水分脅迫對三葉青葉綠體超微結構及黃酮合成關鍵酶的影響
2021-07-12 10:38:42屠玲艷吳學謙許海順
浙江農林大學學報 2021年3期
屠玲艷,吳學謙,許海順
(浙江農林大學 省部共建亞熱帶森林培育國家重點實驗室,浙江 杭州 311300)
三葉青Tetrastigmahemsleyanum為葡萄科Vitaceae崖爬藤屬Tetrastigma的藥用植物,主要以地下塊根入藥。在臨床中,三葉青內服可治療咽喉炎、肝炎、肺炎、小兒高熱驚厥及病毒性腦膜炎等炎癥,外用可治毒蛇咬傷、跌打損傷、癰疽等疾病,被譽為“植物抗生素”,是中國獨有的珍貴藥材,主要分布于浙江、湖南、廣東、廣西、四川等地[1]。由于三葉青具有良好的抗腫瘤功效且無毒副作用[2],導致三葉青需求量大量增加,人工栽培面積不斷擴大。人工種植三葉青多采取遮光大棚霧噴管理,藤葉生長茂盛,但塊根品質卻遠不如生長環境苛刻的野生三葉青[3?5]。逆境脅迫雖抑制植物的生長發育,特別是營養生長,但同時也會促進或者抑制植物中黃酮等一些次生代謝產物的累積[6?9]。三葉青喜陰濕環境,野生三葉青多生長于山坡或山溝、溪谷兩旁的針闊混交林或雜木林林下背陰處[10?11],可見水分條件對三葉青的生長影響較大。目前,溫度[12?13]、光照[14?15]、土壤肥料[16?17]等環境因子對三葉青生長的影響已有研究,但關于水分脅迫對三葉青特別是三葉青黃酮含量的影響還未見報道。因此,本研究分析了水分脅迫對三葉青生長、黃酮含量及其合成途徑中關鍵酶的影響,為人工種植高品質三葉青提供理論依據。
1 材料與方法
1.1 供試材料
三葉青由浙江五養堂藥業有限公司遂昌縣金竹藥王谷三葉青基地提供。為了保持試驗材料的一致性,以專用袋標準化繁育的2年生三葉青植株作為供試材料。
1.2 材料處理
實驗地位于浙江農林大學東湖校區 (30°15′28″N,119°43′35″E),該區全年降水量 1 628.6 mm,全年平均氣溫16.4 ℃,年日照時數1 847.3 h,無霜期237.0 d。取長勢良好的2年生的三葉青植株,種植在上口徑20 cm、下口徑15 cm、高20 cm的塑料花盆中,栽培基質為過篩土∶營養基質=2∶1(質量比),每盆1株,緩苗5周,緩苗期間定時定量澆水。5周后,選取長勢基本一致的2年生植株57株,隨機分為3組,每組19盆,設置3個處理組,分別為干旱、水澇和對照。干旱組在試驗期間一直不澆水,水澇組保證地下部分在試驗期間一直處于淹水狀態,對照組在試驗期間正常澆水。
1.3 樣品獲取
干旱組在脅迫處理的第1、8、16、24、32、40天,于相同葉位隨機取樣3盆(預試驗發現三葉青在不澆水40 d左右出現死亡現象),水澇組在脅迫處理的第1、4、8、12、16、20天,于相同葉位隨機取樣3盆(預試驗發現三葉青在水淹20 d左右出現死亡現象),對照組在脅迫處理的第1、4、8、12、16、20、24、32、40天,于相同葉位處隨機取樣3盆。每個試驗組設置3個重復,所有樣品取樣后立即用清水洗凈擦干后再立即用液氮預冷,于?80 ℃冰箱保存待測。
電鏡樣品取樣,在脅迫處理最后1 d,取植株的中部成熟葉片,中間靠近大葉脈處部位切成長0.1 cm、寬0.2 cm左右的小片(避開大葉脈)后迅速放入體積分數為2.5%的戊二醛溶液中,抽真空至材料沉入固定液底部,4 ℃固定過夜,每個試驗組重復3次。
1.4 研究方法
1.4.1 三葉青葉片及塊根表型拍攝 隨機取實驗三葉青進行葉片表型拍攝,再取塊根,洗凈后擦干進行塊根表型拍攝。
1.4.2 葉綠體超微結構電鏡觀察預處理 倒掉固定液,用0.1 mol·L?1pH 7.0的磷酸緩沖液漂洗樣品3次,每次15 min;用質量分數為1%的鋨酸固定樣品1~2 h后漂洗3次。然后用不同體積分數的乙醇溶液(30%、50%、70%、80%、90%和95%)對樣品進行脫水處理,每種體積分數處理15 min,再用純乙醇處理20 min,最后用純丙酮處理20 min。接著用Spurr包埋劑與丙酮的混合液(V/V=1/1)處理樣品1 h;再用Spurr包埋劑與丙酮的混合液(V/V=3/1)處理樣品3 h;最后用純包埋劑處理樣品過夜。將經過滲透處理的樣品包埋起來,70 ℃加熱過夜,即得到包埋好的樣品;樣品在LEICA EM UC7型超薄切片機中切片,獲得70~90 nm的切片;切片經檸檬酸鉛溶液和醋酸雙氧鈾50%乙醇飽和溶液(體積分數)各染色5~10 min。最后在Hitachi H-7 650型透射電鏡中觀察葉肉細胞中的葉綠體。
1.4.3 黃酮質量分數測定 黃酮質量分數的測定方法參照文獻[18?19]。以蘆丁不同質量分數(mg)為縱坐標(y),以波長500 nm下的吸光度D(500)為橫坐標(x)繪制標準曲線,得到回歸方程y=0.245 9x+0.001 6(R2=0.999 9)。測定樣品在500 nm波長下的吸光度D(500),計算三葉青總黃酮質量分數,每個樣品3個重復。
1.4.4 黃酮合成關鍵酶活性測定 酶液制備及苯丙氨酸解氨酶(PAL)活性的測定參照徐琳煜[20]的方法,以每克鮮質量葉片每分鐘波長290 nm下的吸光度D(290)變化0.01為1個酶活力單位(16.67 nkat·g?1·min?1)。查爾酮合成酶 (CHS)活性測定參照試劑盒法(上海晶抗生物工程):以標準物的濃度(×16.67 nkat·L?1)為縱坐標(y),波長450 nm下的吸光度D(450)為橫坐標(x),得到標準曲線直線回歸方程y=92.238x+1.193 8(R2=0.999),然后根據樣品的D(450)計算樣品相應的濃度。查爾酮異構酶(CHI)活性測定參照試劑盒法(上海晶抗生物工程):以標準物的濃度(×16.67 nkat·L?1)為縱坐標(y),波長450 nm下的吸光度D(450)為橫坐標(x),得到標準曲線直線回歸方程y=1 165.3x+50.552(R2=0.999),然后根據樣品的D(450)計算出樣品相應的濃度。
1.5 數據處理
用Excel進行數據處理與制圖;運用單因素方差分析和最小顯著極差法LSR進行方差分析和多重比較 (α=0.05);采用 Pearson’s進行相關性分析。
2 結果與分析
2.1 水分脅迫下三葉青葉片和塊根表型特征
水分脅迫嚴重影響三葉青的營養生長。從葉片表型來看,未遭受水分脅迫的三葉青(對照)葉片飽滿,色澤鮮艷,葉脈清晰;干旱脅迫40 d后,三葉青葉片失水,萎蔫,逐漸發黃枯萎;水澇脅迫20 d后,三葉青葉片失綠變黃(圖1)。從塊根的表觀形態上來看,對照組的塊根飽滿,形狀完好,而干旱組的塊根失水皺縮,表皮布滿溝壑,水澇組的塊根綿軟,表皮極易脫落,塊根幾乎腐爛(圖2)。

圖 1 水分脅迫對三葉青葉片的影響Figure 1 Effects of water stress on leaves of T.hemsleyanum

圖 2 水分脅迫對三葉青塊根的影響Figure 2 Effects of water stress on roots of T.hemsleyanum
2.2 水分脅迫下三葉青葉綠體超微結構特征
葉綠體結構的完整性及其內部結構的有序性對光合作用的光能吸收和傳遞有著重要作用。與對照相比,水澇和干旱處理下葉綠體數目明顯減少(圖3A1,圖3B1,圖3C1)。水分脅迫下,三葉青葉片中的葉綠體在細胞內的排列方式發生變化,與對照的緊貼細胞壁并且整齊分布相比,水澇和干旱處理后,葉肉細胞中的葉綠體與細胞壁分離,擠向液泡,在細胞中呈現擁擠混亂的現象,葉綠體向細胞中間靠攏,完全隨機分布在細胞中(圖3A1,圖3B1,圖3C1)。水澇和干旱處理后,葉肉細胞葉綠體膨脹,失去原本形態,葉綠體中淀粉粒數量增多,可能是代謝不正常導致淀粉粒無法運出造成的(圖3A2,圖3B2,圖3B3)。與對照相比,水澇和干旱處理后,葉綠體基質顏色變淺,葉綠體膨大,質體小球顏色變淺,數量增多,體積變大(圖3A3,圖3B3,圖3C3)。對照基粒片層結構整齊,緊密排列,而干旱和水澇處理后,基粒片層失去整齊緊密結構,與細胞質之間不再有完整界限,結構被破壞(圖3A4,圖3B4,圖3C4)。

圖 3 水分脅迫對三葉青葉片葉綠體超微結構的影響Figure 3 Effects of water stress on chloroplast ultrastructurey of leaves in T.hemsleyanum
2.3 水分脅迫對三葉青塊根黃酮質量分數的影響
水分脅迫影響三葉青塊根的黃酮質量分數。水澇脅迫8 d后,總黃酮質量分數顯著高于對照(P<0.05),在脅迫12 d時達到最大值。干旱脅迫16 d后,總黃酮質量分數顯著高于對照(P<0.05),在脅迫16 d時達到峰值(圖4)。對照總黃酮質量分數保持在比較穩定的范圍內,而水澇和干旱處理后,總黃酮質量分數呈先低后高再回落的變化趨勢。

圖 4 水分脅迫下三葉青塊根黃酮質量分數的變化Figure 4 Change of content of flavonoid in T.hemsleyanum under water stress
2.4 水分脅迫對三葉青塊根黃酮合成關鍵酶活性的影響
水分脅迫對三葉青塊根黃酮合成關鍵酶活性的影響較大。水澇脅迫4 d后,PAL活性顯著提高(P<0.05),并且在脅迫4 d時達到峰值,之后下降且在脅迫16 d時顯著低于對照(P<0.05)。干旱脅迫16 d后,PAL活性顯著高于對照(P<0.05),并且在脅迫16 d時達到峰值,之后下降且在脅迫32 d時顯著低于對照(P<0.05)。即PAL活性呈先上升后下降的趨勢,脅迫處理前期大于對照,脅迫后期則低于對照 (圖 5)。
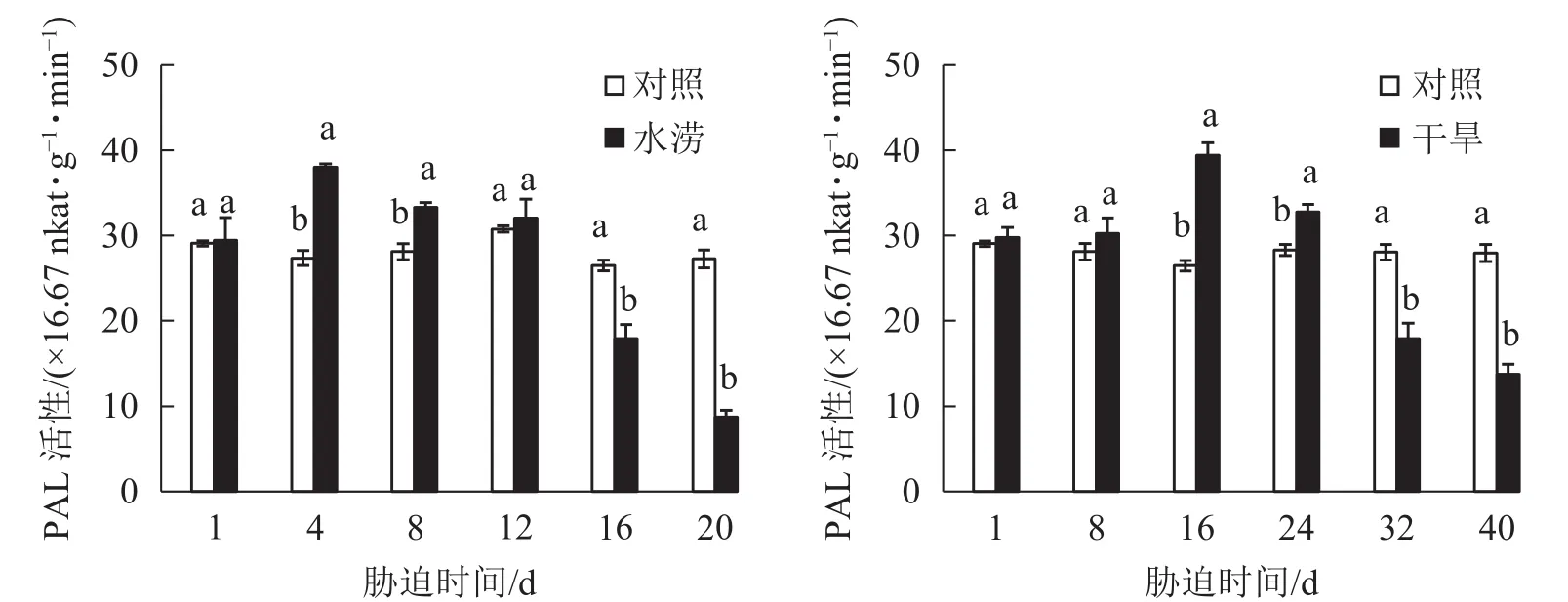
圖 5 水分脅迫下三葉青塊根PAL活性的變化Figure 5 Change of activities of PAL in T.hemsleyanum under water stress
水分脅迫影響三葉青CHS活性。水澇脅迫4 d后,CHS活性顯著高于對照(P<0.05),且在脅迫處理的12 d達到最大值,之后回落到與對照近似水平。干旱脅迫16 d后,干旱處理的CHS活性顯著增強(P<0.05),并且在脅迫16 d達到峰值,之后下降至與對照近似水平。整個研究過程中,水澇和干旱處理的CHS活性呈先上升后下降的趨勢,且酶活性大于對照組,對照組基本不變(圖6)。

圖 6 水分脅迫下三葉青塊根CHS活性的變化Figure 6 Change of activities of CHS in T.hemsleyanum under water stress
與對照相比,水澇脅迫12 d后,CHI活性顯著增強(P<0.05),并且在脅迫處理的12 d達到峰值,之后開始下降。干旱脅迫下,CHI活性在8~16 d迅速增加,且在脅迫16 d顯著高于對照(P<0.05),之后開始下降(圖7)。總體來說,水澇和干旱的CHS活性呈先上升后下降的趨勢,且酶活性大于對照組,而對照組的CHI活性一直比較穩定。此外,相比于CHS活性,CHI對脅迫的響應略有延緩,說明CHI可能在CHS之后起作用。

圖 7 水分脅迫下三葉青塊根CHI活性的變化Figure 7 Change of activities of CHI in T.hemsleyanum under water stress
2.5 水分脅迫下黃酮質量分數與黃酮合成關鍵酶的相關性分析
從表1可以看出:干旱和水澇脅迫下,三葉青黃酮質量分數、PAL、CHS、CHI活性之間均表現為正相關關系,個別呈顯著相關(P<0.05),表明黃酮質量分數受酶活性的調控。脅迫處理組的PAL與CHS、CHI之間呈正相關,但相關不顯著(P>0.05);CHS和CHI之間呈極顯著相關(P<0.01),表明CHS和CHI共同促進黃酮質量分數增加。

表 1 水分脅迫下黃酮質量分數、PAL、CHS、CHI之間的相關性分析Table 1 Correlation analysis between flavonoid content, PAL, CHS and CHI under water stress
3 討論與結論
水分影響三葉青的光合作用,而葉綠體作為光合作用的反應場所,其結構的完整性是影響光合作用正常運行的關鍵。本研究在三葉青葉綠體超微結構中發現:脅迫使三葉青葉綠體數量減少且不斷向細胞中央靠攏,可能是三葉青葉片發黃的原因。王順才等[21]在研究干旱脅迫對3種蘋果屬Malus植物的葉綠體超微結構的影響時也證實了這一點。本研究發現:干旱和水澇下,葉綠體形態結構發生改變,葉綠體膨大變圓,淀粉粒大量積累無法運出,質體小球的數量增多、體積變大、顏色變淺。楊鳳軍等[22]研究了干旱脅迫對番茄Lycopersiconesculentum葉面葉綠體超微結構后表明:逆境導致質體小球數量增加;因為質體小球是類囊體降解脂類聚集的結果,其數量的變化可作為葉細胞受損的標志[23]。本研究還觀察到水澇和干旱的三葉青葉綠體基粒片層失去整齊緊密排列結構,變得松散模糊,沒有界限,這與李冬林等[24]的研究結果一致;表明水澇和干旱使三葉青葉片葉綠體膜結構嚴重受損,失去生理活性,從而影響三葉青的光合作用和營養生長。
次生代謝過程是連接生態環境與藥效成分含量的中間環節,產生的次生代謝產物在植物自我保護、生理調節等生命活動方面起著重要作用[25],并且是藥用植物的主要藥效成分。環境影響次生代謝產物的形成和積累[26]。目前,關于非生物因子影響藥用植物次生代謝產物的研究已有很多,ZHU等[27]研究認為:柴胡Bupleurumchinense在水分脅迫下通過增加次生代謝產物的含量來提高抗氧化能力,進而抵制因水分脅迫產生的自由基;有研究發現:低溫培養的黃豆Glycinemax,其大豆黃素和染料木苷的代謝水平顯著增高[28];LI等[29]在研究擬南芥Arabidopsis thaliana應對鹽脅迫的響應時發現:黃酮類化合物含量上升。三葉青作為中藥材,其藥用活性成分主要有黃酮類化合物、酚酸類化合物、萜類化合物等次生代謝產物,本研究發現:水分脅迫促進了三葉青黃酮的積累,導致總黃酮質量分數高于對照,說明水澇和干旱使三葉青黃酮代謝增強,增加了三葉青塊根單位干質量的黃酮質量分數。小麥Triticum aestivum、丹參Salviamiltiorrhiza等在水分脅迫下同樣出現了黃酮類化合物質量分數上升的情況[30?31]。
PAL是連接初級代謝和苯丙烷類代謝途徑中的關鍵酶和限速酶,可催化苯丙氨酸生成香豆酸、肉桂酸等中間產物,并且可以進一步轉化為綠原酸、香豆素,也能夠形成酯,再經歷多條途徑,進一步轉化為木質素、類黃酮等物質。研究發現:藥用植物在遭遇逆境脅迫時,PAL活性提高,當脅迫過嚴重時,PAL活性變弱[32],本研究也驗證了這一點。脅迫前期,PAL活性增強,催化苯丙氨酸代謝,促進黃酮代謝的進行;隨著脅迫時間加長,PAL活性降低。CHS和CHI分別是黃酮類化合物合成途徑中的第1個和第2個關鍵酶,脅迫會上調CHS、CHI活性[33]。本研究中,CHS和CHI活性在PAL之后增強,而黃酮質量分數也在這時達到峰值,這與酶作用途徑中的順序一致;而在脅迫后期,PAL、CHS、CHI活性不斷下降,三葉青黃酮質量分數也有所回落,這可能是由于脅迫使三葉青生長受到威脅,嚴重抑制各種生命活動,各種酶的活性也不斷下降。
郭肖等[34]對水芹Oenanthejavanica黃酮質量分數和PAL活性的相關性分析發現:黃酮含量與PAL活性的變化趨勢一致。本研究發現:三葉青塊根中PAL、CHS、CHI活性與黃酮質量分數呈正相關,且部分指標顯著相關(P<0.05),表明三葉青黃酮質量分數與關鍵酶活性相關,關鍵酶活性增強可能是引起三葉青黃酮高積累的關鍵因素。許多研究也證明了這一點,如LI等[35]研究發現:轉基因煙草Nicotianatabacum的總黃酮質量分數與CHI活性呈正相關;在溫度與水分脅迫下,參與銀杏Ginkgo biloba黃酮類代謝的PAL、CHS基因表達量的變化趨勢與類黃酮質量分數變化趨勢基本一致[36]。
水分脅迫影響三葉青的生長,表現為葉片塊根受損、葉綠體超微結構損傷、黃酮質量分數以及黃酮合成途徑相關酶活性變化等。三葉青通過產生一系列應激反應以應對水分脅迫,適度的水分脅迫可通過增強黃酮合成途徑中的關鍵酶活性以增強抗逆性,同時增加黃酮的產量,提高三葉青的藥材品質。
猜你喜歡
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
四川蠶業(2021年3期)2021-02-12 02:38:46
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
中成藥(2018年11期)2018-11-24 02:57:00
中成藥(2017年8期)2017-11-22 03:19:40
中成藥(2017年10期)2017-11-16 00:50:13
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中成藥(2017年4期)2017-05-17 06:09:50
汽車觀察(2016年3期)2016-02-28 13:16:26
