牛奶甘油三酯脂肪酸酯化位置結構特點及其在乳腺中合成代謝的研究進展
2021-07-12 03:04:20哈斯額爾敦薩茹麗敖長金
動物營養學報 2021年6期
哈斯額爾敦 白 晨 薩茹麗 敖長金
(內蒙古農業大學動物科學學院,內蒙古自治區高校動物營養與飼料科學重點實驗室,呼和浩特010018)
乳脂肪是牛奶重要營養組成,是膳食脂肪來源。乳脂肪中含有400種以上脂肪酸,其中不乏功能性脂肪酸,如多不飽和脂肪酸、短鏈脂肪酸等,同時含有具有潛在健康危害的中長鏈飽和脂肪酸[1]。因此,調控乳脂肪組成與產量對改善乳及乳制品營養品質,并以此平衡飼養效率具有重要意義。乳脂肪是乳成分中最易變化的組分,科學家在此特性上獲得了充分的試驗證據,主要包括提高乳脂率和乳脂產量或與其相反降低乳脂率(乳脂降低綜合征),以及針對性提高功能性脂肪酸含量或降低有潛在危害脂肪酸的比例等方面[2]。
甘油三酯是天然脂肪的主要存在形式,占乳脂肪95%以上,乳脂肪內甘油三酯的結構特點對其膳食營養價值和產品加工工藝有重要意義[2]。近年在人脂肪營養的研究中發現,除脂肪酸組成外,脂肪酸在甘油三酯中的酯化位置對于脂肪和脂肪酸在體內的消化、吸收和代謝途徑及代謝效率同樣具有決定性作用[3]。因此,從人膳食健康角度考慮,了解牛奶脂肪中甘油三酯結構特點及其合成代謝的影響因素是必要的。乳腺合成乳脂肪過程中甘油三酯酯化反應是必經途徑,脂肪酸在此過程中被分配于甘油骨架上。該反應的發生、發生的程度和產物的結構決定乳脂肪的組成、產量及其理化特性。以往的研究雖然認為哺乳動物乳腺內合成甘油三酯時酯化位置具有一定的特異性和保守性,但就奶牛而言,該“特異性和保守性”的范圍隨乳脂肪組成和產量的變化而發生的變化更為復雜。因此,本文綜述了甘油三酯脂肪酸酯化位置結構檢測方法與營養意義、牛奶甘油三酯脂肪酸酯化位置結構特點、乳腺合成代謝甘油三酯及其對牛奶甘油三酯結構特點的影響方面的研究進展,并討論了未來研究趨勢,以期為相關領域的研究提供參考。
1 甘油三酯脂肪酸酯化位置結構檢測方法與營養意義
1.1 檢測方法
甘油三酯是天然油脂的主要存在形式,是1分子甘油的3個羥基位分別與3分子脂肪酸羧基位逐一發生酯化反應的產物。酯化的空間結構位置排序(stereo-specifically numbered,sn)分別以sn-1、sn-2和sn-3表示(圖1)。
由于檢測繁瑣以及定量不準確等因素,天然油脂(包括乳脂肪)甘油三酯脂肪酸酯化位置結構特點一直沒有得到廣泛的分析研究。隨分析化學檢測技術及合成甘油三酯異構體工業技術的發展,甘油三酯脂肪酸酯化位置分布的檢測技術也不斷得以改進,操作方法趨于簡便。目前檢測天然油脂中甘油三酯脂肪酸酯化位置分布的方法主要有脂肪酶法、磷脂酶A2法、衍生化試劑法、高效液相色譜(HPLC)法、核磁共振定量檢測法等。其中反向高效液相色譜(RP-HPLC)法、專一性脂肪酶法以及二者結合的方法可有效地定性和定量檢測脂肪酸酯化位置分布[4-5]。近年的研究報道,超高效液相色譜聯用三倍四級質譜儀(UPLC-QQQ)或超臨界流體色譜聯用飛行時間質譜(SFC-MC)可提高甘油三酯脂肪酸酯化位置結構的檢測時效[6-7]。另外,磷核磁共振可定量檢測部分酯化甘油酯,而碳核磁共振可檢測甘油酯脂肪酸酯化位置結構[8]。
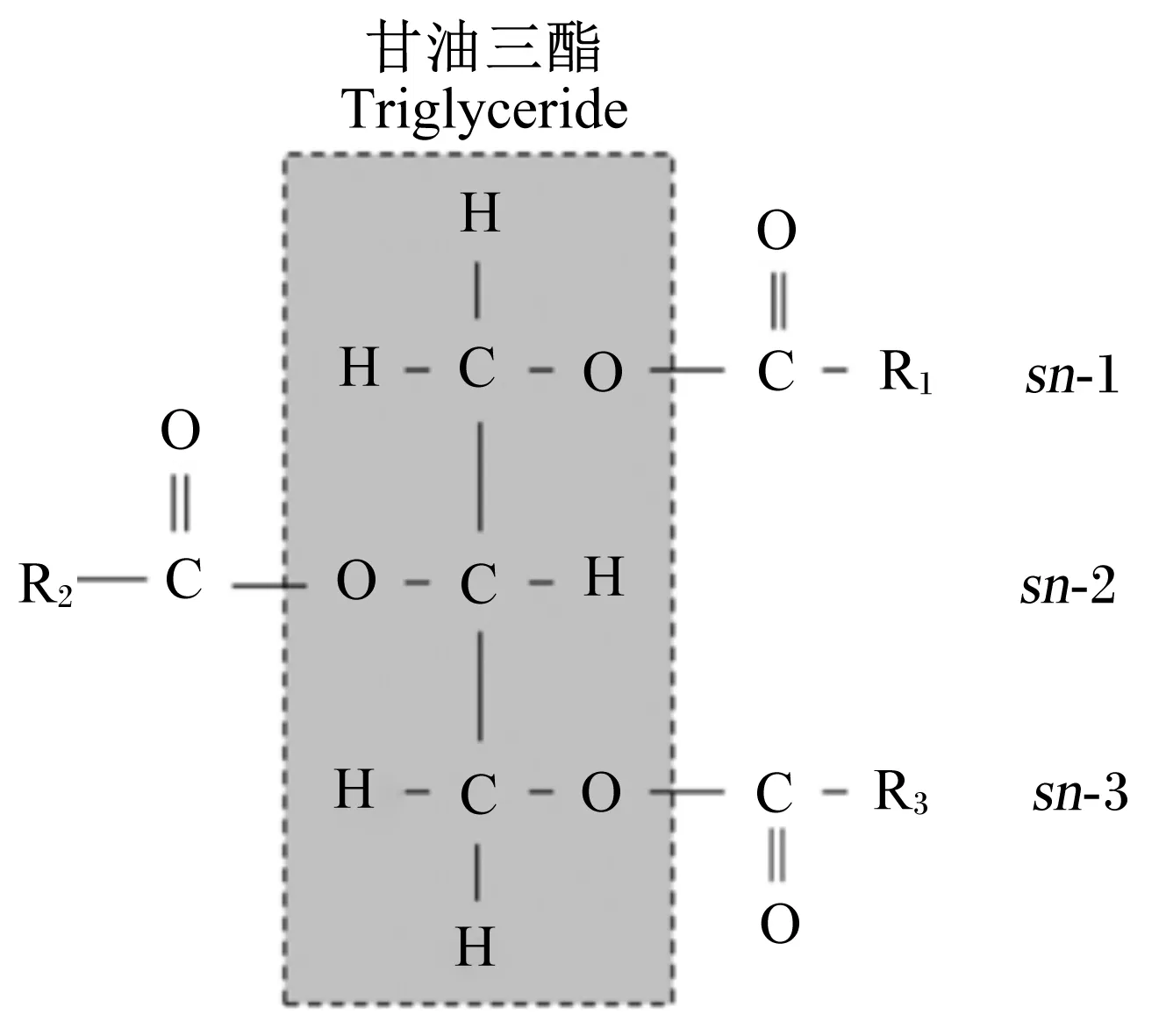
R表示脂肪酸,R1、R2、和R3表示脂肪酸分別在sn-1、sn-2和sn-3位酯化。
1.2 營養意義
甘油三酯的結構會影響油脂的理化特性及營養生理功能。近年圍繞甘油三酯脂肪酸不同酯化位置影響嬰兒對脂肪消化吸收的研究較為深入。Innis等[3,9-10]指出,人母乳甘油三酯內C16∶0特異結合于sn-2,這樣的結構不僅能夠防止嬰兒C16∶0吸收缺乏,同時在腸道和其他組織中當C18∶1cis-9增加時可節約C16∶0的釋放,從而維持脂肪的有效吸收利用,保證高效的能量正平衡,以維持嬰兒組織的凈生長。此外,研究指出sn-2位的脂肪酸與嬰兒腸道微生物區系的形成密切相關,其中sn-2位的C16∶0和C22∶6n-3(DHA)與多數腸道微生物相關[11-12]。
在成人脂肪營養相關研究中發現,甘油三酯脂肪酸酯化位置結構也會顯著影響脂肪酸的消化、吸收與代謝。長鏈脂肪酸在甘油三酯外側(sn-1或sn-3)時,會降低該脂肪酸乃至甘油三酯上的其他脂肪酸的消化、吸收與代謝[13]。研究發現,甘油三酯上脂肪酸酯化位置影響試驗小鼠體內脂肪酸的降解速率,sn-2位的棕櫚酸、油酸和亞麻酸相較于sn-1和sn-3位較慢被降解,而sn-2位的C20∶5n-3(EPA)較快被降解,DHA的降解速度不受酯化位置的影響[14]。體外胰脂肪酶水解試驗發現游離脂肪酸的釋放速率受不同油脂及脂肪酸甘油三酯酯化位置結構的影響[15]。在大鼠上的試驗得知sn-2位的DHA相較于sn-1和sn-3位消化率高[16],sn-3位的EPA可降低小鼠肝臟中甘油三酯含量,降低動脈壁炎癥的發生率[17]。
脂肪酸在甘油三酯上的不同酯化位置也會影響脂肪的加工及其他特性,從而間接影響營養品質。研究發現,甘油三酯脂肪酸酯化位置結構影響奶酪和奶油的物理特性和風味[18]及皮下脂肪的滑動性(slip point)和硬度[19],此外sn-3位的乙酸甘油酯(acTAG)可降低脂肪的黏稠度和熱值[20]。植物油脂肪酸的氧化穩定性與甘油三酯脂肪酸酯化位置結構相關,如sn-2位亞油酸的氧化穩定性高于sn-1和sn-3位[21];另有研究表明牛乳脂肪中的共軛亞油酸在甘油三酯的sn-2位酯化相對sn-1(或sn-3)位擁有更高的氧化穩定性[22]。此外,也有研究報道甘油三酯脂肪酸酯化位置結構與抗微生物特性相關,如sn-1和sn-2位的肉豆蔻酸甘油單酯均具有抗菌和抗真菌作用,且sn-1位強于sn-2位[23]。
2 牛奶甘油三酯脂肪酸酯化位置結構特點
通過對乳脂肪甘油三酯脂肪酸酯化位置的檢測發現,人母乳脂肪酸在甘油三酯內的結合位置存在較高的保守性,即飽和脂肪酸,尤其是C16∶0,特異結合在中心位置sn-2位上,而不飽和脂肪酸結合在外側位置sn-1和sn-3位上[6],然而,Jensen等[24]報道,牛奶甘油三酯上脂肪酸的結合位置卻并未呈現此規律(表1)。早期研究針對不同牛群來源乳脂肪以及采集季節對牛奶甘油三酯內脂肪酸的結合位置進行了分析,雖然得出的結論是牛奶中脂肪酸在甘油三酯上的酯化位置并非隨機,而是存在一定的特異性,并有學者推論,乳腺內合成甘油三酯時對脂肪酸具有特異的選擇性,并不受飼糧養分的影響,即脂肪的變化局限于甘油三酯數量的改變而不是其結構的變化[25]。但是,其他研究卻發現牛奶甘油三酯結構變異較為寬泛[26]。Fox等[27]總結了目前已知的牛奶甘油三酯的結構特性,并推論C12~C16首要酯化于sn-2位,而C4和C6首要酯化在sn-3位;C4和C18的含量可能從底物角度具有限速酯化作用,以保持乳脂肪在正常牛體溫條件下呈現液態的需求。然而對于一些微量具生物活性功能的脂肪酸,如多不飽和脂肪酸的報道較少,其他物種油脂,如植物籽實油脂中的C18∶4n-3和γ-亞麻酸主要酯化在甘油三酯的sn-2位[28]。同位素標記法研究表明,海洋原生生物合成的極長鏈多不飽和脂肪酸(VLCPUFA)首先酯化于磷脂,其次酯化于甘油三酯的sn-2位[29]。
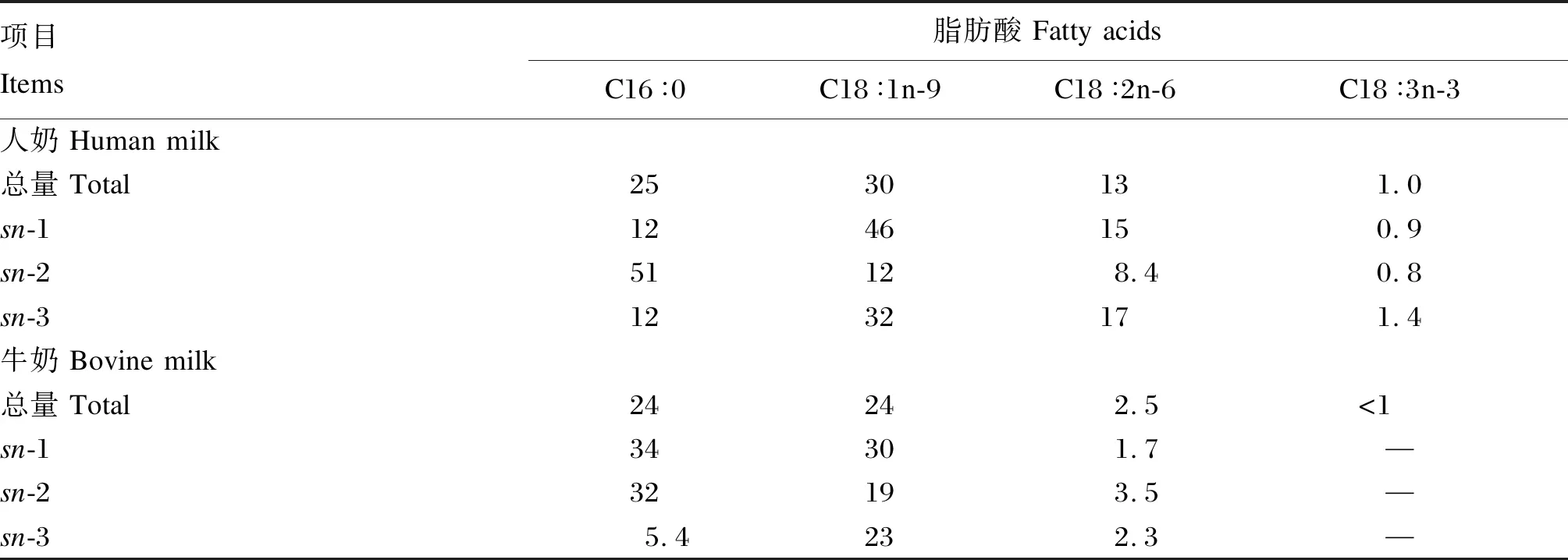
表1 人奶和牛奶中長鏈脂肪酸在甘油三酯中的酯化位置
3 乳腺合成代謝甘油三酯及其對牛奶甘油三酯結構特點的影響
3.1 前體物脂肪酸對牛奶甘油三酯酯化結構的潛在影響
了解乳成分前體物脂肪酸在牛乳腺酯化為甘油三酯的特點與規律可提升該營養途徑調控的準確性。近年在奶牛乳脂肪調控方面的研究證實,乳脂肪是乳成分中最易變化的組分。通過飼糧調控,乳脂率可產生上下3%的變化,Meta分析結果表明隨奶牛十二指腸十八碳脂肪酸流量增加,乳中相應脂肪酸產量均呈現線性或二次曲線升高[30]。另外,通過過瘤胃處理或直接瘤胃后灌注脂肪(或脂肪酸)的研究表明,除乳脂肪含量變化明顯外,脂肪酸組成也變化顯著,如亞油酸在乳脂肪中的比例由原來的19%升高至57%,亞麻酸的比例由0.6%升高至25.0%[31-32]。大量研究表明,乳腺對長鏈脂肪酸的攝取量升高時內源從頭合成脂肪酸的總量就會隨之降低[30-32],所以如果不降低總甘油三酯產量,而要提高某種脂肪酸的含量,那么在乳腺合成甘油三酯時,該脂肪酸就必須搶占更多酯化位置,但此規律與調控因素的研究暫無報道。
3.2 乳腺甘油三酯合成關鍵酶與調控
乳腺合成乳脂肪過程是營養底物與乳腺泌乳生理調控之間的平衡過程。平衡的水平表現于乳脂肪產量、含量與組成。乳腺內泌乳代謝效率與其調節作用決定了乳脂肪甘油三酯的結構和甘油三酯中總的酰基碳數(CN)(通常在26~54)[27]。反之,通過乳脂肪甘油三酯結構和甘油三酯內CN可反饋泌乳代謝效率與調節作用的表現。因此,了解二者的相關互作規律至關重要。
血漿中富含甘油三酯的乳糜微粒(CM)和極低密度脂蛋白(VLDL)是乳腺攝取長鏈脂肪酸的主要來源,而中短鏈脂肪酸主要源于乳腺內從頭合成[33-34]。哺乳動物乳腺合成甘油三酯主要通過三磷酸甘油途徑,暫無證據表明可由單酰甘油途徑酯化脂肪酸合成乳脂肪[27,35]。催化三磷酸甘油途徑合成甘油三酯的相關酶關聯于內質網和線粒體膜內外,研究發現在內質網上形成的初級脂滴內仍有較高的相關酶活性[33,36]。三磷酸甘油途徑主要由脂酰輔酶A:sn-3-磷酸甘油酰基轉移酶(glycerol-3-phosphate acyltransferases,GPATs)、1-乙酰甘油-3-磷酸酰基轉移酶(1-acylglycerol-3-phosphate acyltransferases,AGPATs)、磷脂酸磷酸酶(lipin phosphatidic acid phosphatase,PAP)和二甘油酯酰基轉移酶(diacylglycerol acyltransferases,DGATs)等序列式催化酶系組成[27,33]。目前在各組織細胞,包括乳腺細胞內質網上,三磷酸甘油途徑合成甘油三酯的酶催化過程和調控機制已了解地較為明晰,但在初級脂小滴內的催化過程和調控機制鮮有報道[33]。
三磷酸甘油的酰基反應是乳腺合成甘油三酯的第1個關鍵步驟,三磷酸甘油源于糖酵解途徑或者血液中的游離甘油代謝途徑[27]。該反應的關鍵酶是GPATs,它是甘油三酯合成過程中活性最低的酰基轉移酶,同時具有甘油三酯合成的限速調控作用[33]。雖然多數組織中微粒體GPATs是總GPATs活性的主要組成,并由磷酸化和去磷酸化作用激活或抑制,但甘油三酯合成中的該調節機制還尚不明確[34]。GPATs和AGPTAs使脂肪酸酯化于sn-1和sn-2位,即合成1,2甘油二酯-sn-3-磷酸,該磷脂酸占據生物合成脂類的中心,可進一步轉化為胞嘧啶二磷酸-甘油二酯,作為酸性磷脂類的前體物,亦或去磷酸化生成甘油三酯的主要前體物甘油二酯、磷脂酰絲氨酸、磷酸卵磷脂和磷脂酰乙醇胺[34]。GPATs的活性受營養底物和激素的調控,如脂肪細胞分化可增強其翻譯水平,同時受饑餓和采食以及磷酸化和去磷酸化的反饋調節[34]。在甘油三酯的合成中,磷脂酸的去磷酸化反應需要磷脂酸磷酸酯酶-1(PAP-1),該酶由胞漿運行至有脂肪酸和脂酰輔酶A的內質網,肝臟中該酶的活性受胰高血糖素、糖皮質激素、環腺苷酸(cAMP)和生長激素的刺激活化,而受胰島素抑制[33-34]。體外山羊乳腺細胞培養研究發現,短鏈脂肪酸中丙酸和丁酸可上調AGPAT6的表達,從而提高甘油三酯合成量[37]。另有多項研究報道,在多個物種中,油酸被GATPs催化后主要酯化于sn-1位,而棕櫚酸被首要酯化于sn-2位,但在奶牛乳腺中發現棕櫚酸在sn-1和sn-2位的酯化數量相當[5,24,38]。在較早的研究報道中指出泌乳奶牛乳腺中AGPATs可將C8~C18轉移至sn-2位,而不轉移C4~C6[39]。
DGATs酯化長鏈和短鏈脂肪酸于sn-3位[27]。研究指出,缺乏DGAT-1基因的小鼠不具備泌乳功能[40];同樣,在植物合成甘油三酯過程中發現DGAT-1是重要的限速酶[41]。對DGATs活性的調控在動物肝臟和脂肪組織中已有報道,但關于乳腺組織中的相關信息仍不明確。脂肪酸含量升高會提高該酶的表達,由此推測,在乳腺組織中該酶可能具有較高活性[27]。近年的研究表明在反芻動物乳腺組織或乳中體細胞內均可檢測到DGATs基因表達,并隨著底物脂肪酸的不同,該酶表達發生變化,初步證實了上述推測[37,42]。在山羊乳腺細胞培養體系中添加短鏈脂肪酸時DGAT-1表達上調,并且伴隨甘油三酯合成量增加,其中丙酸和丁酸的該作用更顯著[37]。另有研究表明,通過在泌乳奶牛飼糧中添加富含長鏈不飽和脂肪酸的油脂(大豆油或魚油),可提高血液多不飽和脂肪酸含量,并可下調乳腺來源體細胞內DGAT-1基因的表達[42]。
4 小 結
甘油三酯脂肪酸酯化位置結構與脂肪酸組成共同影響乳脂肪的營養特性。在甘油三酯合成代謝過程中,牛乳腺根據底物脂肪酸種類、數量、比例等不同條件,可相對靈活的調節其代謝程度和方向,從而合成不同結構的甘油三酯產物,進而影響乳脂肪的營養與加工等特性。但目前有關牛乳腺合成不同結構甘油三酯的規律及其與底物的互作尚不清晰。此外,乳腺細胞內甘油三酯合成關鍵酶的研究在逐步明朗,這為進一步研究奶牛乳腺細胞甘油三酯合成代謝提供了線索以及觀測目標。但在不同營養底物條件下,特別是血液來源脂肪酸前體物供給發生變化時,這些關鍵酶的活性及調節途徑的變化規律有待進一步研究總結。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
哲學評論(2021年2期)2021-08-22 01:53:34
今日農業(2021年11期)2021-08-13 08:53:34
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
中華詩詞(2019年7期)2019-11-25 01:43:04
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
