三門灣海域漁業生物群落結構及營養級變化特征
2021-06-21 09:02:54俞松立王詠雪韓曉鳳章翊涵來洪運章凱張麗源水柏年
大連海洋大學學報 2021年3期
俞松立,王詠雪,韓曉鳳,章翊涵,來洪運,章凱,張麗源,水柏年*
(1.浙江海洋大學 水產學院,浙江 舟山 316022; 2.臺州市海洋環境監測中心,浙江 臺州 318000)
海洋漁業生物是指海域中具開發利用價值的魚類、甲殼類、頭足類等,是海洋生態系統中重要的生態類群之一,對其開展洄游、分布、數量等要素的調查與研究,可為漁業資源的保護、修復和管理等措施與政策的制定提供依據。關于漁業生物群落特征的研究國內對各海域已有較多報道,張煥君等[1]利用聚類分析、非度量多位標度分析等方法對萊州灣的游泳動物群落特征進行了研究;劉鴻雁等[2]利用相對重要性指數和ABC曲線等方法對青島嶗山灣人工漁礁底層游泳動物群落結構進行了研究;姜亞洲等[3]采用物種多樣性指數和多元分析等方法對象山港游泳動物群落結構進行了研究;胡成業等[4]結合物種多樣性、ABC曲線、群落聚類及非度量多維測度等方法對溫州甌飛灘鄰近海域春季游泳動物群落結構及多樣性進行了研究;張洪亮等[5]運用物種多樣性指數與環境因子相結合的方法分析了浙江南部近海魚類的多樣性;李淵等[6]利用相對重要性指數、物種多樣性等對南沙群島西南部和北部灣口海域的游泳動物多樣性進行了研究。然而,關于三門灣海域環境、資源調查研究僅有少量報道,如蔡萌等[7]對三門灣海域魚類的種類組成進行了研究,聶振林等[8]對該海域魚類群落與水溫、鹽度、pH、溶解氧、化學需氧量和懸浮物等環境因子的關系展開了分析,冀萌萌等[9]對該海域游泳動物的群落組成和多樣性進行了研究。綜上,目前對三門灣海域的調查存在分析方法、時間和指標等不全面的問題,不能全面反映該海域漁業生物群落特征。
三門灣是浙江省的三大半封閉海灣之一,水系發達、餌料豐富,是海洋生物生長、繁衍的良好場所,也是國家海洋局“十三五”規劃中需要生態修復的海灣之一。近年來,受沿岸工農業發展、海底管線工程、核電站建設及高強度捕撈生產影響,三門灣海域海洋生態環境形勢較為嚴峻,漁業資源出現加速衰退[10-11]。為了更深入全面地對該海域漁業資源進行相關研究,需采用多種方法,如相對重要性指數、資源密度、物種多樣性、ABC曲線、聚類分析、非度量多維標度分析、相似性百分比、生態位寬度和營養級結構等方法,同時從多項指標對三門灣海域漁業生物群落特征變化進行綜合分析。本研究中,基于2018年春、夏、秋、冬 4季三門灣海域漁業資源的調查,對該海域漁業生物群落的種類組成、結構及營養級等進行了全面研究,以期為漁業資源增殖放流和海洋生態修復策略提供科學依據。
1 材料與方法
1.1 調查海域及時間
依據《海洋調查規范》[12]、《海洋漁業資源調查規范》[13],于2018年的春季(4月)、夏季(7月)、秋季(10月)和冬季(12月)對三門灣海域進行底拖網漁業資源調查,共設20個站位(圖1)。在拖網過程中,網口張開寬度約14 m,網囊網目尺寸為20 mm,每網拖曳時間為0.5 h,平均拖速為5.56 km/h,對所得漁獲物進行低溫保存,運回實驗室后進行鑒定、分類。
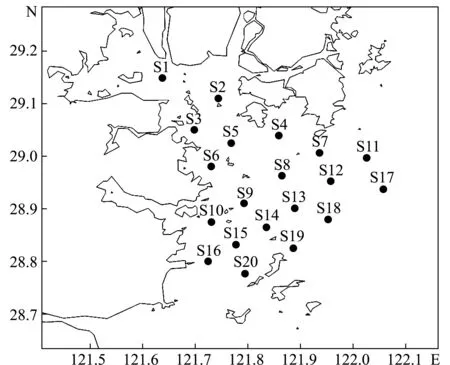
圖1 三門灣海域漁業資源調查站位
1.2 相關指數的計算
1.2.1 相對重要性指數 利用相對重要性指數(IRI)[14]對該海域中的優勢種、重要種進行劃分,計算公式為
IRI=(N+W)×F×10 000。
(1)
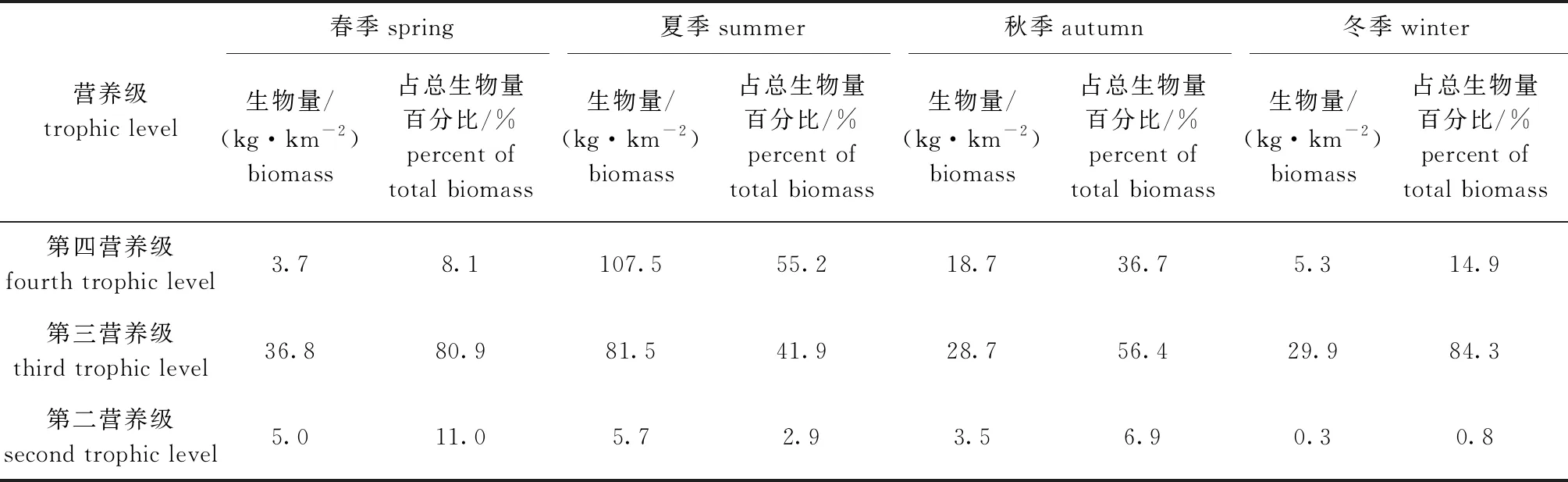
其中:N為物種個體數占總個體數的百分比;W為物種質量占總質量的百分比;F為出現頻率,即某種類出現的站位數與總站位數的百分比。參照王雪輝等[15]的研究方法,IRI>1 000的種類為優勢種,100 1.2.2 相對資源密度 漁業生物的資源密度估算采用掃海面積法[16],其計算公式為 Di=Ci/(Aiq)。 (2) 其中:Di為第i站位的漁業資源密度(數量密度,103ind./km2;質量密度,kg/km2);Ci為第i站位的每小時拖網漁獲量(數量,103ind./h;質量,kg/h);Ai為第i站位單位時間拖網的掃海面積(km2/h),即網口水平擴張寬度(km)、拖網速度(km/h)和實際拖網時間(h)的乘積;q為可捕系數,本研究中取0.5。 1.2.3 物種多樣性指數 物種多樣性指數包括Shannon-Wiener多樣性指數(H′)[17]、Pielou均勻度指數(J)[18]和Margalef豐富度指數(D)[19]。由于不同種類及同種類的不同個體間差異較大,因此,Wilhm[20]、沈國英等[21]提出根據質量更適合計算漁業生物的物種多樣性,因此,本研究中根據質量對物種多樣性進行計算,計算公式為 (3) J=H′/log2S, (4) D=(S-1)/log2N。 (5) 其中:S為漁業生物的物種數;N為總質量(g);Pi為第i種漁業生物質量占總質量的比值。根據Khalaf等[22]的方法進行判斷:當H′>3時,表明漁業生物群落基本不受干擾;當1≤H′≤3時,表明群落受到中度的干擾;當0 1.2.4 ABC曲線 該方法是根據豐度與生物量優勢度曲線的相對位置情況來判斷群落受干擾程度的方法,兩條曲線與坐標軸圍成的面積為A,其計算公式[23]為 (6) 其中:ai與bi分別為物種i的豐度累積百分比與生物量累積百分比;S為總種類數。 當生物量優勢度曲線整條在豐度優勢度曲線上方時,即A>0,表明生物群落未受擾動;當生物量優勢度曲線與豐度優勢度曲線出現相交或重合時,即A值趨于0,表明群落受到中度干擾;當豐度優勢度曲線在生物量優勢度曲線上方時,即A<0,表明群落受到嚴重干擾。 1.2.5 組平均聚類和非度量多維標度分析 漁業生物的群落結構分析采用組平均聚類法和非度量多維標度分析法。其中,根據脅迫系數(stress)能對非度量多維標度分析的可信度進行評估[24],當stress≥0.2時,排序結果不能正確解釋群落結構關系;當0.1≤stress<0.2時,排序結果用排序圖表示具有一定解釋意義;當0.05≤stress<0.1時,結果基本可信;當stress<0.05時,結果有較好代表性。 1.2.6 生態位 生態位寬度采用Shannon指數[17],其計算公式為 (7) 其中:Bi為生態位寬度,取值范圍為[0,R];R為總站位數;Pij為物種i在j個站位中的個體數占總個體數的比例。Bi值越大,生態位寬度就越大。 1.2.7 營養級結構 通過查詢Fishbase網站及相關文獻[25-29]得出三門灣海域漁業生物的營養級,并對其進行劃分,劃分標準參照鄧景耀等[30]的方法,第一營養級為自養級,第二營養級為草食性種類和雜食性種類,第三營養級為低級肉食性種類和中級肉食性種類,第四營養級為高級肉食性種類。本研究中,將各營養級中漁業生物的個體數量和生物量進行匯總,以便對三門灣海域漁業生物群落的營養級結構進行分析。 調查中春季共鑒定出漁業生物66種,隸屬于16目39科55屬,其中,甲殼類數量和質量所占百分比最大,分別為57.9%和59.3%;夏季共鑒定出生物84種,隸屬于16目41科64屬,其中,魚類數量和質量所占百分比最大,分別為75.3%和79.4%;秋季共鑒定出生物69種,隸屬于14目31科53屬,其中魚類數量和質量所占百分比最大,分別為46.2%和58.4%;冬季共鑒定出生物42種,隸屬于13目22科34屬,其中,甲殼類數量和質量所占百分比最大,分別為57.9%和59.3%。 從表1可見:春季漁業生物優勢種為三疣梭子蟹、孔蝦虎魚和日本蟳,重要種為細螯蝦、矛尾蝦虎魚等11種;夏季優勢種為小黃魚、銀鯧、龍頭魚、帶魚、三疣梭子蟹、哈氏仿對蝦,重要種為中國毛蝦、白姑魚等12種;秋季優勢種為三疣梭子蟹、龍頭魚、尖頭黃鰭牙魚或、口蝦蛄、脊尾白蝦和棘頭梅童魚,重要種為海鰻、拉氏狼牙蝦虎魚等11種;冬季優勢種為三疣梭子蟹和脊尾白蝦,重要種為棘頭梅童魚、口蝦蛄等7種。 表1 三門灣海域漁業生物優勢種和重要種的相對重要性指數(IRI) 從表2可見:春、夏、秋、冬4季漁業生物的數量密度平均值依次為17.4 ×103、89.1 ×103、15.3 ×103、7.5 ×103ind./km2,其中,4個季節最大值和最小值依次出現在15、4號站位(春季)、4、3號站位(夏季)、15、4號站位(秋季)、18、16號站位(冬季);春、夏、秋、冬4季漁業生物的質量密度平均值依次為136.4、584.0、152.5、106.6 kg/km2,其中,4個季節最大值和最小值依次出現在15、4號站位(春季)、14、3號站位(夏季)、15、8號站位(秋季)、15、8號站位(冬季)。 表2 三門灣海域漁業生物資源密度 從表3可見:春、夏、秋、冬4季物種的Shannon-Wiener多樣性指數(H′)、均勻度指數(J)和豐富度指數(D)的范圍均為夏、秋季較高,其中,4個季節H′、J的平均值均為秋季最大,分別為2.3、0.8,D的平均值為夏季最大,為3.2;單因素方差分析結果顯示,不同季節物種多樣性無顯著性差異(H′:F=0.201,P>0.05;J:F=0.291,P>0.05;D:F=1.295,P>0.05)。 表3 三門灣海域漁業生物物種多樣性指數 從圖2可見:4個季節豐度優勢度曲線與生物量優勢度曲線均存在交叉或重疊的現象,這顯示三門灣近岸海域漁業生物群落在春、夏、秋、冬4季均受到了外界中等程度的干擾;春、夏季大部分生物量優勢度曲線位于豐度曲線之上,而秋、冬季則相反,這表明秋、冬季節漁業生物群落受到外界干擾的程度較春、夏季大,且夏季受到外界干擾最小,秋季最大。 圖2 三門灣海域漁業生物群落ABC曲線 從圖3可見,NMDS排序分析可知stress系數為0.1,這表明排序結果可用NMDS的二維點圖表示,且能較清楚地反映同一群組內的相似性排序結果。 SP1~SP20依次為春季S1~S20站位;SU1~SU2依次為夏季S1~S20站位;AU1~AU20依次為秋季S1~S20站位;WI1~WI20依次為冬季S1~S20站位,下同。 從圖4可見:不同季節內部具有較高的相似性,即春季所有站位(B組)在41.95%的相似性水平上聚為一組,夏季所有站位(A組)在46.30%的相似性水平上聚為一組,秋季所有站位(D組)在53.05%的相似性水平上聚為一組,冬季所有站位(C組)在55.89%的相似性水平上聚為一組;秋季(D組)和冬季(C組)可在41.20%的相似性水平上聚為一組。 圖4 三門灣近岸海域漁業生物群落結構的組平均聚類分析 從表4可見:春、夏、秋、冬4季主要漁業生物生態位寬度值的變化范圍依次為0.61~4.45、0.18~5.98、0.31~5.80和0.71~5.06;春季生態位寬度值最大為三疣梭子蟹,最小為長吻紅舌鰨,夏季生態位寬度最大為小黃魚,最小為鮸,秋季生態位寬度最大為龍頭魚,最小為擬穴青蟹,冬季生態位寬度值最大為三疣梭子蟹,最小為日本蟳。 表4 三門灣海域主要漁業生物生態位寬度值 三門灣海域漁業生物群落各營養級物種組成見表5。從表6和表7可見:春、秋、冬季均為第三營養級種類的個體數量和生物量所占百分比最大,依次為85.3%和80.9%(春季)、58.8%和56.4%(秋季)、96.4%和84.2%(冬季),夏季第四營養級物種的個體數和生物量所占百分比最大,分別為48.1%和55.2%;夏、秋、冬季第二營養級種類的個體數和生物量所占百分比最小,依次為12.7%和2.9%(夏季)、10.1%和6.9%(秋季)、1.1%和0.8%(冬季),春季第四營養級物種的個體數和生物量所占百分比最小,分別為2.7%和8.1%。由此可見,在不考慮浮游動物的情況下,第二營養級和第四營養級種類的個體數和生物量總體上較第三營養級小,三門灣海域漁業生物群落的第二、三、四營養級結構呈現異常的“鼓形”現象。 表5 三門灣海域漁業生物營養級種類組成 表7 三門灣海域漁業生物群落3個營養級種類的生物量結構 本次調查得到春季漁業生物66種(游泳動物55種)、夏季84種(游泳動物72種)、秋季69種(游泳動物60種)、冬季漁業生物42種(游泳動物40種),較冀萌萌等[9]2015年春季和2014年秋季在三門灣海域調查到42種和31種游泳動物種類數有所增加,而數量密度和質量密度在春、秋兩季均出現下降。夏季漁業生物資源量遠高于其他季節,這得益于在中國不斷完善的伏季休漁制度,以及近年來浙江省開展的漁場修復振興暨“一打三整治”專項執法行動,這與聶振林等[31]、同春芬等[32]的研究結果相一致。 與冀萌萌等[9]的研究結果相比,三門灣海域漁業生物的優勢種存在較大變化,如2015年春季(5月)六絲矛尾蝦虎魚和棘頭梅童魚為優勢種,而在2018年春季(4月)被日本蟳和細螯蝦代替,這可能是春季調查時,大黃魚、小黃魚、棘頭梅童魚、花鱸等近距離洄游性魚類尚未進入三門灣海域,而日本蟳、三疣梭子蟹等甲殼類已先期進入該海域所致。另外,三疣梭子蟹在2014年秋季、2015年春季及2018年4個季節調查中均為優勢種,且在2015年春季和2018年春、秋、冬季中均為第一優勢種,對第二、三、四營養級結構“鼓形”現象的形成具有很大的作用。三疣梭子蟹在2018年春、秋、冬季的生態位寬度值也較大,這表明其分布范圍廣、數量多,對資源和環境的適應能力較強,在資源與環境的競爭中占據較大優勢。據查詢,近年來三門灣海域沿岸的三門縣、寧海縣及象山縣等對三疣梭子蟹開展了大規模的增殖放流,這可能是導致三疣梭子蟹數量、質量明顯提高及分布范圍擴大的原因。 本研究表明,三門灣海域漁業生物物種多樣性總體呈現夏、秋季大于春、冬季的特點。夏季豐富度指數平均值最高,主要原因可能是臺灣夏季暖流已完全進入三門灣海域,海水水溫升高,沿岸浮游生物大量繁殖,為漁業生物提供了適宜的生存條件和豐富餌料,另外,夏季調查正處于休漁期,因此,漁業生物群落豐富度最高,這與郭建忠[33]的研究結果一致;Shannon-Wiener多樣性指數和均勻度指數平均值夏季低于秋季,這可能是夏季優勢種占有絕對地位,導致物種分布不均勻,從而使得這兩個指數值在夏季略低于秋季,這與梅春[34]的研究結果相似。 Shannon-Wiener多樣性指數和ABC曲線研究結果均表明,三門灣海域4個季節漁業生物群落均受到中度的干擾,呈現出三疣梭子蟹、龍頭魚等生長速度快、繁殖能力強的r選擇種類(有利于增大內稟增長率的選擇)所占優勢較大,這可能是由于伏季休漁結束后,大黃魚、花鱸等k選擇種類(有利于競爭能力增加的選擇)被大量捕撈,導致漁業生物群落中r選擇種類逐步占據優勢,群落的穩定性發生變化,這與梁海等[35]、程家驊等[36]的研究結果一致。本研究顯示,三門灣海域夏季漁業生物群落受干擾程度最小,主要原因是夏季調查正處于休漁期,有別于其余3個季節,與未受高強度的捕撈干擾有關,這表明漁業捕撈仍然是影響三門灣海域海洋生物群落結構的重要因素。另外,沿岸工業、核電站及發達的養殖業等也可能是漁業生物群落受干擾的重要來源。 本研究表明,三門灣近岸海域漁業生物群落季節分布結構呈現同一季節內各站位相似性較高,而不同季節異質性顯著的現象。究其原因:一是調查時間的跨度較大,導致非生物環境因素(溫度、鹽度、海流等)變化較大,三門灣為半封閉式海灣,水文狀況復雜,在淡水徑流和外海水季節交替的共同作用下,灣口水域生態環境具有顯著的季節差異[37];二是不同生物種類因季節性差異產生的產卵、索餌、越冬等行為,導致群落結構顯示時間異質性。春季一些長距離洄游、沿岸產卵物種,如海鰻、鮻等種類隨臺灣暖流向北向岸延伸分布,降低了各站位物種分布的相似性;夏季產卵結束的大個體繼續洄游,留下大量幼體則在附近索餌育肥,該類群的物種數、個體數等比值顯著提升,各站位相似性較春季有所提高;冬季航次調查時期,大部分洄游性種類前往較深海域或外海越冬,留下龍頭魚、蝦虎魚、日本蟳等定居性或短距離洄游的土著種類,因此,該季各站位相似性最高。 本研究表明,不同季節漁業生物的生態位寬度值存在較大差異,同一種類在不同季節的生態位寬度也會產生差異,這可能是受不同種類自身生物學特性及非生物環境因素(如底質、鹽度、水溫等)影響所致,而生態位寬度的季節間差異也反映群落內不同種類的分布、生態適應性及對資源和環境的利用能力存在差異,造成不同種類在群落中所占地位發生變化,這也可能導致群落出現季節異質性,這與吳東麗等[38]的研究結果相似。 本研究表明,三門灣海域漁業生物的營養級結構呈現出第三營養級種類的個體數量和生物量均較第二、第四營養級大,由此可見,在不考慮浮游動物生物量情況下,3個營養級的結構均呈現出畸形的“鼓形”現象,因此,傳統的能量金字塔結構可能遭受破壞。據查詢,多年來浙江海洋增殖放流種類多為營養級較高的兇猛肉食性類,放流了大量的第三、四營養級種類,這就大幅增加了第三、四營養級種類資源量,加速了第二營養級種類資源量的減少,而浙江主流捕撈漁具為底拖網、流刺網及張網等,漁獲對象多為第四營養級和部分的第三營養級種類,長期的高強度捕撈居高不下,這一系列因素導致了第四營養級種類資源量較第三營養級少。綜上所述,多種因素造成營養級結構整體上呈現畸形的“鼓形”現象。因此,三門灣海域漁業生物群落近幾年出現的營養級結構畸形的“鼓形”現象應該得到學術界與管理部門重視。建議今后三門灣海域漁業資源增殖放流、海洋生態修復及生產管理措施的制定應以此生物群落變化特征為參考,遵循科學規律,實現因生境制宜。 1)在伏季休漁制度和“一打三整治”專項執法工作順利收官的背景下,三門灣海域漁業生物種類數得到恢復,但在高強度捕撈影響下,漁業資源衰退情況并未得到遏制,蝦虎魚類、非重要經濟蝦蟹類等種類仍在該海域占有較大優勢。 2)三門灣海域漁業生物群落在2018年4個季節中均受到中等程度的干擾,高強度捕撈壓力、沿岸工業、核電站及發達的養殖業等因素均對漁業生物群落的穩定性造成了一定影響。 3)三門灣海域漁業生物群落因物種生物學特性差異、環境變化等呈現季節異質性,因此,在漁業生物群落修復時需遵循因生境制宜、因種制宜、因時制宜等原則。 4)第三、四營養級種類的大量增殖放流,導致第二營養級種類資源量大幅減少,在強大的捕撈壓力下,第四營養級與部分第三營養級種類的資源量出現下降,最終造成三門灣海域漁業生物營養級結構呈現“鼓形”現象,因此,漁業資源增殖放流策略亟待研究與改革。2 結果與分析
2.1 種類組成
2.2 優勢種和重要種

2.3 資源密度

2.4 物種多樣性指數

2.5 ABC曲線
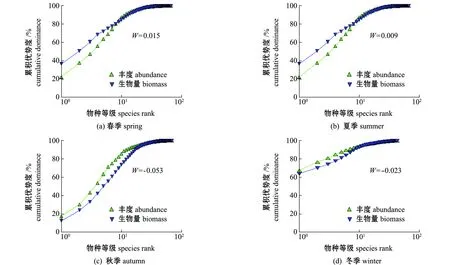
2.6 群落結構
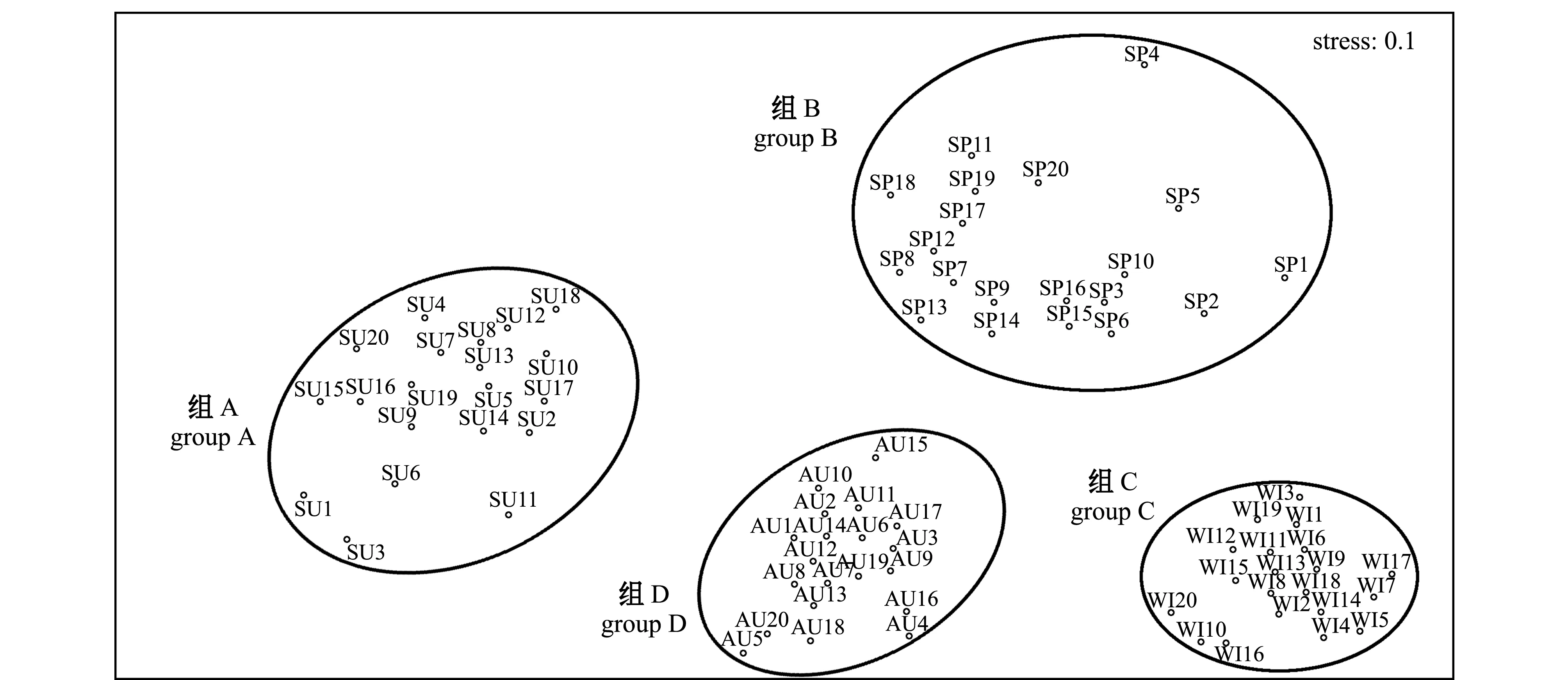
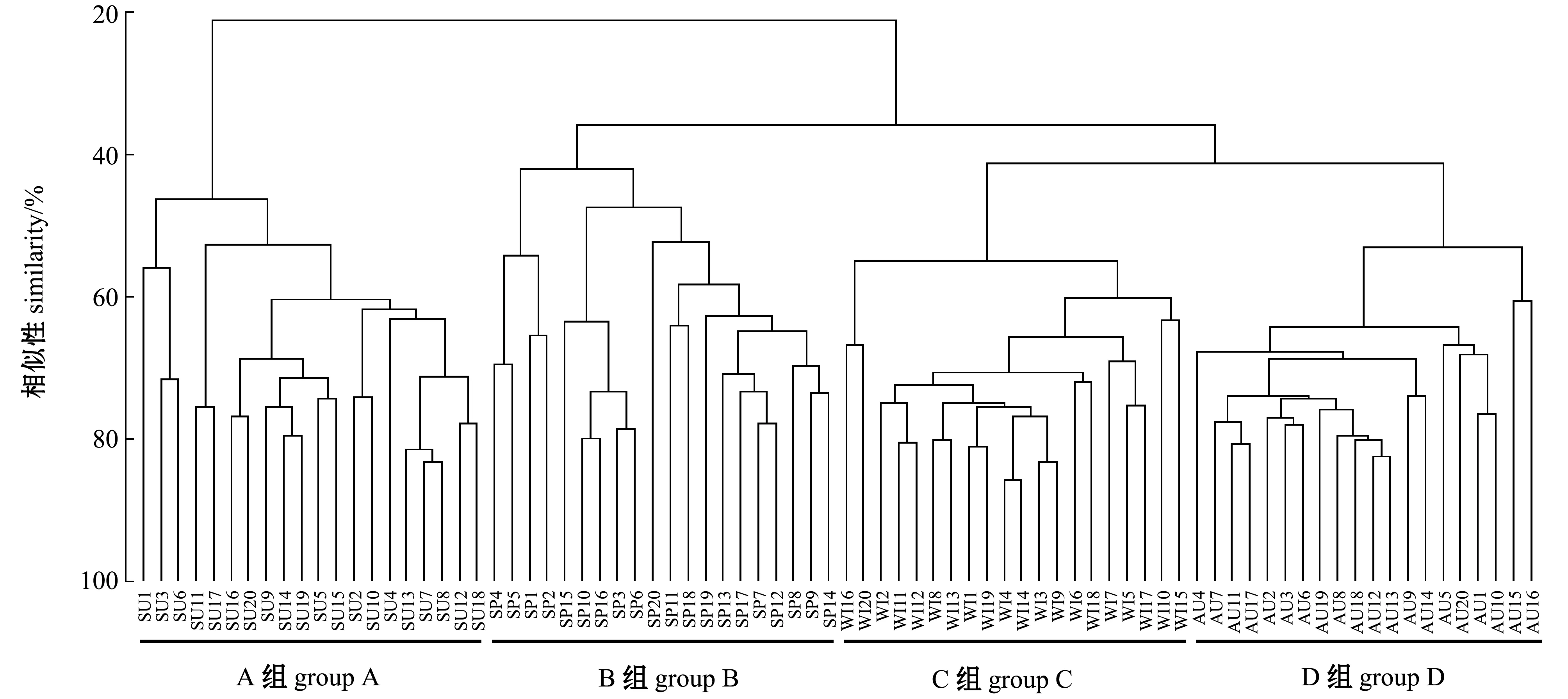
2.7 生態位寬度

2.8 營養級結構

3 討論
3.1 三門灣海域漁業生物種類組成特征
3.2 三門灣海域漁業生物群落受干擾程度
3.3 三門灣海域漁業生物群落結構特征
3.4 三門灣海域漁業生物營養級結構變化
4 結論
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18天天愛科學(2022年9期)2022-09-15 01:12:54天天愛科學(2022年4期)2022-05-23 12:41:48當代水產(2022年3期)2022-04-26 14:26:56今日農業(2021年11期)2021-08-13 08:53:34航空世界(2020年10期)2020-01-19 14:36:20科學大眾(中學)(2019年3期)2019-05-17 10:04:30中國生殖健康(2019年8期)2019-01-07 01:18:24幸福(2018年33期)2018-12-05 05:22:46汽車觀察(2018年10期)2018-11-06 07:05:26
