巴斯德畢赤酵母SUT2和ATG26響應甲醇代謝機制
2021-06-29 01:00:40王鵬程趙天宇戰春君白仲虎楊艷坤
東北農業大學學報 2021年5期
王鵬程,李 翔,趙天宇,戰春君,2,3,白仲虎,2,3,楊艷坤,2,3*
(1.江南大學糧食發酵工藝與技術國家工程實驗室,江蘇 無錫 214122;2.江南大學工業生物技術教育部重點實驗室,江蘇 無錫 214122;3.江南大學糖化學與生物技術教育部重點實驗室,江蘇 無錫 214122)
嗜甲醇巴斯德畢赤酵母通過甲醇利用途徑(Methanol utilization pathway,MUTpathway)代謝甲醇[1]。因其具有高效且嚴格調控的醇氧化酶1啟動子(PAOX1)、培養條件簡單、高密度發酵、蛋白翻譯后修飾系統等優良特性,現已成為使用最廣泛外源蛋白表達宿主之一[2]。其表達系統基于甲醇代謝通路,但涉及大量轉錄和信號因子,且具體分子機理尚不清楚[3]。為尋找甲醇代謝潛在信號因子,分析K.phaffii以甲醇為碳源及甘油為碳源轉錄組數據[4],發現被甘油抑制而被甲醇顯著誘導的基因PAS_chr1-4_0516,通過蛋白序列比對,發現其與釀 酒 酵 母(Saccharomyces cerevisiae,S.cerevisiae)SUT2同源。而S.cerevisiae中SUT2為SUT1(Sterol up take 1,固醇攝取基因1)同源基因,可在厭氧條件下協同SUT1參與調控固醇攝取[5-7],故畢赤酵母中響應甲醇誘導的SUT2基因同樣可能為固醇代謝潛在基因。
畢赤酵母細胞中麥角固醇是最主要固醇成分[8],也是過氧化物酶體膜重要組成[9]。過氧化物酶體是甲醇代謝主要細胞器[10-11]。因此,推測響應甲醇誘導的SUT2通過調控麥角固醇代謝,影響過氧化物酶體膜形成以及過氧化物酶體增生,進而參與甲醇代謝。
甲醇代謝時,過氧化物酶體量動態調節通過細胞器生物發生與自噬之間的平衡實現[11]。過氧化物酶體自噬時,自噬相關蛋白ATG26通過GRAM結構域(靶向過氧化物酶體自噬特異性膜突起)和PH結構域(具有脂質結合活性)與ATG30(Autopha?gy-related gene 30,自噬相關基因30)、ATG8(Au?tophagy-related gene 8,自噬相關基因8)、ATG11(Autophagy-related gene 11,自噬相關基因11)等蛋白協同作用,參與過氧化物酶體自噬,控制過氧化物酶體體積及總量[12]。此外,ATG26蛋白也具有甾醇葡萄糖基轉移酶活性,可將麥角固醇轉化為甾醇葡萄糖苷[13]。結合SUT2潛在功能,本文深入研究SUT2與ATG26對麥角固醇作用及參與甲醇代謝機理。
1 材料與方法
1.1 主要材料
菌株和質粒如表1所示。文中所需引物均由蘇州金唯智公司合成;吡啶、氫氧化鉀購自國藥集團化學試劑有限公司,正己烷購自上海麥克林生化科技有限公司,N,O-雙(三甲基硅基)三氟代乙酰胺購自北京索萊寶科技有限公司;氣相色譜質譜聯用儀GCMS-QP2020購自日本島津公司;CO2培養箱3111購自賽默飛;其他材料、設備參照文獻[14]。

表1 菌株和質粒Table 1 Strains and plasmids
1.2 質粒構建
1.2.1 sgRNA-ATG26敲除質粒
輸入ATG26基因序列至在線網站(http://chop?chop.cbu.uib.no/),選擇菌種Pichia pastoris,獲得推薦的sgRNA序列;開展脫靶效應分析,選擇特異性、評分較高且GC含量合適的sgRNA序列[15]。并根據sgRNA序列設計引物對sg-F/R,詳細序列如表2所示。將上下游引物通過寡鏈退火形成重組片段,采用Golden Gate assembly方法將其和BsaⅠ線性化的sgRNA-Cas9質粒組裝并轉化至大腸桿菌JM109感受態,對陽性轉化子抽提質粒并測序后獲得陽性克隆(此CRISPR-Cas9基因編輯系統由實驗室構建)。
1.2.2 pGAP-ATG26過表達質粒
利用表2所示引物對ATG26-F/R,以GS115野生型基因組為模板作PCR,得到ATG26基因片段。將pGAPZB質粒利用KpnⅠ、XhoⅠ雙酶切得到載體片段。對片段和載體采用Gibson assembly,并轉化至大腸桿菌JM109感受態,對陽性轉化子抽提質粒,利用引物對26-YZ-F/R作PCR并測序后獲得陽性克隆。
1.3 畢赤酵母感受態制作和轉化
畢赤酵母感受態制作和轉化(電擊法)參考Invitrogen官方手冊。
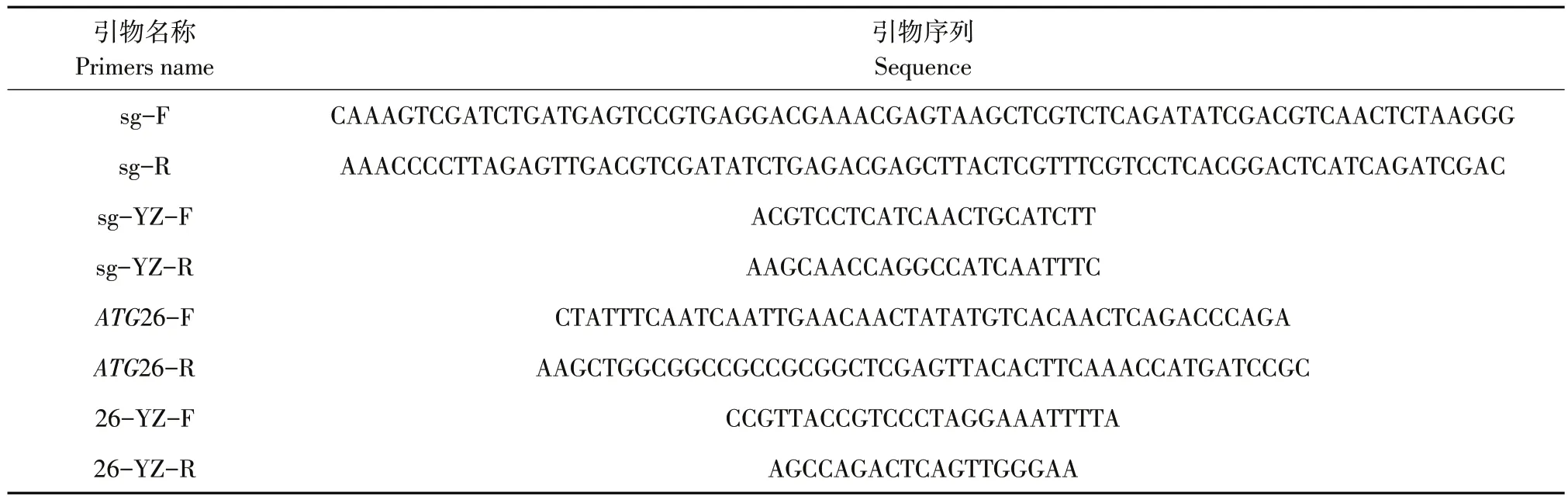
表2 研究所用引物Table 2 Primers in thisstudy
1.4 敲除菌株篩選及質粒丟失
將電轉后酵母感受態涂布于博來霉素平板上,30℃培養3~5 d至形成單菌落,挑取轉化子于博來霉素YPD培養基培養24 h后,提取酵母基因組DNA。利用表2所示引物對sg-YZ-F/R擴增目的片段并提交蘇州金唯智公司測序驗證。
將陽性轉化子接種至無抗性MM培養基,用以丟失sgRNA質粒。30℃、220 r·min-1震蕩培養24 h后于無抗性平板中劃線分離,30℃培養3~5 d后,挑取單菌落至博來霉素YPD培養基和無抗性YPD培養基。30℃、220 r·min-1震蕩培養24 h后,在無抗性YPD培養基中生長而抗性YPD培養基中無法生長的克隆,即為質粒丟失成功克隆,用于后續試驗。
1.5 主要培養基及菌株培養條件
1.5.1 主要培養基
YPD培養基:1%酵母提取物、2%Peptone、2%葡萄糖。
MM培養基:1.34%YNB、0.5%生物素、1%甲醇。
MG培養基:1.34%YNB、0.5%生物素、0.5%甘油。
酵母培養時,若需添加抗生素,博來霉素和氨芐青霉素終濃度均為100μg·mL-1。
1.5.2 菌株培養條件
將各菌株以1%接種量接種至5 mL YPD培養基,30℃、220 r·min-1震蕩培養,過夜活化。以OD600=0.5初始量分別轉接至50 mL MM(1%甲醇)、MG(0.5%甘油)培養基,并用PBS清洗菌體2次。30℃、220 r·min-1震蕩培養36 h時取樣并補加1%甲醇,培養60 h時再次取樣。
1.6 畢赤酵母胞內麥角固醇含量檢測
取2 mL新鮮菌液,于5 000 r·min-1離心5 min收集菌體。收集細胞用500μL 20%氫氧化鉀溶液(含50%乙醇)重懸,于沸水中煮沸10 min裂解細胞,室溫冷卻后加入500μL正己烷,渦旋震蕩萃取5 min后靜置分層,吸取上層正己烷相至新離心管;下層水相中加入250μL正己烷,重復萃取靜置分層后再次吸取上層正己烷相與第一次萃取液合并。將離心管置于80℃金屬浴揮發正己烷,揮發干凈后加入50μL吡啶和100μL N,O-雙(三甲基硅基)三氟代乙酰胺,80℃反應30 min后樣品制備完成[16]。
采用氣相色譜-質譜聯用儀分析鑒定樣品[17]。所用色譜儀為島津氣相色譜質譜聯用儀GCMSQP2020,色譜柱為DB-5MS(Agilent,30 m×0.25 mm×0.25μm),FID檢測器溫度300℃,載氣氦氣流速為1.2 mL·min-1;進樣口溫度300℃,分流進樣,進樣量1μL,分流比30∶1;柱箱程序升溫起始溫度80℃,然后以20℃·min-1速度升溫至280℃,保持15 min后以20℃·min-1速度升溫至300℃,保持5 min;質譜掃描范圍為40~450 m·z-1。
1.7 畢赤酵母AOX1酶活性檢測
畢赤酵母AOX1酶活性檢測方法參照文獻[14]。
2 結果與分析
2.1 p GAP-ATG26過表達菌株構建
將目的基因PCR產物與雙酶切后pGAPZB載體作Gibson assembly并轉化至JM109感受態。涂布于博來霉素平板,挑取陽性轉化子測序驗證。驗證正確后抽提質粒并單酶切線性化、電轉至GS115野生型菌株,博來霉素平板篩選。挑取陽性轉化子,提取基因組并PCR測序驗證,瓊脂糖凝膠電泳結果如圖1所示。理論長度為812 bp,圖中所示樣品11~13均符合。

圖1 ATG26過表達菌株PCR驗證Fig.1 Confirmation of ATG26 overexpression strainsby PCR
2.2 敲除菌株篩選
將構建成功sgRNA-ATG26敲除質粒電轉至GS115野生型菌株中,于博來霉素抗性平板中篩選陽性克隆,提取基因組DNA并PCR測序。比對結果如圖2所示,相比于野生型菌株,敲除菌株將95 bp處堿基A成功敲除。
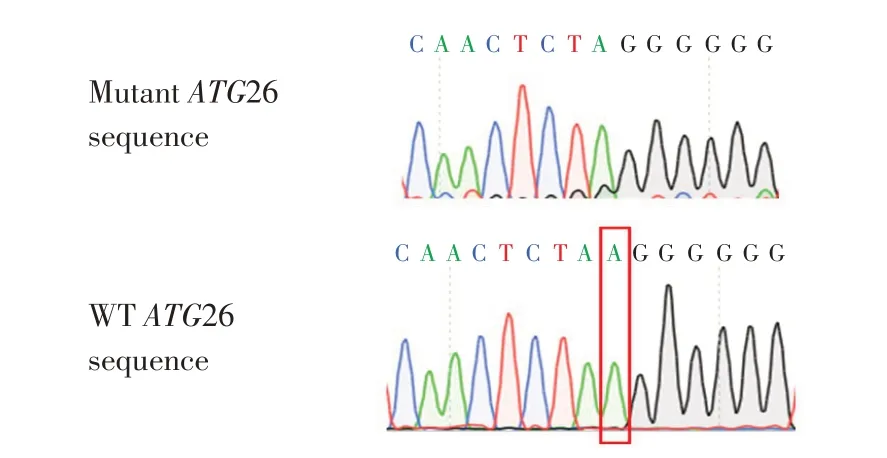
圖2 敲除菌株序列比對結果Fig.2 Sequence alignment of mutant strains and wild typestrains
2.3 SUT2高低表達對胞內麥角固醇及AOX1酶活性的影響
按照1.5中培養及取樣方法分別檢測野生型、SUT2敲除和過表達菌株在常氧及低氧條件下胞內麥角固醇含量及AOX1酶活性。其中,常氧下檢測結果如圖3所示。甲醇培養基中,相比于野生型菌株(WT),過表達SUT2提高約21%胞內麥角固醇含量,而敲除SUT2降低約10%~15%胞內麥角固醇含量(見圖3a)。因所用培養基為不含麥角固醇基本培養基,且胞外上清中均檢測不出麥角固醇(數據未顯示),故3株菌均無法從胞外攝取麥角固醇,也未分泌麥角固醇至胞外。因此本研究推測SUT2可能通過促進麥角固醇合成正向調節胞內麥角固醇含量。而甘油培養基中各菌株胞內麥角固醇含量無明顯變化(見圖3b),說明SUT2對麥角固醇合成的影響僅在甲醇誘導條件下存在,與SUT2在甲醇培養基中被誘導表達現象一致。
通過檢測AOX1酶活性發現,相比于野生型菌株,SUT2過表達菌株(SUT2-OE)的AOX1酶活性提高約27%(見圖3c),說明過表達SUT2對AOX1具有一定程度促進作用。因為,相比于甘油培養基,甲醇培養基中畢赤酵母過氧化物酶體顯著增生[18],而麥角固醇是過氧化物酶體膜重要組成[9],所以甲醇培養基中過氧化物酶體與麥角固醇含量呈正相關;過氧化物酶體又是甲醇代謝主要細胞器[10-11]。結合上述試驗結果,推測SUT2通過促進胞內麥角固醇合成并形成過氧化物酶體膜,促進過氧化物酶體增生,參與甲醇代謝并促進AOX1表達。
此外,SUT2敲除菌株(SUT2-KO)AOX1酶活性與野生型相近,但同樣有所提高(見圖3c)。可能因SUT2敲除后激活其他代謝通路導致。
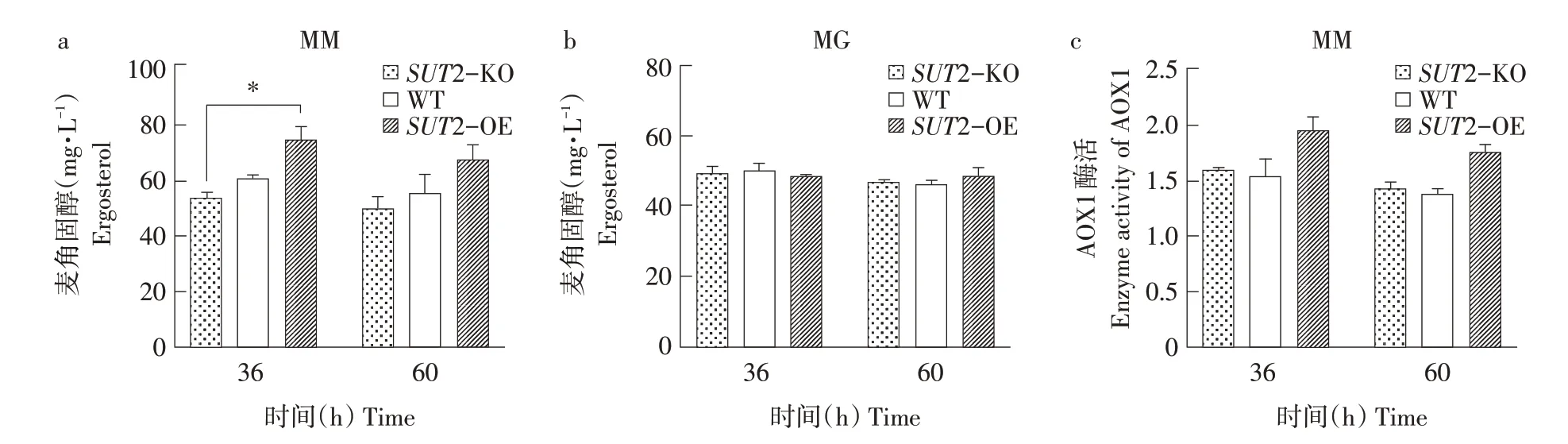
圖3 常氧條件下SUT2敲除、過表達及野生型菌株胞內麥角固醇含量及AOX1酶活性Fig.3 Intracellular ergosterol content and AOX1 enzymeactivity of SUT2 knockout,overexpression and wild typestrainsunder normoxia
因S.cerevisiae中SUT2為厭氧條件下與SUT1協同發揮作用基因[5-7],故本研究給予近似培養條件進一步研究,在1.5培養方法上作調整,實現低氧條件:靜置于低氧培養箱(CO2∶空氣=1∶4)中培養,同時將三角瓶瓶口用封口膜多次纏繞封閉。
低氧條件下檢測結果如圖4所示。相比于野生型,SUT2過表達對胞內麥角固醇含量無明顯影響,敲除SUT2降低約9%~13%胞內麥角固醇含量(見圖4a)。同時,SUT2過表達菌株AOX1酶活性具有一定程度提高,SUT2敲除菌株AOX1酶活性與野生型相似(見圖4b)。因為甲醇代謝為耗氧過程,低氧條件下甲醇代謝減弱[19],AOX1和SUT2蛋白活性減弱,導致對胞內麥角固醇合成以及AOX1酶活性促進作用均不明顯;若達到完全厭氧條件,改變SUT2表達量可能對胞內麥角固醇含量及AOX1酶活性均無明顯作用。
通過在不同氧含量、碳源條件下對畢赤酵母SUT2基因敲除、過表達研究,發現相比于S.cerevisiae,畢赤酵母SUT2基因具有不同功能:常氧條件下,甲醇培養基中畢赤酵母SUT2通過促進胞內麥角固醇合成,形成過氧化物酶體膜,促進過氧化物酶體增生,從而參與甲醇代謝。
2.4 ATG26高低表達對胞內麥角固醇及AOX1酶活性的影響
將構建成功的ATG26敲除、過表達菌株同樣按照1.5方法培養及取樣,結果如圖5所示。甘油培養基中各菌株胞內麥角固醇含量無明顯變化(見圖5b)。而甲醇培養基中,相比于野生型菌株(WT),ATG26過表達菌株(ATG26-OE)胞內麥角固醇含量下降約18%,AOX1酶活性下降約14%~20%;而ATG26敲除菌株(ATG26-KO)胞內麥角固醇含量提高約9%,AOX1酶活性提高約8%(見圖5a、c)。即ATG26對胞內麥角固醇含量以及AOX1酶活性均具有反向調控作用。又因甲醇培養基中過氧化物酶體與麥角固醇含量呈正相關,故ATG26通過改變胞內麥角固醇含量影響過氧化物酶體含量實現對AOX1酶活性反向調控作用。該調控作用源自ATG26具有的甾醇葡萄糖基轉移酶活性,可促進麥角固醇轉化為甾醇葡萄糖苷[11],減少過氧化物酶體膜形成,進而減少過氧化物酶體增生以及AOX1酶活性。此外,ATG26還可與ATG30、ATG8、ATG11等蛋白協同作用,參與過氧化物酶體自噬,調控過氧化物酶體總量及體積[10]。
2.5 巴斯德畢赤酵母SUT2與ATG26調節過氧化物酶體含量初步模型
基于上述試驗結果和已知調控通路,總結得到巴斯德畢赤酵母SUT2與ATG26調節過氧化物酶體含量初步模型,如圖6所示。
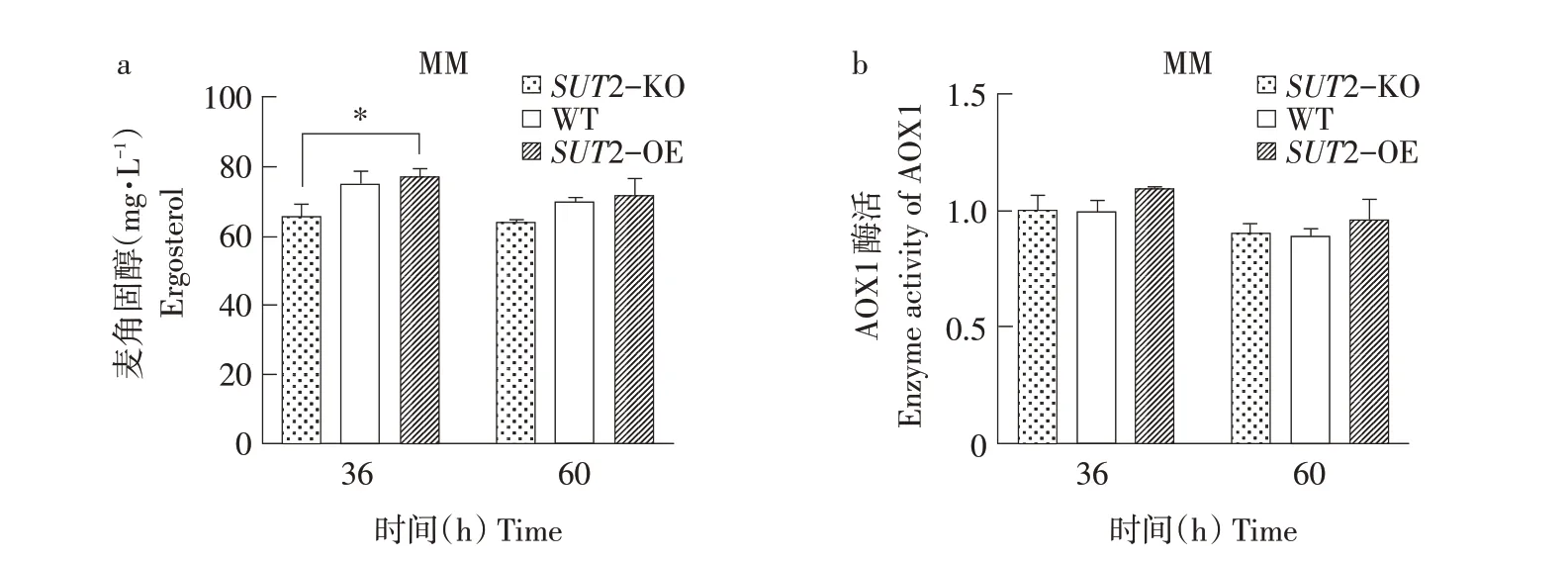
圖4 低氧條件下胞內麥角固醇含量及AOX1酶活性Fig.4 Intracellular ergosterol content and AOX1 enzymeactivity under hypoxia conditions
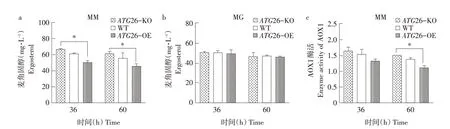
圖5 常氧條件下ATG26敲除、過表達及野生型菌株胞內麥角固醇含量及AOX1酶活性Fig.5 Intracellular ergosterol content and AOX1 enzymeactivity of ATG26 knockout,overexpression and wild type strains under normoxia
與釀酒酵母SUT2參與厭氧條件下胞外麥角固醇攝取不同,畢赤酵母SUT2受甲醇顯著誘導,雖然其具體誘導機制尚不清楚[4],但誘導表達的SUT2蛋白可能通過促進胞內麥角固醇合成并形成過氧化物酶體膜,進而促進過氧化物酶體增生并提高AOX1酶活,以此參與甲醇代謝。而同時存在ATG26蛋白一方面發揮甾醇葡萄糖基轉移酶活性促進麥角固醇轉化為甾醇葡萄糖苷,減少胞內麥角固醇含量,進而減少過氧化物酶體形成;另一方面作為自噬相關蛋白,與ATG30、ATG8、ATG11等協同作用,參與過氧化物酶體自噬。
因此,SUT2促進麥角固醇合成,ATG26將麥角固醇轉化為甾醇葡萄糖苷,兩者動態調節胞內麥角固醇含量及過氧化物酶體含量,進而調控甲醇代謝。
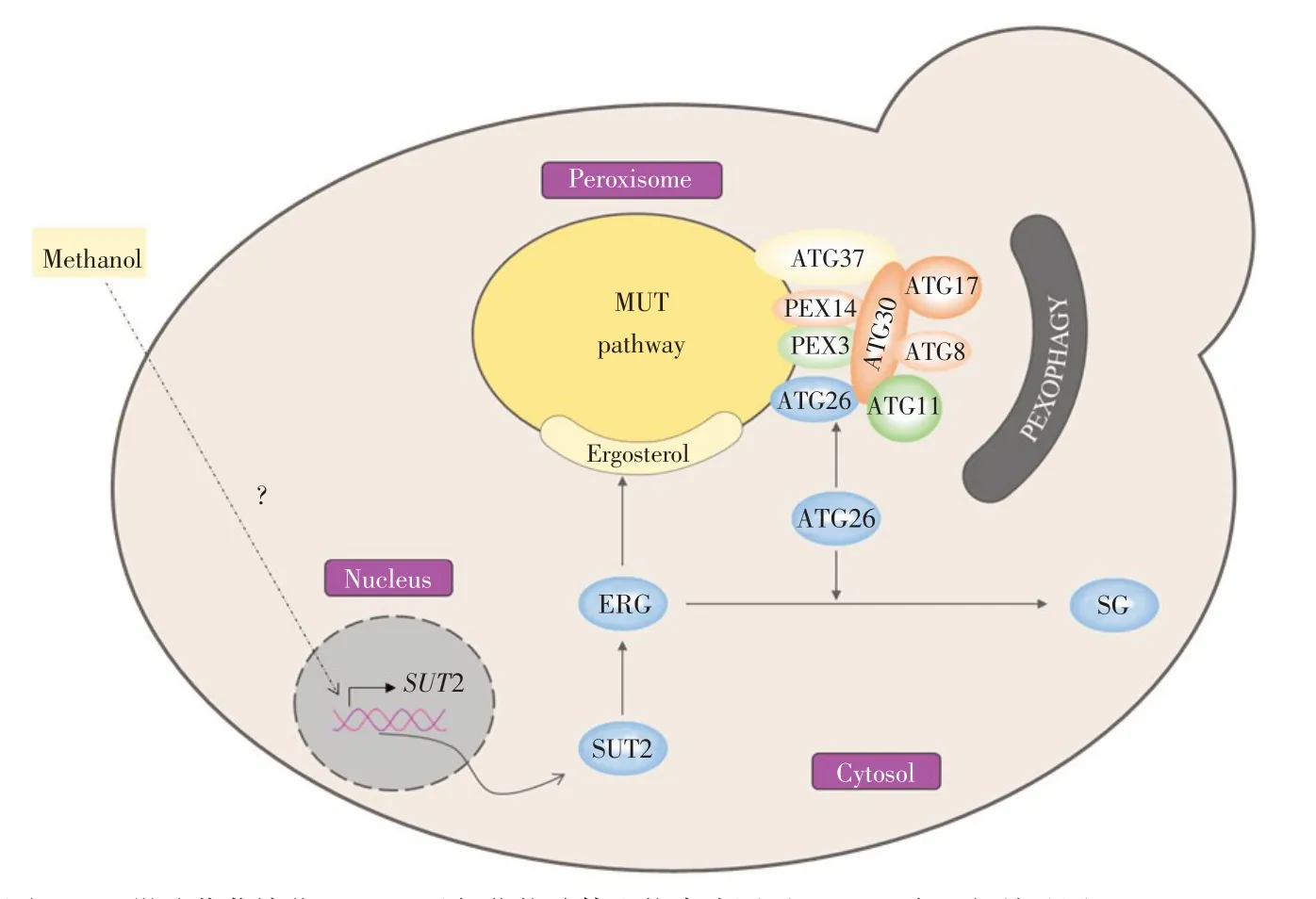
圖6 巴斯德畢赤酵母SUT2與ATG26調節過氧化物酶體含量模型Fig.6 Model of K.phaffii SUT2 and ATG26 regulating peroxisomecontent
3 討論與結論
受甲醇嚴格誘導的畢赤酵母PAOX1表達系統是目前應用最廣泛表達系統之一,該系統基于甲醇利用途徑,涉及較多潛在信號因子。本研究通過轉錄組數據分析發現,此前在K.phaffii中未被表征和深入研究的甲醇相關潛在因子SUT2,其具體功能機制尚不清楚。在S.cerevisiae中,SUT2響應于厭氧條件,協同SUT1參與固醇攝取[5-7];在植物細胞中,SUT2為糖轉運蛋白,參與蔗糖代謝[20]。本研究結合釀酒酵母SUT2功能,合理推斷畢赤酵母SUT2功能并驗證。最終發現畢赤酵母SUT2具有不同于釀酒酵母和植物細胞SUT2的全新功能,畢赤酵母SUT2響應于有氧條件下甲醇代謝,促進麥角固醇合成并形成過氧化物酶體膜,促進過氧化物酶體增生,參與甲醇代謝。
畢赤酵母胞內過氧化物酶體含量動態調節通過生物發生和自噬降解之間平衡實現[11],生物發生時PEX11(Peroxisomal protein 11,過氧化物酶體膜蛋白11)等蛋白從頭合成過氧化物酶體,胞內麥角固醇含量對過氧化物酶體增生具有重要作用。此前研究報道中,一般通過改變PEX11等過氧化物酶體膜蛋白表達量調節過氧化物酶體含量[21],尚未見利用胞內麥角固醇含量調節過氧化物酶體含量方法。本研究結合ATG26和SUT2全新功能,通過調節胞內麥角固醇含量,動態調節過氧化物酶體含量,調控甲醇代謝全新模型。該模型中,SUT2促進麥角固醇合成,進而促進過氧化物酶體增生和甲醇代謝;而ATG26將麥角固醇轉化為甾醇葡萄糖苷,減少胞內麥角固醇含量和過氧化物酶體增生,減弱甲醇代謝。該模型建立豐富K.phaffii甲醇代謝機理研究,有助于進一步完善畢赤酵母甲醇調控網絡。此外,過表達SUT2和敲除ATG26均有利于提高AOX1酶活;即通過改造該模型,有利于增強基于甲醇利用途徑的PAOX1表達系統,為工業上應用PAOX1表達系統提供一種新思路。
本研究SUT2受甲醇誘導具體機制尚不清楚,仍需后續進一步研究,發現新信號因子,進一步闡明K.phaffii甲醇代謝調控機理。
