飼糧添加驢源羅伊氏乳桿菌對斷奶驢駒生長性能、營養物質表觀消化率、血清免疫球蛋白含量及直腸菌群結構的影響
2021-07-12 03:22:30馮培祥李光玉司華哲劉曉穎姜桂苗趙付偉劉晗璐
動物營養學報 2021年6期
王 卓 馮培祥 李光玉 司華哲 劉曉穎 姜桂苗 趙付偉 張 婷 劉晗璐*
(1.中國農業科學院特產研究所,吉林省特種經濟動物分子生物學省部共建國家重點實驗室,長春130112; 2.聊城大學驢產業科技協同創新中心,聊城252000;3.國家膠類中藥工程技術 研究中心,東阿阿膠股份有限公司,聊城252201)
驢為高附加值家畜,驢皮、驢肉、驢奶深受消費者喜愛,驢產品的消費潛力為驢特色養殖業的發展提供了極大動力。在驢的養殖中,斷奶期是非常重要的時期,飼糧改變、母子分離以及環境改變等均易導致免疫力降低。此外,斷奶會導致腸道形態和功能變化,從而影響小腸的消化和吸收能力[1-2]。
2020年7月1日起我國飼料生產中開始了全面禁止添加抗生素,這意味著依靠抗生素促生長和預防腸道性疾病發生的時代已經結束,這無疑將使國內養殖產業鏈產生一定變革。在初生和受應激動物飼糧中添加直接飼喂微生物(direct-fed microbials,DFM),可以提高飼料利用率和通過抑制病原體改善動物健康,并具有調節免疫功能、提高其他有益微生物的增殖、調節胃腸道微生物區系平衡(GIT)等作用,常被用來代替或減少抗生素的使用[3-5]。將DFM作為提高動物生長性能和促進動物健康的功能性食品添加劑的研究也逐漸增加。然而,DFM在食草單胃動物驢的研究較少。
乳酸菌是應用最為廣泛的DFM之一[6-7]。Canzi等[8]證實,乳酸菌對兔微生物區系平衡的恢復具有積極作用。本實驗室從健康成年驢腸道內分離和鑒定了1株羅伊氏乳桿菌L2-2,經過小鼠安全性驗證證實了該菌的安全性,并且有促進小鼠胸腺發育的作用[9]。因此,本試驗旨在研究飼糧添加羅伊氏乳桿菌L2-2對斷奶驢駒生長性能、營養物質表觀消化率、血清免疫球蛋白含量及直腸菌群結構的影響,為驢源微生態制劑的開發和利用提供理論依據。
1 材料與方法
1.1 試驗菌株
從健康成年驢腸道中分離、篩選、鑒定后得到羅伊氏乳桿菌L2-2,現保藏于中國農業科學院特產研究所特種動物營養與飼養科技創新團隊實驗室。取復蘇的菌株接種到乳酸細菌培養基(MRS)固體斜面培養基,37 ℃培養24 h取出,刮取菌泥,用15%脫脂乳粉作為保護劑凍干,平板計數法測出有效活菌數。
1.2 試驗動物和試驗設計
選擇5~6月齡雌性斷奶驢駒10頭,平均體重為(113.05±14.66) kg,隨機分成2組,每組5個重復。對照組(CT組)喂基礎飼糧;試驗組(LR組)在基礎飼糧中添加羅伊氏乳桿菌,按1×1011CFU/kg添加于精飼料中。精飼料每日按照驢駒體重的1.3%定量飼喂,每日07:00和16:00飼喂精飼料,粗飼料(豆秸)自由采食。預試期7 d,正試期60 d。所有驢單圈飼養。精飼料組成及營養水平見表1,粗飼料營養水平見表2。
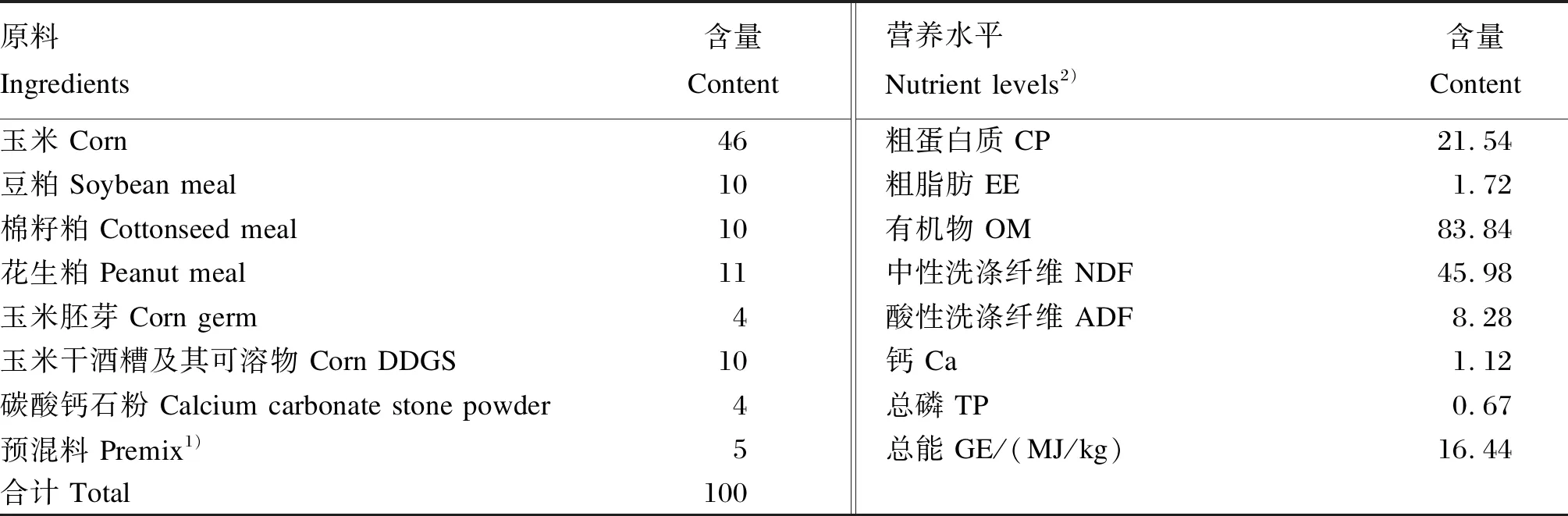
表1 精飼料組成及營養水平(風干基礎)

表2 粗飼料營養水平(風干基礎)
1.3 檢測指標
1.3.1 生長性能
每天記錄采食量,每15 d晨飼前稱量試驗驢體重,每隔3 d對每頭驢精飼料和粗飼料投料量和剩料量進行記錄,用于精確計算試驗期間每頭驢采食量、平均日增重(ADG)和料重比(F/G)。
1.3.2 營養物質表觀消化率
試驗期間連續3 d每日14:00小心收集每頭驢駒的部分新鮮糞便,避免驢毛、沙粒等污染,收集的糞樣稱重后按鮮重的5%加入10%硫酸以避免氮的損失。糞便樣品在80 ℃下殺菌2 h,然后降到65 ℃烘干48 h。分析飼糧和糞便樣品中干物質(DM)、有機物(OM)、粗蛋白質(CP)、鹽酸不溶灰分、粗脂肪(EE)、中性洗滌纖維(NDF)、酸性洗滌纖維(ADF)含量[10]。DM、CP、EE、NDF和ADF的表觀消化率采用2 mol/L鹽酸不溶灰分法[11]測定,通過以下公式計算表觀消化率:
D=100-[100×(A/A1)×(B1/B)]。
式中:D為營養物質表觀消化率;A為飼糧中鹽酸不溶性灰分含量;A1為糞便中鹽酸不溶性灰分含量;B為飼糧中該營養物質含量;B1為糞便中該營養物質含量。
1.3.3 血清免疫球蛋白含量
試驗第45天,晨飼前對驢頸靜脈采血10 mL,加入帶分離膠的促凝采血管中,離心(3 500 r/min,10 min)獲得血清樣本,-20 ℃保存,待分析驢駒的血清免疫球蛋白G(IgG)、免疫球蛋白A(IgA)、免疫球蛋白M(IgM)含量,使用酶聯免疫吸附試驗(ELISA)試劑盒測定,試劑盒購自南京建成生物工程研究所。
1.3.4 直腸菌群結構
試驗第50天采集直腸內容物20 g,無菌操作下攪拌混勻,取5 g直腸內容樣本,使用Fast DNA Spin Kit for Feces試劑盒(美國MP公司)進行微生物組DNA提取。提取后的DNA送往北京諾禾致源科技股份有限公司,使用IonS5TMXL測序平臺對微生物基因組的16S rRNA V3~V4區進行測序。所用引物為341F(5′-CCTAYGGGRBGCASCAG-3′)和806R(5′-GGACTACNNGGGTATCTAAT-3′)。在進行分析之前,用Cutadapt[12](Martin M. 2011)軟件對數據進行過濾和質控,對數據進行了均一化處理(normalize),應用Silva132數據庫進行微生物分類注釋。使用UPARSE軟件[13]根據97%的相似度對序列進行操作分類單元(OTUs)聚類,并在聚類的過程中去除單序列和嵌合體[14]。QIIME 1.9.1用來計算α-多樣性指數(ACE指數、Chao1指數、Simpson指數和Shannon指數)[15]。
1.4 數據分析
使用SPSS 21.0軟件進行統計分析。所有數據采用Independent-Samplest檢驗分析差異顯著性。數據用平均值±標準差表示,P<0.05表示差異顯著。
2 結果與分析
2.1 飼糧添加羅伊氏乳桿菌對斷奶驢駒生長性能的影響
如表3所示,對照組和試驗組的15、30、45和60 d體重以及平均日增重、料重比均無顯著差異(P>0.05)。試驗組的粗飼料采食量顯著低于對照組(P<0.05)。
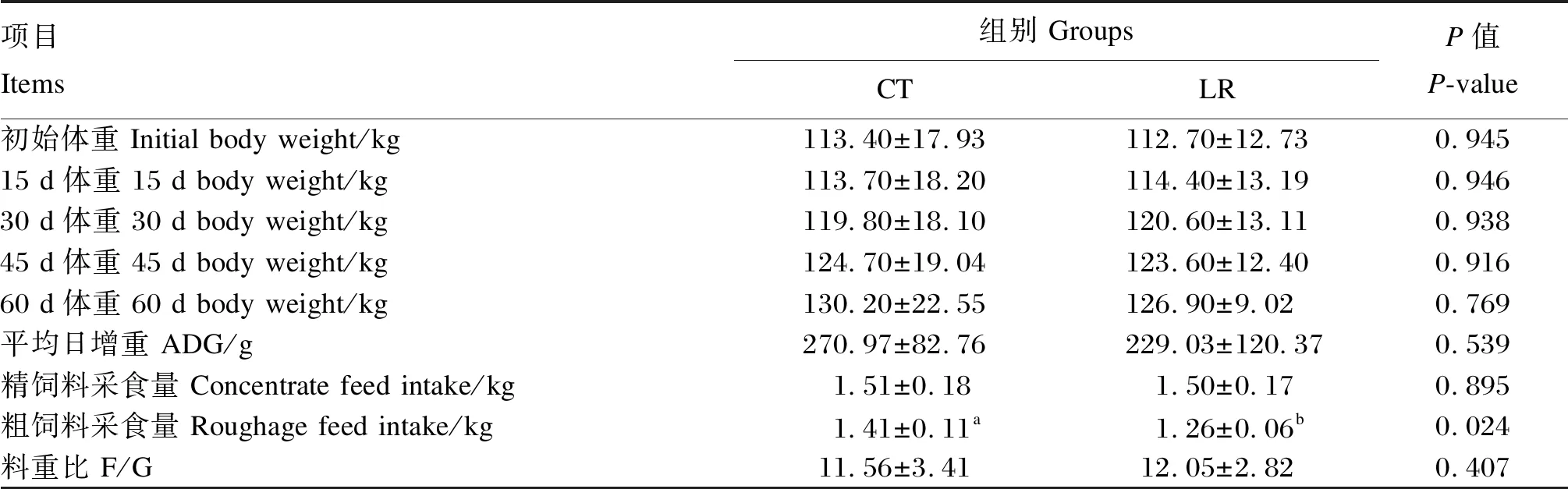
表3 飼糧添加羅伊氏乳桿菌對斷奶驢駒生長性能的影響
2.2 飼糧添加羅伊氏乳桿菌對斷奶驢駒營養物質表觀消化率的影響
如表4所示,對照組和試驗組的DM、CP、EE、ADF、NDF表觀消化率均無顯著差異(P>0.05)。與對照組相比,試驗組的ADF表觀消化率提高了6.2%,NDF表觀消化率提高了3.4%。
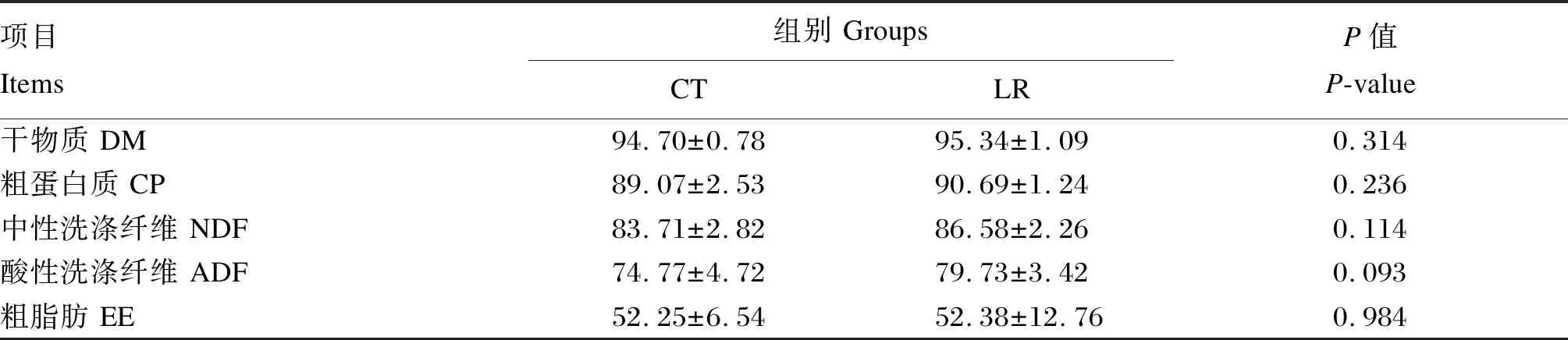
表4 飼糧添加羅伊氏乳桿菌對斷奶驢駒營養物質表觀消化率的影響
2.3 飼糧添加羅伊氏乳桿菌對斷奶驢駒血清免疫球蛋白含量的影響
如表5所示,試驗組的血清IgA含量顯著高于對照組(P<0.05)。對照組和試驗組的血清IgG和IgM含量均無顯著差異(P>0.05)。
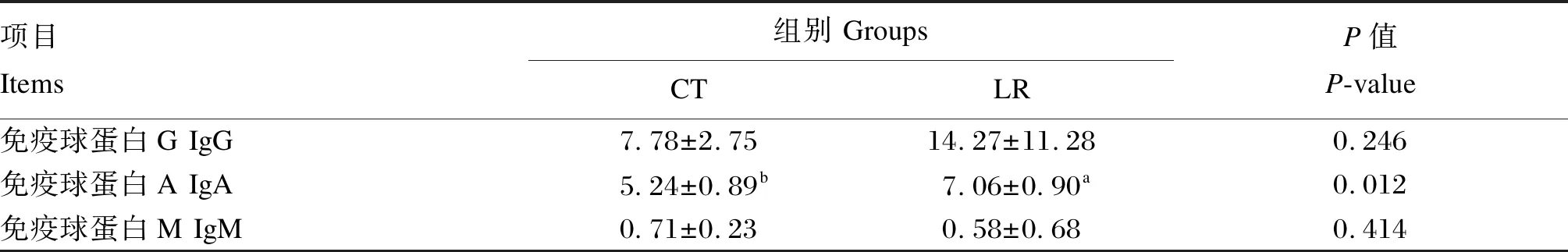
表5 飼糧添加羅伊氏乳桿菌對斷奶驢駒血清免疫球蛋白含量的影響
2.4 飼糧添加羅伊氏乳桿菌對斷奶驢駒直腸菌群結構的影響
如表6所示,對照組和試驗組的覆蓋度指數均高于0.995,說明測序深度足夠,結果可呈現99.5%以上的菌群組成。試驗組的Shannon指數和Simpson指數高于對照組,Chao1指數和ACE指數低于對照組,但差異不顯著(P>0.05)。這表明飼糧添加羅伊氏乳桿菌對斷奶驢駒直腸菌群α-多樣性指數無顯著影響。
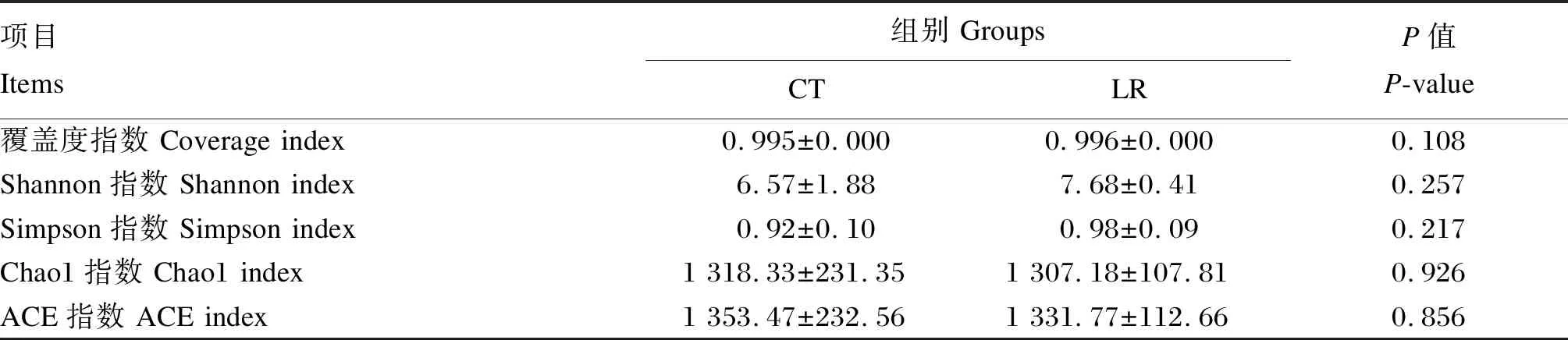
表6 飼糧添加羅伊氏乳桿菌對斷奶驢駒直腸菌群α-多樣性指數影響
按97%相似度聚類后得到2 369個OTUs,對照組獨有的OTUs為250個,試驗組獨有的OTUs為245個。飼糧添加羅伊氏乳桿菌后,斷奶驢駒直腸菌群組成的門水平和屬水平上微生物組成如圖1和圖2所示。對照組和試驗組的直腸菌群在門水平上主要歸類為厚壁菌門[(54.76±10.91)%]、擬桿菌門[(27.74±12.60)%]和變形菌門[(10.71±17.00)%]。試驗組的擬桿菌門相對豐度[(35.97±7.68)%]高于對照組[(19.52±11.37)%],對照組中有2頭斷奶驢駒的變形菌門相對豐度高達36.46%~48.45%,這表明飼糧添加羅伊氏乳桿菌會提高斷奶驢駒直腸擬桿菌門相對豐度,可能會抑制變形菌門相對豐度。在屬水平上,對照組和試驗組中相對豐度較高的菌群分別為不動桿菌屬[(9.07±17.28)%]、土壤芽孢桿菌屬[(4.78±6.00)%]、鏈球菌屬[(6.34±6.63)%]、未鑒定瘤胃球菌科[(3.54±1.96)%]、未鑒定梭菌目[(2.60±4.17)%]。其他菌屬相對豐度均低于0.1%,在對照組和試驗組的其他菌屬分別占(54.65±28.70)%和(79.30±10.81)%。對照組中2頭斷奶驢駒的變形菌門相對豐度較高,其不動桿菌屬相對豐度也較高,達到34.35%~48.12%。試驗組的厚壁菌門中未鑒定瘤胃球菌科相對豐度[(4.82±1.80)%]高于對照組[(2.26±1.15)%]。對照組和試驗組的直腸菌群在其他屬水平上無明顯區別。

CT1~CT5代表對照組的5頭斷奶驢駒,LR1~LR5代表試驗組的5頭斷奶驢駒。圖2同。
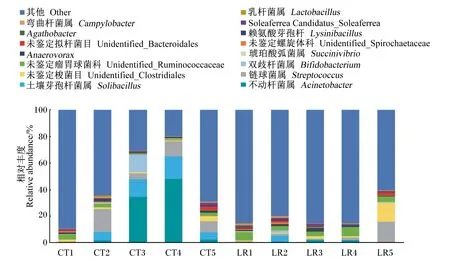
圖2 斷奶驢駒直腸菌群屬水平相對豐度
如圖3所示,主坐標分析(PCoA)顯示2個軸相加解釋79.80%的變量組成,2組之間一些樣品的菌群組成相似,但仍有一些差異,這表明飼糧添加羅伊氏乳桿菌對斷奶驢駒直腸的菌群組成具有一定影響。
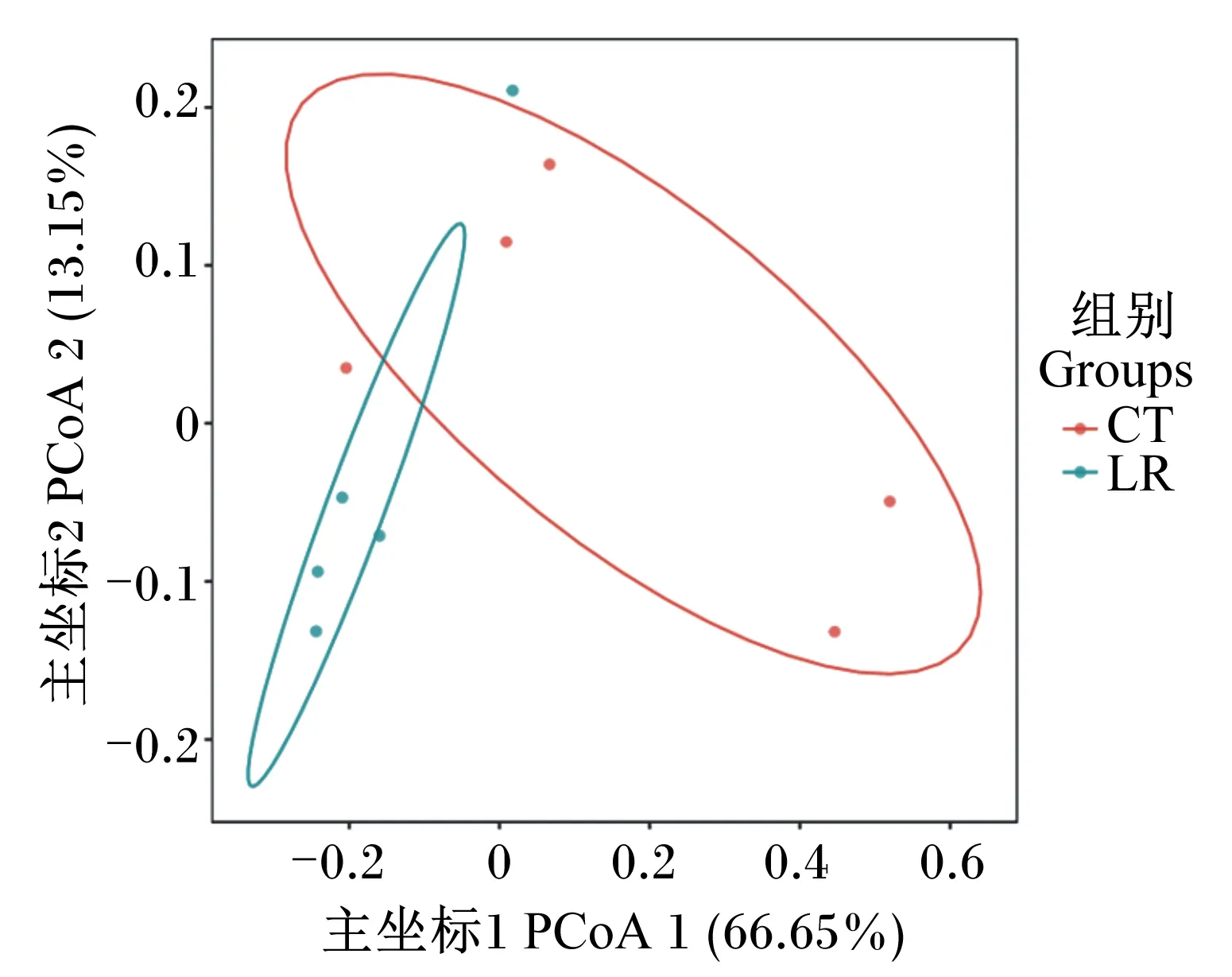
圖3 斷奶驢駒直腸菌群主坐標分析
3 討 論
DFM被認為可以通過定性或定量影響宿主腸道微生物群,同時幫助宿主完成各種生理生化功能[8]。DFM可以為宿主提供大量微生物菌體蛋白,可以改善營養物質消化率,通過微生物分泌的消化酶和提供維生素B族以及各種酶的輔助因子而提高宿主消化能力[16-18]。而有關飼喂乳酸菌后對動物益生作用的研究結果并不一致,這可能與動物的品種、狀態以及菌的功能和來源有關。Ellinger等[19]、Abu-Tarboush等[20]、Krehbiel等[3]和Raeth-Knight等[21]報道,在飼喂乳酸桿菌后DM攝入量、ADG以及飼料轉化率沒有改善。Krehbiel等[3]報道,飼糧中添加DFM增加了瘤胃中纖維素降解菌的數量和比例。Giang等[1]報道,在仔豬斷奶后的前2周補充復合乳酸菌顯著增加了CP和粗纖維表觀消化率。而本試驗結果表明,飼糧添加羅伊氏乳桿菌后,粗飼料采食量顯著減少,驢駒的生長性能未受到影響,同時發現驢駒的NDF和ADF表觀消化率分別提高了3.4%和6.2%。因此,飼糧添加羅伊氏乳桿菌可以直接或間接增加驢駒對纖維素的消化率,同時可增加驢駒的飽腹感,在生產中有利于減少飼料成本的投入。
腸道細菌通過代謝產物參與宿主的免疫系統[22]。口服乳酸桿菌可以增強先天免疫反應(即增強吞噬作用和自然殺傷細胞活性),以及提高人類和動物中血清IgA含量和減少血清IgE含量[23-24]。血清免疫球蛋白是體液免疫系統的主要成分。本研究也發現,飼糧添加羅伊氏乳桿菌可以提高斷奶驢駒的血清IgA含量。IgA是黏膜免疫的主要抗體,其主要功能是在非特異性免疫防護機制的協助下減少病原菌。本實驗室也研究發現,將驢源羅伊氏乳桿菌連續灌胃小鼠7 d后,羅伊氏乳桿菌可以通過增加小鼠胸腺指數來增強小鼠的免疫功能[9]。
從斷奶驢駒直腸菌群組成的門水平和屬水平的試驗結果發現,飼糧添加羅伊氏乳桿菌后,乳桿菌屬相對豐度并沒有明顯增加,這可能是直接飼喂裸菌后細菌經過胃腸液消化并沒有有效存活,而是通過細菌的代謝產物發揮作用。在模擬胃酸環境下,單純沒有被包被的益生菌很難在胃中存活2 h[25]。但我們觀察到飼糧添加羅伊氏乳桿菌后刺激了斷奶驢駒直腸中門水平上擬桿菌門和屬水平上未鑒定瘤胃球菌科的增長。擬桿菌門是飼料碳水化合物和蛋白質的重要降解菌,可促進胃腸道免疫系統的發育[26]。有研究表明,瘤胃球菌科可降解多糖和纖維,產生大量揮發性脂肪酸(VFA),這可能有助于通過增強飽腹感減少食物攝入量[27-29]。本試驗也發現在飼糧添加羅伊氏乳桿菌后斷奶驢駒粗飼料采食量減少,ADF表觀消化率提高,可能與未鑒定瘤胃球菌科相對豐度增加有關。瘤胃球菌科已被證明可在體外產生抗炎肽[30],而且可以降低小牛的腹瀉發病率和死亡率[31]。此外,瘤胃球菌科的數量與結腸炎、絞痛以及應激因素如禁食、運輸和麻醉有顯著負相關[32-34]。本試驗中,對照組2只斷奶驢駒直腸內不動桿菌的相對豐度較高。Regalado等[35]研究表明,不動桿菌可引起動物胃腸炎的發生。本試驗中,試驗組并未發現這樣的個體,這也從側面證明了飼糧添加羅伊氏乳桿菌可在一定程度上維持腸道穩態。因此,羅伊氏乳桿菌可能通過間接增加未鑒定瘤胃球菌科的相對豐度而預防和治療腹瀉等胃腸道疾病,關于這一點還需要進一步研究確定。
4 結 論
研究表明,飼糧添加羅伊氏乳桿菌可以降低斷奶驢駒的粗飼料采食量,提高ADF和NDF表觀消化率,增加血清IgA含量,提高直腸中擬桿菌門和未鑒定瘤胃球菌科相對豐度。
