larvae.
1.2 幼蟲表皮碳氫化合物的提取與分析
采用浸提法對云南切梢小蠹不同齡數幼蟲的表皮碳氫化合物進行提取(Pageetal., 1997; Blomquistetal., 2010)。首先將從木段中解剖出來的幼蟲立即置入-20 ℃冰箱內殺死,超聲波清洗后(冰浴),依據頭殼寬確定齡數,每齡重復15次,每次測定60只,用400 μL正己烷洗滌2次,第1次10 min(靜置5 min,震蕩1 min,再靜置3.5 min,震蕩30 s),第2次靜置1 min,然后將混合液放置在2 mL棕色試劑瓶內,-20 ℃保存待用。在GC-MS(Agilent 7890A-5975C,USA)分析以前,先將樣品用氮氣吹干,滴入少量正己烷,接著加入內標化合物正十四烷(n-C14)和正十六烷(n-C16)各2 μL(2 ng·μL-1),定容至50 μL。接著取1 μL定溶液從進樣口注入,質譜柱型號為極性HP-5 MS(30 m × 0.25 mm × 0.25 μm)。升溫程序: 起始溫度為40 ℃,以10 ℃·min-1升溫至150 ℃,以5 ℃·min-1升溫至300 ℃,保持10 min。高純度氮氣作為運載體,流速為1 mL·min-1,進樣口溫度250 ℃,柱前壓100 kPa,進樣采用不分流模式。MS條件: EI源,70 eV,傳輸線溫度250 ℃,離子源溫度230 ℃,質量掃描范圍全掃描。采用NIST 08譜庫檢索法、峰面積標準化法(Aitchison, 1986)、科瓦特指數法(Marquesetal., 2000)及其與C8—C40烷烴混合標準品(500 μg·mL-1,AccuStandard,USA)比對進行鑒定(計算公式如下),化合物含量根據內標的量計算。
Zip= ln[Aip/g(Ap)]。
式中:Zip為轉換后待鑒定化合物的峰面積;Aip為待鑒定化合物的峰面積;g(Ap)為各組分色譜峰面積的幾何均值。
KI=100n+100 ×[tR(x)-tR(n)]/[tR(n+1)-tR(n)]。
式中:KI為科瓦特指數;n為待鑒定化合物前面正構烷烴的碳原子數;x為待測化合物碳原子數;tR(x)為保留時間;tR(n)和tR(n+1)分別為待鑒定化合物色譜峰前一個和后一個正構烷烴的保留時間。
1.3 數據處理與分析
所有的數據統計分析均在開源軟件R(Version 3.5.1,https:∥cran.r-project.org/)中進行,制圖用GraphPad Prism(Version 8.0.2)完成。對測定的頭殼寬度,首先進行頻數分布分析,間隔最小設置為0.02 mm,接著再用體長、上顎長和寬等進行驗證,以此確定云南切梢小蠹幼蟲齡數。齡數劃分后,進一步根據測定指標的均值、標準誤差、變幅、變異系數、Brooks指數、Crosby指數等確定最佳齡數,若變異系數小于20%、Crosby指數小于10%,則表明分齡指標合理(Castaeda-Vildzolaetal., 2016),接著用回歸分析擬合齡數與體長、頭殼寬間的關系,用Tukey氏多重比較法對比各齡幼蟲指標的差異性。對于表皮碳氫化合物,利用FactoMineR軟件包進行主成分分析(Lêetal., 2008),選用極大方差法旋轉因子,并基于相關系數矩陣計算主成分得分系數,提取特征值大于1的成分,最后再用RandomForest軟件包進行化合物貢獻量分析(Mccormicketal., 2014),提取基尼指數(MeanDecreaseGini)大于2的成分; 用Kruskal-Wallis test進行各齡數幼蟲化合物總量和關鍵化合物量的多重比較。
2 結果與分析
2.1 云南切梢小蠹幼蟲的齡數
幼蟲頭殼寬度頻次分布具有3個界線明顯的峰區,而每個峰區中又有明顯的峰值,分別位于0.50、0.825和1.04 mm處(圖2),因幼蟲發育過程中需解除外骨骼限制而產生蛻皮,每次蛻皮幼蟲即會突長一次,進入下一齡,所以從頻次圖上可以推測云南切梢小蠹幼蟲共有3齡。進一步來看,幼蟲頭殼寬度與齡數間的變異幅度是比較小的(7%~14.6%),且Crosby指數小于0.1,并經統計檢驗和回歸分析均達到了極顯著水平(P<0.000 1; 表1),表明將云南切梢小蠹幼蟲齡數劃分為3齡是合理準確的。掃描電鏡觀察顯示云南切梢小蠹幼蟲共有12體節,其表面微觀形態,包括前額心形盾及其表面剛毛數量和排列形式、額須上的味覺感器、上顎及其齒數、內唇等均沒有不同,僅長度與寬度有所增加(圖3; 表1)。當用體長、上顎長和寬進行齡數擬合時,盡管這些指標均可將云南切梢小蠹幼蟲劃分為3齡,統計檢驗和回歸分析也顯示有顯著差異(體長:F=3 383.6,P<0.000 1; 上顎長:F=740.82,P<0.000 1; 上顎寬:F=91.395,P<0.000 1; df=1),但幼蟲體長的變異系數超過20%,2齡和3齡幼蟲的上顎寬度間差異不顯著(表1)。

圖2 云南切梢小蠹幼蟲頭殼寬度頻次分布Fig. 2 Frequency distribution of the head-capsule widths of the larval T. yunnanensis
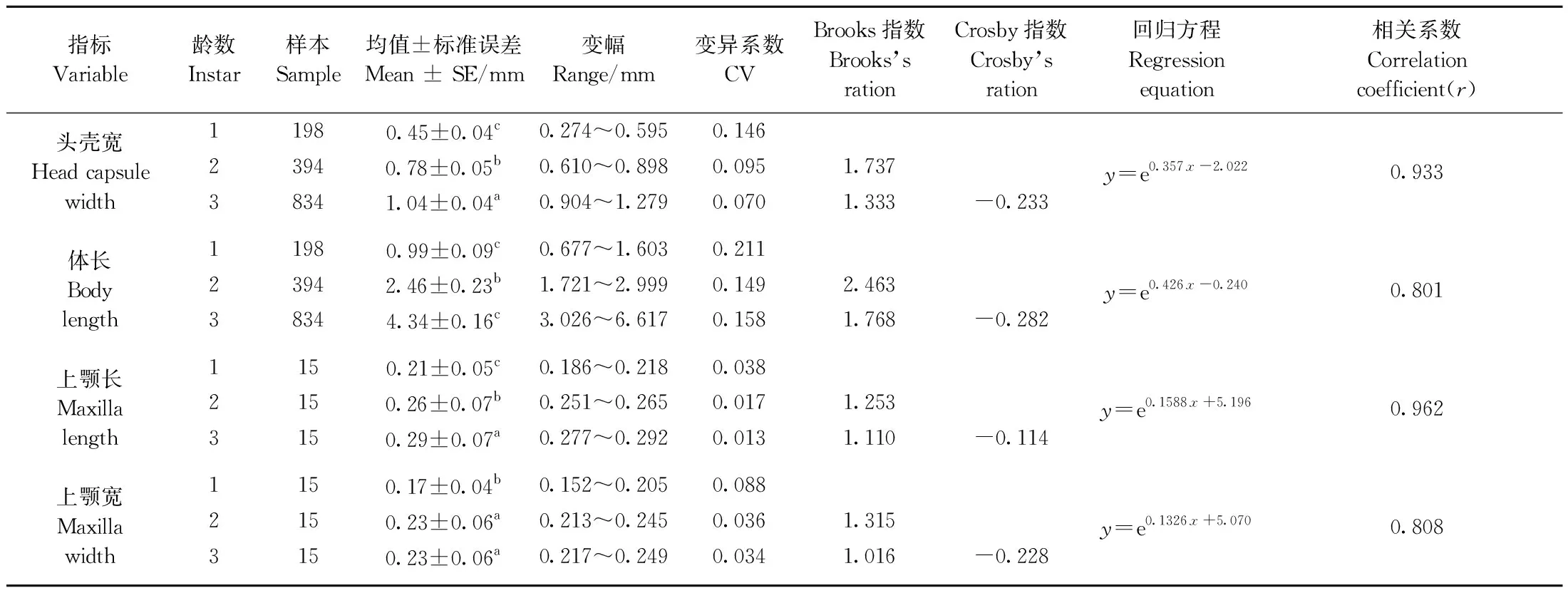
表1 云南切梢小蠹幼蟲各項測量指標的分齡與統計①Tab.1 Measurement and statistical analysis of the four various indexes for the larval instars of T. yunnanensis
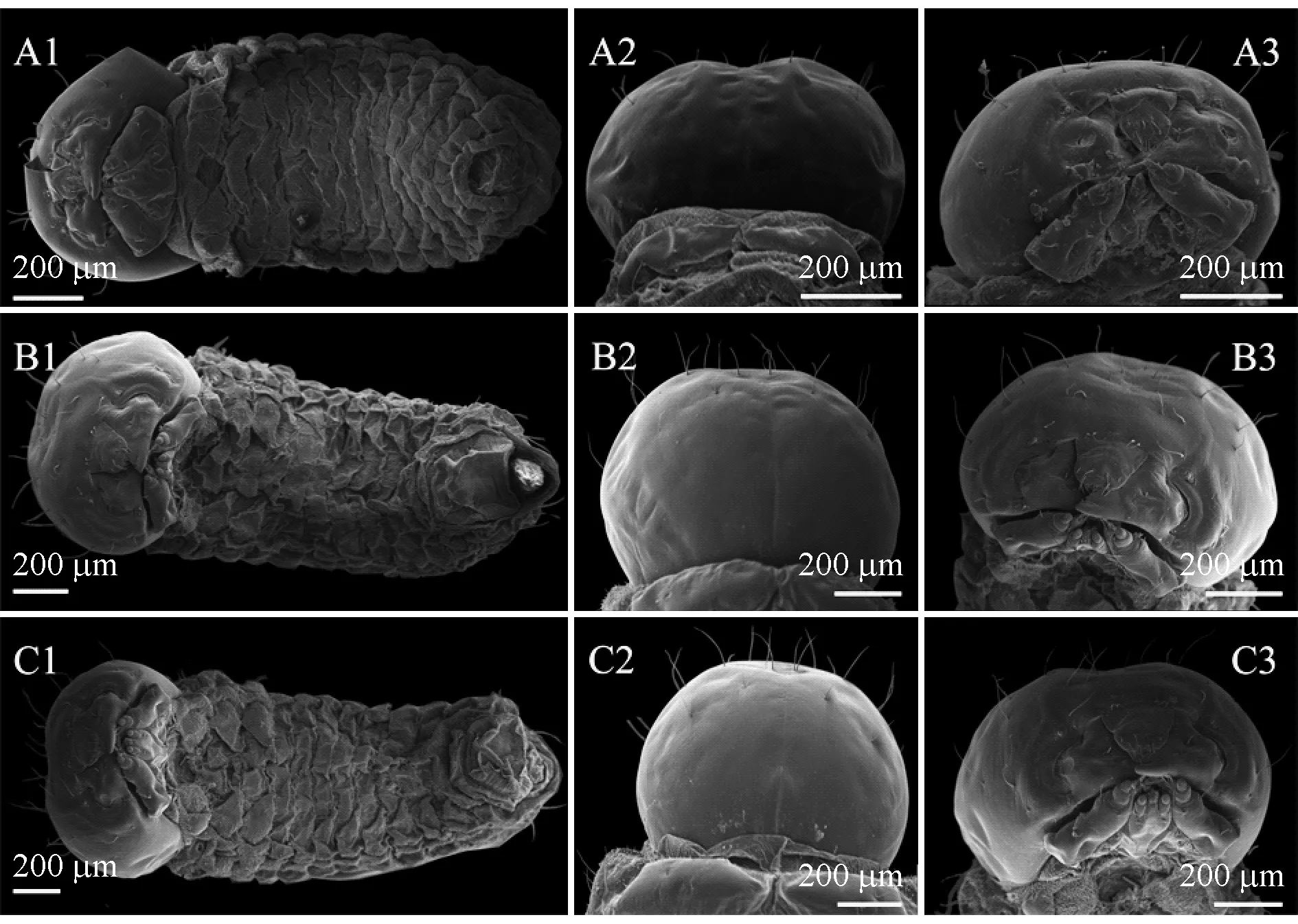
圖3 云南切梢小蠹不同齡數幼蟲的微觀形態Fig. 3 Micrograph of larval instars of T. yunnanensisA—C: 1~3齡幼蟲 The instar numbers of the larvae from the first instar to the third instar; 字母后的數字1~3分別示整體、頭殼背面和腹面 The numerals behind the capital letter indicate the ventral view of the larval body, the dorsal and ventral view of larval head capsule, respectively.
2.2 幼蟲表皮碳氫化合物種類與組成
共從云南切梢小蠹幼蟲中鑒定出19種表皮碳氫化合物,主要為正鏈烷烴、支鏈烷烴和烯烴3類烴類物質,含量分別為每蟲4.41 ng± 0.67 ng、0.26 ng ±0.04 ng和1.22 ng± 0.18 ng,各烴類物質在不同齡級幼蟲中分布相差較大(表2)。其中,正鏈烷烴以正二十五烷和正二十七烷為代表,占比分別為55.02% ± 8.20%和21.37% ± 3.19%; 支鏈烷烴中以13-甲基-二十九烷含量最高,占整個支鏈烷烴的94.04%±14.02%; 烯烴類以1, 21-二十二烷二烯為主,占比為63.03%±0.39%。
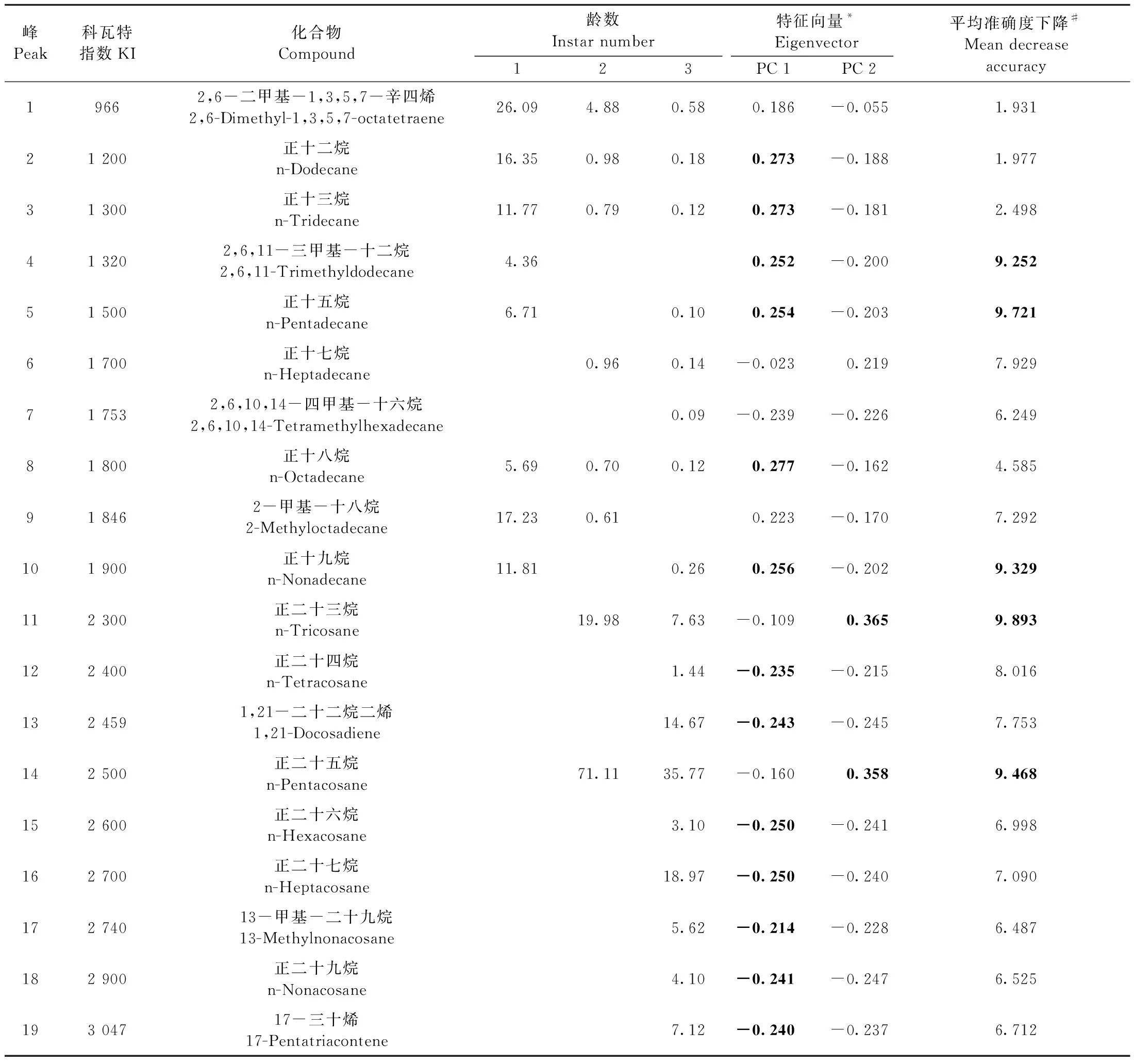
表2 云南切梢小蠹不同齡數幼蟲的表皮碳氫化合物種類、含量及其主成分分析與隨機森林判定①Tab.2 Mean relative content of cuticular hydrocarbon and their results of principal component scores and random forest analysis on the larval instars of T. yunnanensis
2.3 不同齡數幼蟲表皮碳氫化合物對比
云南切梢小蠹初孵幼蟲(1齡幼蟲)的表皮碳氫化合物有8個,碳數介于10~19之間,C10—C14和C15—C19各占近50%; 2齡幼蟲有8個化合物,碳數為10~29,其中C20—C24和C25—C29為主要成分,分別占19.98%和71.11%; 而老熟幼蟲(3齡幼蟲)則有17個,以C25—C29和C30—C35為主,分別占61.94%和12.74%(表2)。隨著齡級增加,幼蟲表皮碳氫化合物的含量分別為每蟲2.32 ng±0.59 ng、44.40 ng±11.49 ng和280.75 ng±72.48 ng,齡級間有著顯著差異(χ2=39.13,P<0.000 1)(圖4)。主成分分析顯示第一主成分和第二主成分聯合貢獻率為81.7%,其中正十二烷(7.46%)、正十三烷(7.44%)、2,6,11-三甲基-十二烷(3.48%)、正十八烷(7.67%)等12種化合物超過第一主成分貢獻量,正二十三烷(13.32%)和正二十五烷(12.78)是第二主成分的重要貢獻者,不同齡數幼蟲的化合物分離程度高,齡數與齡數間無重疊區域(表2、 圖5)。在幼蟲表皮碳氫化合物構成的重要性方面,隨機森林分析顯示2,6,11-三甲基-十二烷、正十五烷、正十九烷、正二十三烷和正二十五烷5種化合物具有較大的重要性(表2)。其中,2,6,11-三甲基-十二烷僅存在于1齡幼蟲,而正十五烷在3齡幼蟲中相對含量僅為0.1%,為一種微量化合物。在不同齡級幼蟲上,這5種化合物的含量上有著顯著差異(P<0.000 1),2,6,11-三甲基-十二烷為1齡幼蟲的標識性物質,而正二十三烷和正二十五烷可作為2齡和3齡幼蟲的識別物(表2、圖6)。

圖5 云南切梢小蠹不同齡數幼蟲表皮碳氫化合物的主成分得分Fig. 5 Scores plot of principal components extracted from cuticular hydrocarbons of the larval instar of T. yunnanensis
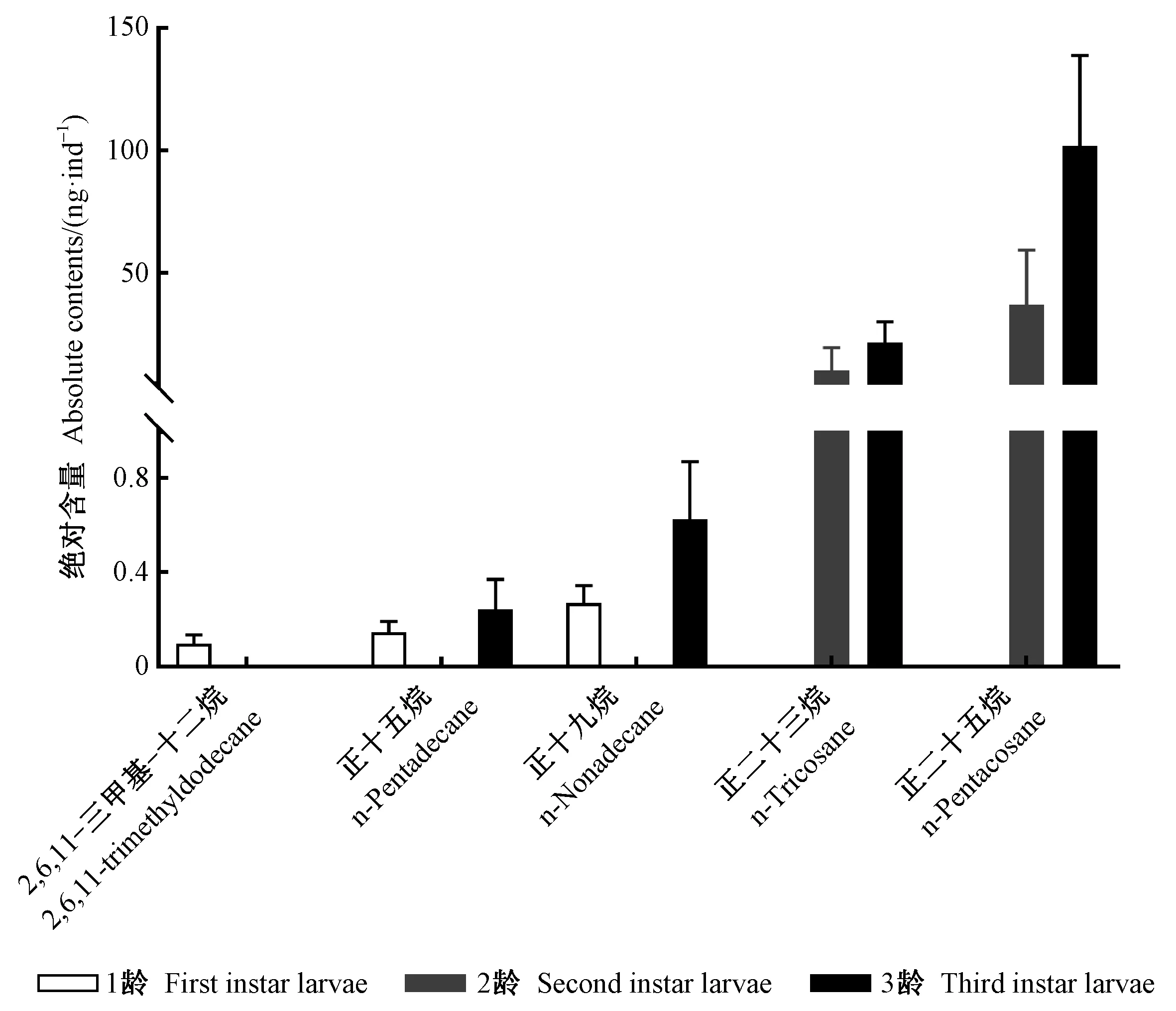
圖6 云南切梢小蠹不同齡數幼蟲表皮中5種重要碳氫化合物的含量Fig. 6 Mean contents of five important compounds of cuticular hydrocarbons on the larval instar of T. yunnanensis
3 討論
昆蟲表皮碳氫化合物是昆蟲化學分類中采用時間最久的生物學指標(Bioindicators)(Jackson, 1970; Blomquistetal., 2010; 李群臣等, 2019),如早在20世紀70年代,Jackson (1970)就發現碳數為23~26的正二十三烷、順-9-二十三烯、3-甲基-二十三烷、11-甲基-二十三烷、13-甲基-二十五烷等5種長鏈烷烴可輕松將3種大蠊(Periplaneta)區分開。隨著化學分析技術及其多元統計分析方法的大量應用,發現表皮碳氫化合物除可區分蜚蠊目外,還可用來區分鞘翅目(Pageetal., 1997)、鱗翅目(Hay-Roeetal., 2007)、雙翅目(Barbosaetal., 2017)、半翅目(Yoonetal., 2012)、等翅目(Blomquistetal., 2010)、同翅目(Raboudietal., 2005)、膜翅目(Soaresetal., 2017)、直翅目(Brozaetal., 2000)等眾多昆蟲,并且一些表皮碳氫化合物反映了其胚后發育狀態(Zhuetal., 2006; Blomquistetal., 2010; Kuoetal., 2012; Yoonetal., 2012)。例如,漸變態的點蜂緣蝽(Riptortuspedestris)卵的標識物是3-甲基-二十七烷,若蟲是正二十九烷,成蟲是正二十七烷、正三十一烷、13,17-/15,19-二甲基-三十三烷(Yoonetal., 2012); 而全變態的紅顏金蠅(Chrysomyarufifacies)幼蟲期主要是正烷烴類,隨齡級增加短鏈碳氫化合物含量呈指數級減少,代表性物質是正二十五烷和正二十九烷(Zhuetal., 2006)。不僅如此,形態極為相似的近似種鱗翅目夜蛾科的棉鈴蟲與煙青蟲2種幼蟲表皮碳氫化合物組分也出現區別,并且材料為5~6齡幼蟲碳數分布24~32與云南切梢小蠹幼蟲3齡幼蟲主要碳數分布25~35極為相似(高明媛等, 1999)。從云南切梢小蠹幼蟲表皮碳氫化合物的結果來看,不同齡級幼蟲的化合物組成差別較大,特別是對分類有重要意義的2,6,11-三甲基-十二烷、正十五烷、正十九烷、正二十三烷和正二十五烷這5種化合物,良好的分離效果可能反映了其在齡級中的動態變化,也可能與總含量的指數級增加有關(y=e0.086 5x-0.629,R=0.911)(圖4)。盡管一些化合物含量極低,但其在化合物整體組成中不可或缺,顯示了一些新的成分分析方法,如隨機森林、非度量多維尺度分析(non-metric multidimensional scaling,NMDS)、典型辨別分析(canonical discriminant analysis,CDA)等,對于探索多元數據背后所隱含生物學含義具有重要的作用(Mccormicketal., 2014)。另一方面,云南切梢小蠹幼蟲在樹干內發育,且每一個幼蟲均有各自獨立的坑道進行取食,考慮到樹皮提供的保護及相對高含水量的環境,防止體內水分流失應不是營隱蔽生活昆蟲所面臨的問題,更多應該與其生理活動和代謝水平有關(Blomquistetal., 2010),正如1齡幼蟲的烷烴類化合物僅有8個,且碳數均小于20,而隨著齡級的增加,老熟幼蟲有17種,而碳數也達到最大的35個,其發育進度中化合物種類與碳數增加的結果與棲境相似昆蟲在發育進度的變化相符(Zhuetal., 2006; Kuoetal., 2012; Yoonetal., 2012),但與裸露生活的點蜂緣蝽、葉甲、胡蜂、齒小蠹成蟲等差別較大(Pageetal., 1997; Yoonetal., 2012; Zhangetal., 2014; Soaresetal., 2017)。另外,昆蟲表皮碳氫化合物鏈長每增加一個仲碳原子(-CH2-),其熔點會提升2 ℃,進而會增加幼蟲體壁的堅硬程度,對溫度的容忍能力亦會提升(Menzeletal., 2017),這也很好地解釋了隨著昆蟲齡數的增加,包括云南切梢小蠹在內的眾多昆蟲的表皮碳氫化合物鏈長為何增加的現象。
用幼蟲表皮碳氫化合物可輕松地將云南切梢小蠹3個齡級幼蟲區分開,比用頭殼寬度、體長、上顎長與寬的分離效果更好(表1、圖5)。昆蟲的生長周期表現出明顯的周期性,特別是鱗翅目(Godinetal., 2002)、鞘翅目(Mizelletal., 1979; Castaeda-Vildzolaetal., 2016)、雙翅目(Richardietal., 2013)、膜翅目(Vilhelmsen, 2011)等幼蟲頭殼骨化程度高且相對穩定的昆蟲,身體各部的長寬會按指數級增長,局部的生長狀況可以指示整個蟲體的生長狀況。根據云南切梢小蠹幼蟲的4個測定指標,雖然均可將其劃分為3齡,但因各齡幼蟲體長和上顎寬重疊度較大,加之顎的長與寬測定相對困難,以它們作為齡數劃分依據,存在諸多的限制條件,因此,出頭殼寬度仍不失是一個良好的形態指標。但是,林間大量抽樣過程中仍會出現明顯錯誤,特別是頭殼寬為0.60~0.675 mm和0.9~0.94 mm這些介于齡數之間的蟲體樣本,而用幼蟲表皮碳氫化合物能夠明顯克服這些缺點,特別是用基尼指數大于2的5種化合物或者對齡期有重要標識的2,6,11-三甲基-十二烷、正二十三烷和正二十五烷。未來,將幼蟲表皮碳氫化合物的研究結果與便攜式氣質聯用儀、電子鼻、表面增強拉曼光譜等快速分析鑒定方法相結合(Castexetal., 2018; Turlingsetal., 2018),會實現云南切梢小蠹發育進度的實時監測,為提升預測預報精度和綜合治理該蟲危害奠定基礎。
4 結論
云南切梢小蠹幼蟲表皮碳氫化合物由正鏈烷烴、支鏈烷烴和烯烴3類19種烴類物質組成,從1齡幼蟲至3齡幼蟲,化合物數量從8個增加到17個,碳數從10增加到35,含量從每蟲0.139 ng±0.007 ng增至16.845 ng±1.366 ng,呈指數級遞增。幼蟲表皮碳氫化合物可明顯將不同齡數的幼蟲區分開,齡數與齡數間無重疊,劃分標準顯著優于頭殼寬、體長、上顎長與寬等形態指標法。2,6,11-三甲基-十二烷、正十五烷、正十九烷、正二十三烷、正二十五烷具有較大的重要性,含量上亦有顯著差異,其中2,6,11-三甲基-十二烷可用于1齡幼蟲的標識性物質,正二十三烷和正二十五烷可識別2齡和3齡幼蟲,這將為研究云南切梢小蠹幼蟲發育進度及監測預報提供科學依據。
