哺乳動物外周組織中5-羥色胺功能的研究進展
2021-06-21 06:04:48閆曉茹張志飛高慧杰鄭惠玲
中國獸醫學報 2021年4期
杜 瑋,閆曉茹,張志飛,高慧杰,鄭惠玲
(西北農林科技大學 動物科技學院,陜西 楊凌 712100)
5-羥色胺(5-hydroxytryptamine,5-HT)又名血清素,是一種生物胺和色氨酸衍生物。5-HT可分為神經元5-HT和外周5-HT,之前的研究主要集中在神經元5-HT對情緒和認知的調控作用上,但是近年來的研究表明外周5-HT在機體的穩態調節過程中也發揮著非常重要的作用[1-2]。
人體中約98%的5-HT存在于外周,乳腺、腸道、骨骼、胰島等外周組織中均存在5-羥色胺能系統,都能進行5-HT生物合成、轉運和分泌。約90%的外周5-HT來源于腸道,被儲存在血小板中,經血液循環運達全身[3-4]。外周5-HT以自分泌和旁分泌的形式通過作用于不同組織和細胞表面的5-HT受體,進而調控機體代謝。現綜合近年來國內外關于外周5-HT功能的相關研究,旨在為調控哺乳動物乳腺穩態、維持骨骼密度和能量穩態的相關研究提供依據。
1 5-HT的生物合成及代謝
1.1 5-HT的合成5-HT是一種進化過程中高度保守的單胺。動物體中的L-色氨酸先經色氨酸羥化酶(tryptophan hydroxylase,TPH)轉化為5-羥色胺酸(5-hydroxytryptophan,5-HTP),然后經芳香族氨基酸脫羧酶(aromatic amino acid decarboxylase,AADC)脫羧形成5-HT[5]。TPH是5-HT生物合成過程中的限速酶,該酶在脊椎動物中存在兩種亞型,由不同的基因TPH1和TPH2轉錄而來[2]。TPH2在神經元中表達,神經系統中合成的5-HT起著中樞神經遞質的作用。TPH1在腸道、乳腺、胰島等多種外周組織中表達,外周組織中合成的5-HT作為一種激素,可以通過自分泌和旁分泌途徑影響止血、免疫反應、乳腺穩態、腸道生理和鈣代謝等多種機體功能[6-8]。
1.2 5-HT的轉運和代謝完整的5-羥色胺能系統包括了5-HT的合成、分泌、轉運和代謝功能。5-HT的轉運和代謝由5-羥色胺轉運體(serotonin transporter,SERT)和囊泡單胺轉運體(vesicular monoamine transporter,VMAT)來承擔。SERT具有兩大功能:第一是5-HT積累和再攝取,在不能合成5-HT的細胞如血小板中可以利用SERT聚集來自腸道合成的5-HT;SERT的第二個功能是清除細胞外分泌的5-HT,從而終止受體介導的5-HT信號傳導[9]。抗抑郁藥物5-羥色胺再攝取抑制劑(selective serotonin reuptake inhibitor,SSRIs)就是通過抑制SERT從而延長5-HT在細胞外空間(尤其是突觸)的停留時間來增加5-HT的生物活性。5-HT通過VMAT進入分泌囊泡,并被單胺氧化酶(MAO)降解,產生主要的非活性代謝產物5-羥基吲哚乙酸(5-HIAA),最終隨尿液排出體外[10]。
2 5-HT受體
5-HT通過其受體發揮作用,5-HT的受體可以分為7個家族(5-HT1~7),這7個家族又分為14個亞型:5-HTR1A、1B、1D、1E、1F、2A、2B、2C、3A、3C、4、5a、6和7[11]。5-HT受體具有組織和細胞差異性,同一家族的不同受體亞型,其結構、相對分布、對各種激動劑和拮抗劑的敏感性也不相同。除5-HT3家族是配體-門控離子通道外,其他5-HT受體都是G蛋白偶聯受體。其中5-HT1和5-HT5家族的受體是Gi/o耦合的,5-HT2家族是Gq/11耦合,5-HT4、5-HT6和5-HT7是Gs耦合的[12-13]。此外一些5-HT受體基因,如5-HTR2C和5-HTR7,在轉錄過程中可以通過可變剪接產生多種變體,這進一步增加5-HT受體的多樣性。外周5-HT通過其受體發揮作用,5-HT受體的多樣性決定了一個器官乃至整個機體中5-HT廣泛的功能。
3 外周組織中5-HT的功能
3.1 乳腺在穩態條件下,乳腺泌乳是一個周期性的進程,在這個過程中乳腺導管被交替充滿和排空。在乳腺中,通過神經內分泌途徑傳遞的信號和腺體局部信號之間的復雜相互作用,可實現乳腺泌乳穩態[14]。催乳素(PRL)和催產素(OT)通過作用于分泌上皮細胞和肌上皮細胞,引起乳汁的合成、分泌和排出。這種神經內分泌反饋回路是調控泌乳期內穩態的主要外在機制[15]。與神經內分泌系統的調控相比,乳腺內固有的穩態調控機制在很大程度上仍是未知的。近年來的研究表明5-HT是一種乳腺穩態的調節因子。WEAVER 等[16]通過哺乳期乳腺特異性敲除TPH1的小鼠模型,發現乳腺是哺乳期循環5-HT產生的主要來源。乳腺5-HT已被證明具有兩種穩態功能。首先,5-HT調節泌乳,并啟動乳腺進入退化早期階段。第二,5-HT是一種局部信號,可誘導甲狀旁腺激素相關蛋白(PTHrP)表達,動員骨骼中的鈣釋放進入血液并轉移到乳腺[9]。
3.1.1外周5-HT調節乳腺穩態 5-HT被認為是嚙齒動物、反芻動物和人乳腺泌乳的自分泌/旁分泌調節因子。MATSUDA 等[17]在嚙齒動物模型上的研究表明,PRL刺激小鼠乳腺組織中5-HT生物合成必需基因:TPH1和AADC的表達。孕期和哺乳期TPH1 mRNA表達升高,表明5-HT的生物合成增加。哺乳期母鼠的乳汁淤積可誘導體外培養的乳腺球細胞TPH1的表達,提示該基因是由腺泡中的乳汁充盈所控制的。5-HT抑制乳腺上皮細胞中β-酪蛋白基因表達,引起乳腺小泡萎縮[18]。相反,TPH1基因突變或抗5-HT藥物可以導致分泌功能增強和腺泡擴張[17]。因此5-HT可以抑制乳腺乳汁合成,維持乳腺容積穩態。上皮緊密連接的建立和維持是泌乳所必需的生理事件之一。在分娩后不久,乳腺上皮細胞緊密連接的完整功能就建立起來了,并且在整個哺乳期它們都是閉合的。相反,緊密連接的開放被認為是乳腺退化的早期事件之一[19]。STULL等[15]發現5-HT通過位于基底外側膜上的5-HTR7介導了緊密連接的開放從而使乳腺退化。因此,自分泌和旁分泌5-HT信號是調節乳腺內穩態和介導乳腺退化的重要因子。
在反芻動物中的研究表明,TPH1在牛的乳腺上皮細胞中表達且TPH1以PRL依賴的方式表達。在牛乳腺上皮細胞中鑒定出5種5-HT 受體亞型,包括5-HTR1B、5-HTR2A、5-HTR2B、5-HTR4和5-HTR7[20]。5-HTR1B亞型的突變已經被證明可以增加牛的產奶量[21]。PAI等[22]研究表明,5-HTR7受體敲除的母牛乳腺畸形,無法哺乳幼崽,難以從哺乳期過渡到干奶期。因此推測5-HT可以通過5-HTR1B和5-HTR7亞型受體調節奶牛的泌乳和乳腺的發育。人體的藥理學研究表明服用SSRIs的產婦會出現乳腺分泌激活的延遲導致母乳喂養困難[23]。綜上所述,在人、嚙齒動物、反芻動物中5-HT作為泌乳的抑制劑,與PRL形成一個負反饋調節環路,兩者共同調節哺乳期的乳腺穩態。
3.1.2外周5-HT調節乳腺鈣代謝 進入哺乳期后,乳汁的合成迅速消耗母體循環中的鈣,因此,必須從母體骨骼中動員鈣以保持足夠的循環鈣濃度。循環鈣濃度受包括維生素D、甲狀旁腺激素(parathyroid hormone,PTH)、降鈣素和甲狀旁腺激素相關蛋白(parathyroid hormone related protein,PTHrP)在內的多種激素和代謝產物的精密調控[24]。哺乳期5-HT缺乏會導致小鼠乳腺的鈣感應、轉運和分泌功能受損,此外5-HT缺乏小鼠顯示出較低的循環和乳腺PTHrP濃度,其股骨的骨細胞數量減少,表明其在哺乳期間動員骨鈣的能力受損[25-26]。哺乳期5-HT通過5-HTR2B誘導乳腺上皮細胞中PTHrP的合成,從而引起骨動員增加循環鈣濃度。因此5-HT將泌乳與全身鈣代謝結合起來。圖1以一種簡化的方式說明了乳汁分泌驅動PTHrP的反饋回路。
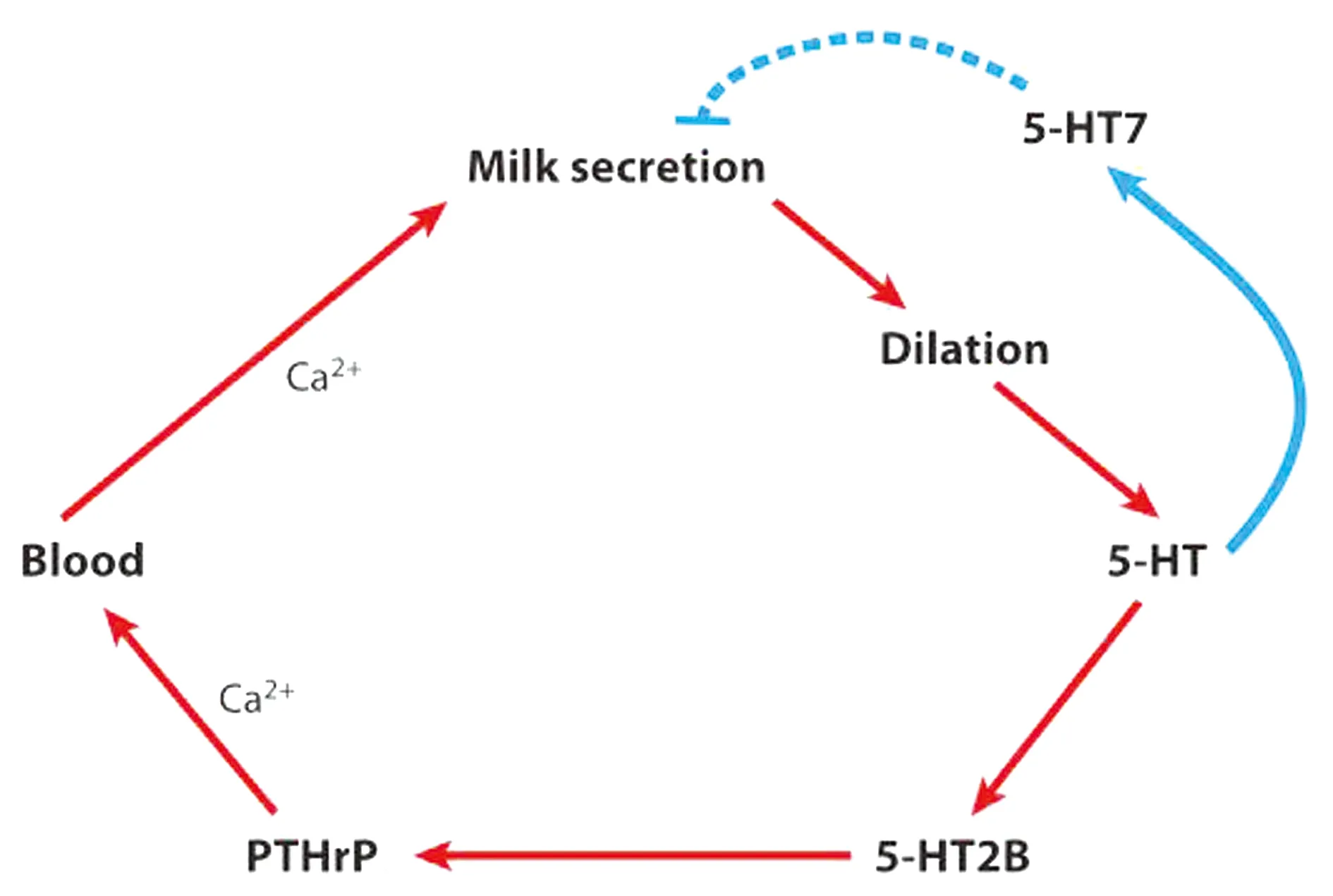
圖1 5-HT調節乳腺泌乳[9]
3.2 骨骼骨骼受多種內在和外在信號因子共同調節其生長和轉化,是一個高度動態的組織。骨骼系統通過成骨細胞(osteoblasts,OB)和破骨細胞(osteoclast,OC)的相互作用來維持其結構和功能,OB負責骨形成而OC負責骨吸收和鈣動員[27]。多種激素、生長因子和細胞因子協同調控成骨細胞和破骨細胞的活性,共同維持骨密度[28]。近年來的研究表明5-HT也是骨組織動力學中的一個關鍵分子。臨床上的數據也表明服用SSRIs治療抑郁癥的患者,他們發生骨折和骨質疏松的概率增加[29]。5-HT通過直接和間接的方式調控骨骼代謝。骨組織中除了含有腸源性5-HT外,自身的OB、OC、骨細胞都可以合成5-HT。
3.2.1腸源性5-HT對骨代謝的調控 腸源性5-HT是由L-色氨酸通過腸嗜鉻細胞中TPH1合成的,合成后的5-HT進入血液在表達有SERT的血小板中聚集。腸源性5-HT是骨骼中5-HT的主要來源[30]。低密度脂蛋白受體相關蛋白5(LRP5)與骨形成的變化相關,LRP5缺失突變會導致骨質疏松,而功能獲得性突變會導致高骨量綜合征[31]。LRP5-/-小鼠十二指腸中的TPH1表達升高,血液中5-HT含量增加,骨密度降低。因此LRP5可通過調控TPH1的表達影響5-HT含量,從而調控骨形成。YADAV等[30]進一步的研究表明,5-HT通過前成骨細胞表面的5-HTR1B受體抑制成骨細胞的增殖(圖2)。在正常生理條件下,叉頭框蛋白O1(FOXO1)在成骨細胞細胞核內與轉錄激活子4(activating transcription factor 4,ATF4)和cAMP反應元件結合蛋白(cAMP response element-binding protein,CREB)相互作用,維持成骨細胞的正常增殖。FOXO1與ATF4的結合促進了FOXO1調節轉錄靶點的表達,而與CREB的結合抑制了FOXO1調節轉錄靶點的表達[32]。血液循環中5-HT水平增加破壞了FOXO1與CREB的相互作用,有利于ATF4-FOXO1異二聚體的形成,這導致細胞周期進程基因被抑制,成骨細胞增殖活性下降,骨形成被抑制[32]。
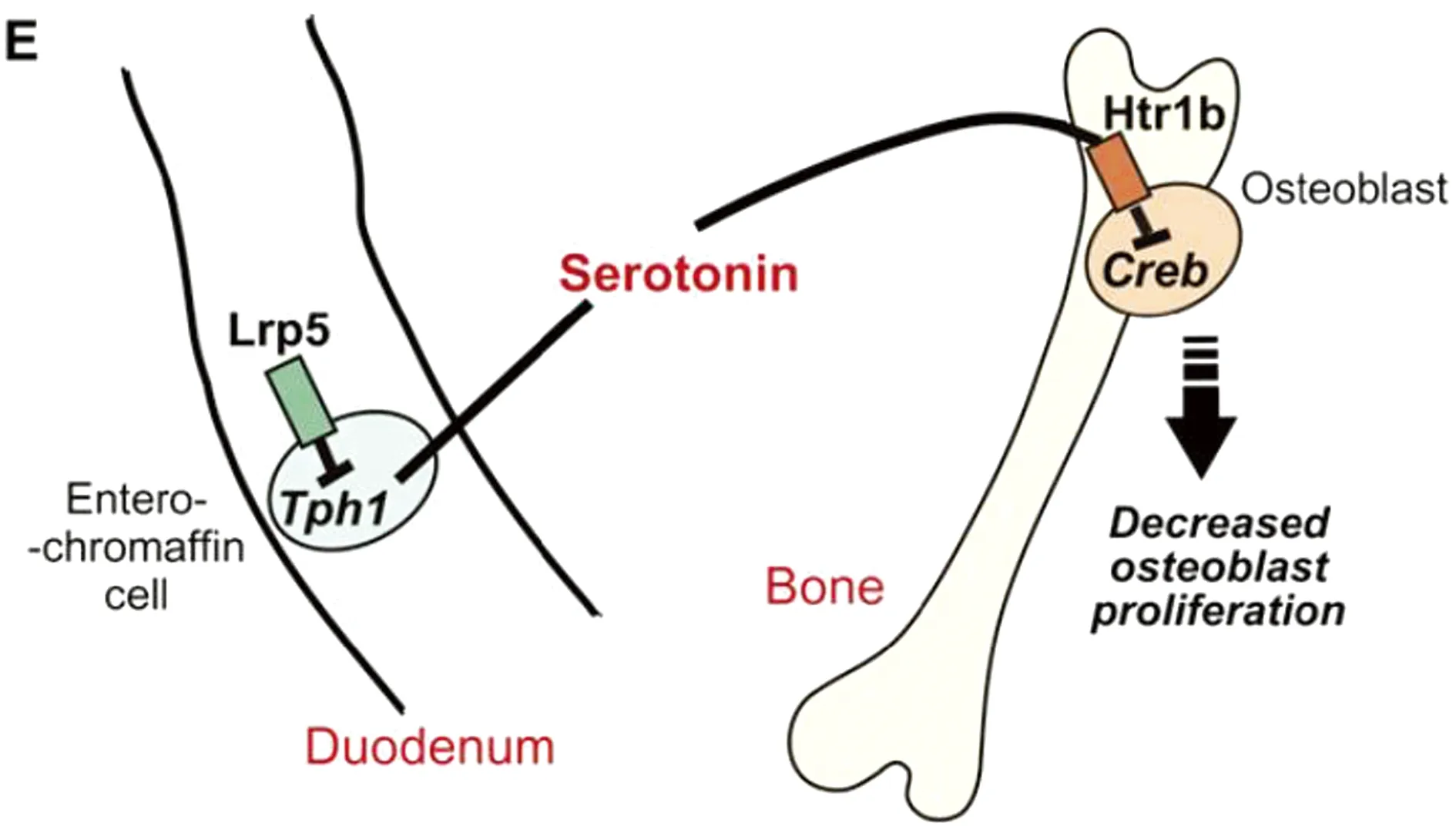
圖2 腸源性5-HT調節成骨細胞增殖[30]
3.2.2骨源性5-HT對骨代謝的調控 近年來的研究表明,成骨細胞、破骨細胞和骨細胞中均表達TPH1、SERT、5-HTRs,這說明骨骼中存在5-羥色胺能系統。骨細胞中產生的5-HT能夠以自分泌和旁分泌的形式作用于細胞膜表面的5-HT受體進而調節骨細胞的增殖和活性。骨源5-HT可以通過5-HTR2B促進成骨細胞增殖而通過5-HTR1B抑制成骨細胞增殖[33]。5-HT可以由破骨細胞前體在NF-κB配體(RANKL)存在的情況下合成,破骨細胞前體內TPH1表達的下調會導致骨吸收減少,從而增加骨密度[33]。CHABBI等[34]使用TPH1持續失活的小鼠,發現破骨細胞可以局部產生5-HT,其通過增加成骨細胞的活性同時減少破骨細胞的產生促進骨形成。
3.2.3PTHrP對骨代謝的調控 骨骼接受來自乳腺釋放的信號因子調節骨代謝,乳腺上皮細胞中產生的5-HT通過位于細胞膜表面的5-HTR2B受體促進細胞內PTHrP的表達。乳腺產生的PTHrP進入血液循環中,在骨組織中與位于OB表面的PTHrP受體(PTH1R)結合,誘導OB分泌RANK配體(RANKL)。RANKL與OC表面的RANK受體結合促進骨吸收,釋放鈣離子進入血液循環中[35]。嚙齒動物和反芻動物上的研究表明,產前5-HT/5-HTP治療可以維持母體產后血液中的鈣濃度,降低了產后低血鈣癥的發生率,使母體由圍產期順利過渡到哺乳期[36-37]。
3.3 肝臟肝臟是人體重要的脂質代謝器官,脂肪酸、甘油三酯、膽固醇的合成分解以及膽汁分泌等脂類代謝活動都是在肝臟中進行的。當體內攝入過多能量時,肝臟中出現異常的脂質聚集從而引發非酒精性脂肪性肝病(nonalcoholic fatty liver disease,NAFLD),這是一種常見的肝臟疾病[38]。此外肝臟也能夠通過糖異生的方式參與葡萄糖代謝,調節體內的血糖水平。糖異生的增加會導致血糖水平升高,導致2型糖尿病(type 2 diabetes mellitus,T2DM)的發生。
3.3.1外周5-HT調節肝臟脂代謝 許多研究表明外周5-HT能夠調節肝臟中的脂質代謝。CHOI等[39]發現TPH1和TPH2在8周齡C57BL/6J小鼠肝臟中不表達,但檢測到了SLC6A4(編碼SERT)、MAOA(單胺氧化酶A)和5-HTRs基因的表達,這表明肝細胞可以轉運和代謝5-HT但不能合成5-HT。此外肝臟是腸源性5-HT經門靜脈第一個遇到的器官,這時門靜脈血中游離5-HT的濃度較高,足以對肝臟的內分泌產生影響[40]。研究表明腸道特異性TPH1敲除(TPH1-GKO)小鼠可以抑制高脂飼喂引起的肝臟脂肪變性,因此腸源性5-HT的產生與NAFDL的發生有關。此外肝臟特異性敲除5-HTR2A(5-HTR2A-LKO)小鼠以及選擇性5-HTR2A拮抗劑治療的小鼠與野生型相比肝臟中脂肪生成相關基因的表達減少,抑制了高脂飼喂引起的肝臟脂肪變性[39]。因此,腸源性5-HT是由腸-肝內分泌軸通過5-HTR2A直接調節肝臟脂質代謝。
3.3.2外周5-HT調節肝臟糖異生 早在1992年就有研究表明,外周5-HT可以促進肝臟中的糖異生。ZABALA等[41]通過給大鼠注射5-HT,上調了肝臟中糖異生過程中關鍵酶磷酸烯醇式丙酮酸羧化酶的表達。SUMARA等[42]進一步的研究表明在肝細胞中腸源性5-HT信號可以通過增加果糖1,6-二磷酸酶(FBPase)和葡萄糖-6-磷酸酶(G6Pase)這兩種糖異生限速酶的活性,從而促進了糖異生。在特異性敲除5-HTR2B小鼠肝細胞中,糖異生的活性受到抑制,因此5-HT可能通過5-HTR2B參與糖異生的調控。此外,肝細胞內的腸源性5-HT信號傳導以葡萄糖轉運蛋白2(Glut2)依賴的方式抑制葡萄糖攝取,從而進一步有利于維持血糖水平。因此,抑制腸源性5-HT的合成可以改善高脂飲食引起的葡萄糖不耐受。使用抑制腸源性5-HT合成的小分子制劑或者使用選擇性5-HTR2B拮抗藥物可能有助于改善T2DM。
3.4 胰島胰島素是由胰腺β細胞分泌的,受血糖水平調控,進食后血糖水平升高可使胰島素分泌增加5~10倍[43]。除血糖外,血脂、細胞因子和激素都能影響胰島素分泌。T2DM是一種以胰島素分泌缺陷和胰島素敏感為特征的代謝性疾病[44]。
研究表明包括胰腺β細胞在內的一些胰島細胞中表達有合成、包裝和分泌5-HT所必需的所有基因。β細胞表達TPH的兩種亞型TPH1和TPH2,β細胞也表達VMAT2,它的作用是將5-HT裝入分泌囊泡的轉運體中[45]。ZHANG等[46]發現T2DM患者血漿中5-HT水平較高,且5-HTR2C在糖尿病小鼠胰島中的表達明顯增加。5-HTR2C激動劑mCPP對Min-6細胞和胰島細胞的胰島素分泌有明顯的抑制作用,5-HTR2C拮抗劑SB242084或RNA干擾5HTR2C可逆轉這種抑制作用;提示胰腺β細胞中5-HTR2C表達增加可能抑制胰島素分泌。
3.4.1外周5-HT參與妊娠適應 妊娠期為滿足代謝需求,母體胰島β細胞的數量和葡萄糖刺激胰島素分泌(glucose-stimulated insulin secretion,GSIS)均增加[47]。妊娠期間在PRL的刺激下,胰島中TPH1、TPH2的兩種亞型表達水平顯著升高[48]。胰島產生的5-HT以自分泌/旁分泌方式發揮作用,在妊娠期間通過5-HT配體-門控離子通道受體(5-HTR3)增加GSIS[48]。5-HT可以引起的通道開放激活了內向電流并使膜去極化。葡萄糖也能使β細胞去極化:糖酵解增加的ATP導致ATP敏感的K+通道關閉,由此產生的去極化通過電壓門控的Ca2+通道引起Ca2+內流,從而觸發胰島素顆粒胞吐[49]。因此,通過5-HTR3激活去極化使β細胞對葡萄糖刺激更敏感。在妊娠中期胰島5-HT通過Gαq-偶聯受體5HTR2B誘導β細胞增殖、增加β細胞質量,在妊娠末期通過Gαi-偶聯受體5HTR1D降低β細胞質量[50]。總之5-HT參與了β細胞對妊娠的適應,這些適應性過程的失敗會導致妊娠期糖尿病的發生。
3.4.25-HT調控胰島素分泌 肥胖會導致胰島素抵抗,從而增加身體對胰島素的需求。隨著胰島素需求的增加,胰腺β細胞質量和胰島素分泌增加[51]。當β細胞不能補償胰島素抵抗時,就會產生T2DM[52]。KIM等[53]研究了在高脂飲食誘導的胰島素抵抗狀態下5-HT在胰島素分泌中的作用。與妊娠期胰島5-HT合成顯著增加不同的是HFD并未提高胰島內5-HT合成或血液5-HT水平。在HFD誘導的胰島素抵抗狀態下5-HTR3敲除(5-HTR3A-KO)小鼠和TPH1β細胞特異性敲除(TPH1β-KO)小鼠均表現出葡萄糖不耐受。5-HT處理可改善TPH1β-KO胰島的胰島素分泌,但不能改善5-HTR3A-KO胰島的胰島素分泌。這進一步證實了5-HTR3在5-HT調控GSIS過程中的關鍵作用,5-HTR3有望成為治療T2DM的潛在靶點。
4 總結與展望
動物體內的多種外周組織都能進行5-HT的生物合成、轉運和分泌。外周5-HT作為一種激素通過自分泌或旁分泌的形式作用于不同的受體,參與了機體穩態的調控。在畜牧業生產中,機體代謝紊亂會損害動物的生產性能,降低養殖業的經濟效益。總結和分析外周5-HT在乳腺、骨骼、肝臟和胰島中的作用(圖3),旨在為調控哺乳動物機體穩態提供依據。隨著對外周5-HT及其受體功能研究的深入,5-羥色胺能制劑將成為防治泌乳紊亂、低血鈣癥、骨質疏松、能量代謝失衡等疾病的有效藥物。

圖3 不同外周組織中5-HT的功能
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年2期)2019-08-23 08:12:10
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
中國醫藥科學(2015年15期)2015-02-27 12:32:27
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國全科醫學(2013年36期)2013-01-25 06:20:58
