抗弧菌殺魚假交替單胞菌2515的潛在毒性及熱處理脫毒后抗菌效果
2021-06-21 09:02:36張宇哲王秀華于黨輝王楓林孫祥山黃經獻
大連海洋大學學報 2021年3期
張宇哲,王秀華,于黨輝,王楓林,孫祥山,黃經獻
(1.中國水產科學研究院黃海水產研究所 農業農村部海水養殖病害防治重點實驗室, 青島市海水養殖流行病學與生物安保重點實驗室,山東 青島 266071; 2.上海海洋大學 水產與生命學院,上海 201306; 3.青島海洋科學與技術試點國家實驗室 海洋漁業科學與食物產出過程功能實驗室,山東 青島 266071; 4.中國水產科學研究院下營增殖站,山東 濰坊 261312)
弧菌病是海水養殖動物的重要病害之一,給海水魚、蝦和貝類養殖帶來了嚴重危害。目前,針對弧菌病的生物防控,研究人員已發現有多種微生物對弧菌具有拮抗活性[1-3],其中,殺魚假交替單胞菌Pseudoalteromonaspiscicida是極具開發潛力的一種菌,該菌不僅抗菌譜廣[4],而且還可提高動物的免疫力,將殺魚假交替單胞菌用于養殖期對蝦及貝類也表現出良好的生物安全性[5-6]。然而也有研究顯示,有些殺魚假交替單胞菌分離株可感染克氏雙鋸魚Amphiprionclarkii卵,導致其存活率較低[7]。喬毅等[8]報道,一株與殺魚假交替單胞菌同源性較近的菌株對黑鯛表現出一定的致病性。
細菌毒性來自其含有的內外毒素,內毒素主要為細菌脂多糖[9],外毒素種類較多,按照作用的靶組織,分為細胞毒素、神經毒素和腸毒素[10]。假交替單胞菌多為海洋菌株,其內毒素毒性通常較低[11],但研究發現,有些假交替單胞菌可分泌細胞毒素,誘導白血病細胞凋亡或溶血[12-13],而有些種類能夠分泌神經毒性的河豚毒素[14],目前對殺魚假交替單胞菌的毒性研究尚未見報道。本研究中,針對前期篩選出的一株具有廣譜弧菌拮抗特性的殺魚假交替單胞菌,測試了其對中國明對蝦Fenneropenaeuschinensis受精卵及幼體發育的影響,以及對凡納濱對蝦Litopenaeusvannamei、魚及羊血細胞的溶血活性,為去除其潛在毒性,還探討了熱處理對菌株滅活、溶血活性和抗菌物質活性的影響,以期為該菌的開發應用提供技術及方法支撐。
1 材料與方法
1.1 材料
試驗用中國明對蝦受精卵、無節幼體和蚤狀幼體由中國水產科學研究院下營增殖實驗站提供。
殺魚假交替單胞菌2515(簡稱菌株2515)、地衣芽孢桿菌Bacilluslicheniformis、對照組假交替單胞菌2905(簡稱菌株2905)和金黃色葡萄球菌Staphylococcusaureus均由農業農村部海水養殖病害防治重點實驗室保存。TCBS培養基、羊血平板均購自北京路橋技術股份有限公司。
1.2 方法
1.2.1 培養基及血平板制備 TCBS培養基:配制方法參照說明書。2216E培養基:胰蛋白胨5 g、酵母膏1 g、FePO4·4H2O 0.01 g、瓊脂粉16 g(固體培養基)、陳海水1 L,pH 7.8,121 ℃高壓滅菌15 min。
魚血平板制備:用無菌注射器抽取大菱鲆尾靜脈血液,置入等體積的阿氏液(Na3C6H5O7·2H2O 1.60 g、C6H8O70.11 g、C6H12O64.10 g、NaCl 0.84 g、蒸餾水200 mL,pH 6.1,115 ℃滅菌30 min)輕輕搖勻備用。將滅菌后的2216E固體培養基冷卻至40~50 ℃時,加入10%的魚血備用液,搖勻后倒平板。
1.2.2 菌液制備 將菌株2515和地衣芽孢桿菌分別接種于2216E固體平板,于28 ℃下培養活化后再接種到液體培養基中,在28 ℃ 下以150 r/min搖床培養24 h,菌液經過6 000×g離心10 min,將沉淀用海水稀釋成1×109CFU/mL菌懸液,4 ℃下冷藏帶到試驗場地,根據試驗設計濃度稀釋后使用。
1.2.3 菌株2515對中國明對蝦受精卵孵化及幼體發育成活影響試驗
1)幼體成活試驗。試驗用對蝦育苗袋(直徑25 cm,高40 cm)132個,分別加入經過消毒殺菌處理的育苗用海水2 L作為試驗水體。根據幼體發育階段設置3個試驗,分別測試菌株2515對中國明對蝦蝦受精卵孵化至無節幼體Ⅲ期(N3)、N3發育至蚤狀幼體Ⅰ期(Z1)、蚤狀幼體Ⅱ期(Z2)發育至糠蝦幼體Ⅰ期(M1)成活率的影響,每個試驗均設置地衣芽孢桿菌陽性對照組及空白對照組,菌株2515及地衣芽孢桿菌均設置105、106、107、108CFU/mL 4個濃度,每個濃度設3組平行。蝦受精卵孵化至N3期試驗中,每個袋投入中國明對蝦受精卵800個;N3至Z1期、Z2至M1期試驗中,每個袋中投入相應期別的幼體40個。在受精卵孵化至N3期、 N3發育至Z1期試驗中,試驗水體中不添加任何餌料;Z2發育至M1期試驗中,每袋添加鹵蟲幼體600個,經充氧氣后打包,放在育苗池中水浴。受精卵孵化期、N3發育至Z1期、Z2發育至M1期的溫度分別控制在17、20、23 ℃,試驗用海水pH 7.8、鹽度30。在各期試驗結束后,將試驗袋內幼體用網目為125 μm的篩絹網過濾到燒杯中逐尾計數,并統計成活率:
成活率=試驗結束時存活數量/試驗初始投放數量×100%。
2)幼體發育觀察。中國明對蝦受精卵孵化至N3期時,從各組隨機挑出無節幼體5尾,通過光鏡觀察幼體體表、附肢和活力情況,評價菌株2515對幼體發育的影響。
1.2.4 溶血試驗
英美新批評理論是對19世紀西方文學研究中對社會歷史、政治、傳記、心理等傾向的反撥與糾偏,以此重新突顯文學研究中作品文本中心這一向度。作為一種策略,文本細讀以及其他的策略,使得這一批評理論流派得以確立。美國華人學者與英美新批評之間有著復雜的糾纏,他們自覺或不自覺地在現代漢詩的相關研究中透露出英美新批評的理論視角與觀念方法。然而,理論的旅行總會發生變異,因具體歷史文化語境與觀照對象的變化,美國華人學者在英美新批評視野下開展的現代漢詩批評實踐也產生不一樣的效應。這些效應彰顯了他們的研究所具有的詩學意義與文化價值。
1)血平板溶血觀察。刮取菌株2515、金黃色葡萄球菌及菌株2905的2216E平板菌落,分別用PBS制成濃度為109CFU/mL的菌懸液,各取5 μL等距點種在羊血瓊脂和魚血瓊脂平板上,每種菌株設3個重復。28 ℃恒溫培養箱培養48 h,觀察溶血情況。
2)蝦細胞溶血顯微觀察。從凡納濱對蝦Litopenaeusvannamei圍心腔抽取血液淋巴與等體積的阿氏液混合作為對蝦血細胞懸液,取5 μL對蝦血細胞懸液與等體積的菌株2515發酵液(濃度108CFU/mL)混合后,滴加在載玻片上,改好蓋玻片,置于濕盒中4 ℃條件下計時存放,顯微鏡下間隔1 h觀察1次蝦血細胞的溶血情況,并設置生理鹽水及大菱鲆血細胞對照組。
1.2.5 熱處理對菌株2515的滅活效果 取發酵24 h的菌株2515發酵液,用PBS稀釋成103CFU/mL,分裝到6個無菌的1.5 mL離心管中,每管400 μL,分別置于40、45、50、55、60、65 ℃的水浴鍋中加熱處理1 h,空白組常溫放置1 h,之后從每管取100 μL涂布于2216E平板。每個樣品設3個平行,28 ℃下培養,觀察有無菌落形成,統計菌落數量。
1.2.6 熱處理后菌株2515的溶血活性 取“1.2.5節”中經40 ℃和55 ℃不同時間熱處理后的菌株2515處理液,按照“1.2.4”節中血平板溶血觀察方法,點種在魚血與羊血平板上,觀察處理液的溶血活性。
1.2.7 熱處理對菌株2515抑菌活性的影響 取菌株2515發酵液(濃度3.9×109CFU/mL)110 mL,以5 000×g離心10 min,棄上清,將沉淀用PBS洗滌2次后,加50 mL PBS重懸,用超聲波細胞破碎儀超聲破碎10 min(300 W),至菌液澄清。將澄清的菌液經0.22 μm孔徑的細菌過濾器過濾,之后置于平皿中-20 ℃下真空干燥12 h。再用20 mL PBS將干燥后的產物洗脫混勻,之后分裝到無菌1.5 mL離心管中(每管0.2 mL),分別在40、45、50、55、60、65、75 ℃水浴鍋中進行溫浴處理,每個溫度設置15、30、45、60 min 4個處理時間段,對照組常溫放置,每組設3個平行。完成熱處理后參照李萍等[15]的方法進行抑菌活性測定,測試用2216E平板含有鰻弧菌Vibrioanguillarum濃度為107CFU/mL,經28 ℃培養24 h后,用游標卡尺十字交叉法測量抑菌圈直徑,根據抑菌圈直徑大小計算菌株2515抗菌活性物質的抑菌活性增長率,即
抑菌活性增長率=(De-D0)/D0×100%。
其中,De、D0分別為試驗組和對照組抑菌圈直徑。
1.3 數據處理
試驗結果以平均值±標準差(mean±S.D.)表示。采用SPSS 16.0軟件進行單因素方差分析,采用Duncan法進行組間多重比較,顯著性水平設為0.05。
2 結果與分析
2.1 菌株2515對中國明對蝦幼體發育的影響
從圖1可見:相同濃度組中,添加菌株2515與添加地衣芽孢桿菌陽性對照組無節幼體的成活率均無顯著性差異(P>0.05);在所有試驗組中,添加濃度為105CFU/mL 2515菌株的幼體成活率最高,顯著高于添加濃度為106、108CFU/mL的地衣芽孢桿菌組(P<0.05),也顯著高于添加濃度107、108CFU/mL的菌株2515組(P<0.05);空白對照組與各濃度組均無顯著性差異(P>0.05)。顯微觀察各組無節幼體的外形特征,可見各試驗組無節幼體活力較好,體表干凈,附肢發育正常,未出現彎曲現象。這表明,所選擇的濃度范圍內,菌株2515對中國明對蝦受精卵孵化至N3期的成活率無明顯影響。
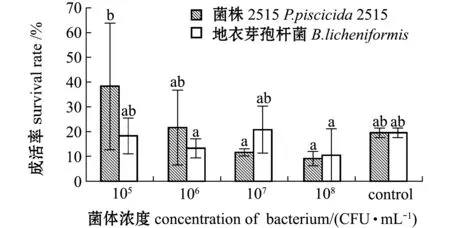
標有不同字母者表示組間有顯著性差異(P<0.05),標有相同字母者表示組間無顯著性差異(P>0.05),下同。
從圖2可見:各濃度組對蝦的成活率差別較大,相同濃度組中,添加菌株2515與添加地衣芽孢桿菌陽性對照組的成活率中僅濃度為105CFU/mL的組與空白對照組無顯著性差異(P>0.05),其他組均顯著低于空白對照組(P<0.05),而添加濃度為106、107CFU/mL的兩組中,添加2515菌株的對蝦成活率顯著低于添加地衣芽孢桿菌組(P<0.05);在所有添加菌株2515的組中,隨著添加濃度的升高對蝦的存活率呈降低趨勢。這表明,在本試驗條件下,菌株2515濃度高于105CFU/mL時對中國明對蝦由N3期發育至Z1期有顯著性影響,可顯著降低無節幼體變態發育至蚤狀幼體的成活率。
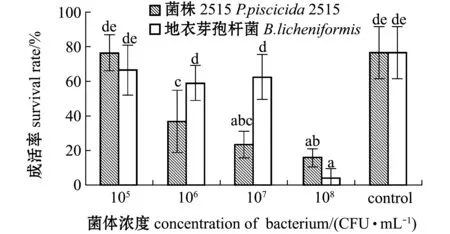
圖2 N3到Z1期變態成活率
從圖3可見,各試驗組與添加地衣芽孢菌組及空白對照組的對蝦成活率均無顯著性差異(P>0.05)。

圖3 Z2到M1期變態成活率
2.2 菌株2515的溶血性
從圖4可見,菌株2515在兩種血平板上均具有溶血活性,陽性對照組金黃色葡萄球菌在羊血平板上產生溶血,對照組假交替單胞菌株2905在羊血平板上未產生溶血,而在魚血平板上產生溶血。
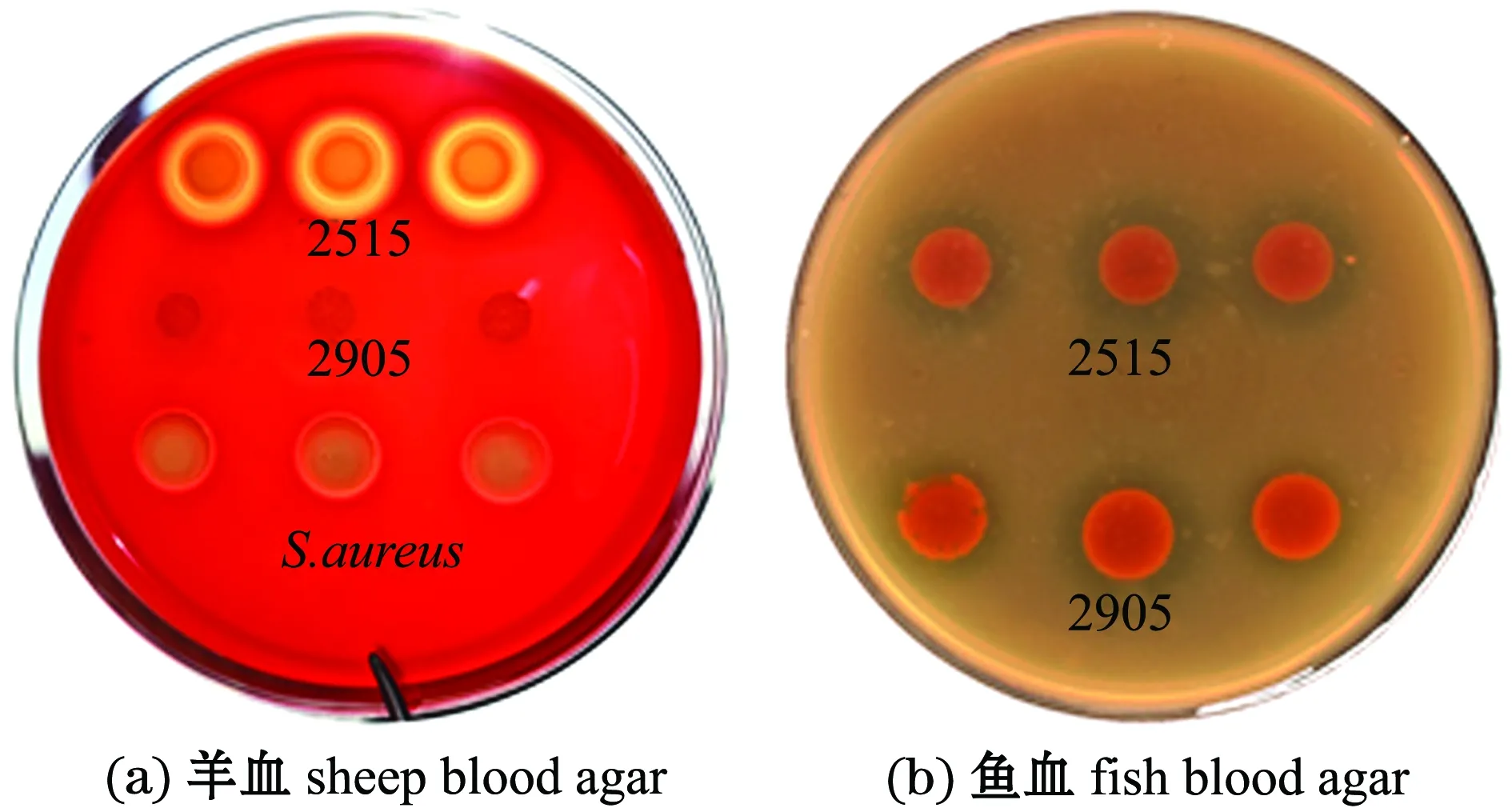
圖4 菌株2515對羊血和魚血的溶血情況
大菱鲆血細胞試驗組,在加入菌株2515 3 h后,紅細胞出現細胞膜溶解(圖5C),胞內物質固縮變形,有些趨于解體,白細胞則不發生溶血,而大菱鲆生理鹽水對照組放置18 h后(圖5B),白細胞及紅細胞的形態仍然完整;對蝦血細胞試驗組(圖5G)與對蝦生理鹽水對照組(圖5F)中,在加入菌株2515及生理鹽水18 h時,兩組對蝦血細胞形態相似,細胞的形態均趨于收縮變圓,但外形完整,未出現溶血現象。
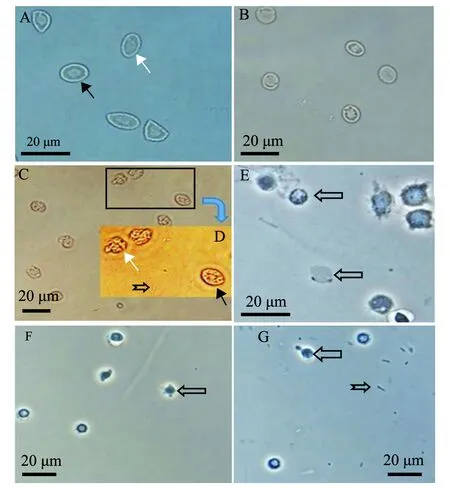
A—試驗開始前大菱鲆血細胞;B—在生理鹽水中18 h的大菱鲆血細胞;C—在菌液中3 h的大菱鲆血細胞;D—圖C中方框區域的放大圖片;E—溶血開始前蝦血細胞;F—在生理鹽水中18 h的蝦血細胞;G—在菌液中18 h的蝦血細胞。圖中白色箭頭為大菱鲆紅細胞;黑色箭頭為大菱鲆白細胞;左向空心箭頭為不同類型蝦血細胞;魚尾箭頭為菌體。
2.3 熱處理對菌株2515的滅活效果
菌株2515經不同溫度滅活1 h后,涂布于2216E平板培養,發現40 ℃處理組中,平板上菌株2515菌落數與未處理組數量相近,分別為(88±12)CFU/平板與(90±8)CFU/平板,在45 ℃處理組中2515菌落數為(77±5)CFU/平板,而在50、55、60、65 ℃處理組平板上,無細菌生長,表明50 ℃熱處理1 h即可將該菌滅活。
2.4 熱處理后菌株2515的溶血活性
破碎液經40、55 ℃處理15~60 min后,各組破碎液失去對魚紅細胞溶血作用(圖6A、B),而對羊紅細胞的溶血效果依然保持較高的活性(圖6C、D)。

A、B—40、55 ℃處理組大菱鲆血平板;C、D—40、55 ℃處理組羊血平板。
2.5 熱處理對菌株2515抑菌活性的影響
從圖7、圖8可見:當處理溫度為40~65 ℃時,各處理時間組的抑菌活性均比對照組高,但抑菌活性存在差別;當溫度為55 ℃時,抑菌活性增長最高,隨著處理時間從15 min增加到60 min,抑菌活性增長率由46.1%提高到55.8%;當溫度為65 ℃時,處理15 min活性增加最高,達到33.3%,之后隨時間延長而降低;當溫度為75 ℃時,處理15 min即可降低抑菌活性,處理45 min時即可使抑菌活性完全失活,增長率達到-100%。

圖7 菌株2515在不同溫度下滅活不同時間對鰻弧菌的抑菌效果
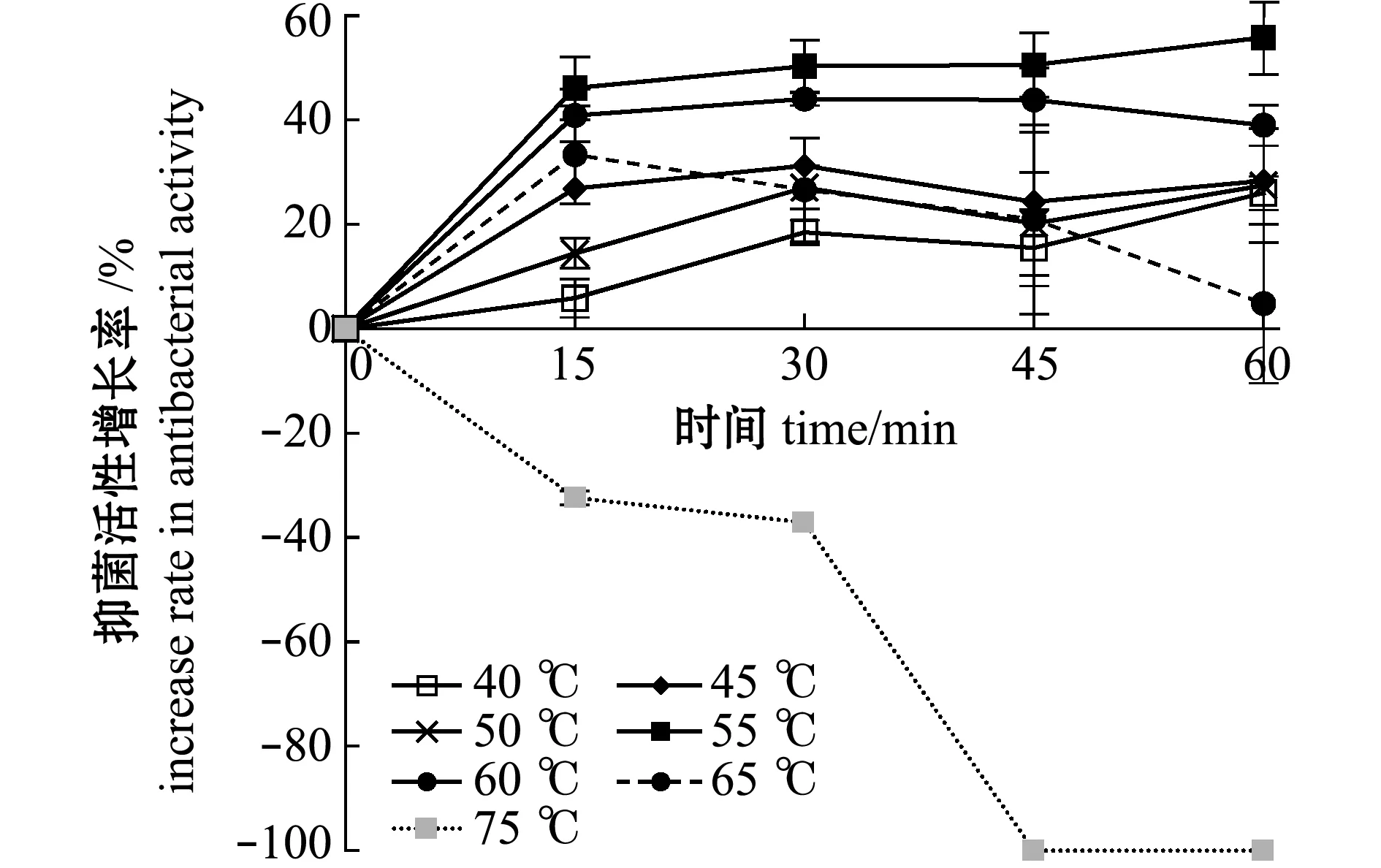
圖8 熱處理菌株破碎液后抑菌物質活性的增長率
3 討論
3.1 殺魚假交替單胞菌生物學功能
目前,已發現假交替單胞菌屬細菌中有20余種能夠產生活性物質,其功效包括降解瓊膠、抗細菌、抗真菌、殺菌、促進水生動物幼體附著等[16]。殺魚假交替單胞菌作為其中的一個種,不僅能分泌廣譜抗菌活性物質[17],還可產生蛋白酶、脂肪酶、淀粉酶和甘露糖酶,用該菌作益生菌并通過鹵蟲作為載體投喂對蝦幼體,能夠提高對蝦生長率、存活率和腸道總菌數[18]。菌株2515是從對蝦養殖池分離出的一株殺魚假交替單胞菌[4],該菌株具有廣譜抗弧菌特性,而對其生物安全性尚未有系統研究。本研究中,初步確定了該菌對魚類的溶血毒性及對中國明對蝦幼體的潛在毒性,獲得了對已知毒素的熱減毒方法,而對其潛在的其他毒性物質種類及致病力尚需后續開展研究。
3.2 菌株2515的溶血素及其熱穩定性
溶血素為細菌的外毒素,種類較多,其均為單鏈多肽蛋白,溶血素的溶血活性是衡量細菌毒力的因子之一,根據溶血素結構、其與細胞結合的方式及形成孔道的機制等不同,可將溶血素分為重復子毒素家族和硫醇活性膽固醇結合細胞溶血素家族[19]。溶血素可導致細胞溶解或裂解、細胞膜損傷等病變,進一步導致細胞內鈉、鉀、鈣離子失衡及細胞凋亡[20]。溶血素在同種細菌的不同分離株中,其基因編碼、數量和基因表達存在差異[21],弧菌中存在5類有代表性的溶血素家族,其中,包括副溶血弧菌熱穩定性溶血素和熱不穩定性溶血素[22]。目前,對殺魚假交替單胞菌溶血素的結構、分類、溶血機制等研究尚為空白,但從本研究結果可知,菌株2515同時含有熱穩定性與熱不穩定性兩種溶血素,分別對羊紅細胞及魚紅細胞具有溶血活性,其中,溶解羊血的熱穩定性溶血素對魚紅細胞不具有溶血活性。由于菌株2515在水產養殖中具有潛在開發應用價值,對該兩種溶血素在水產動物中的致病性強弱尚需進一步研究。本研究結果也提示,在進行水生動物益生菌溶血性評價時,應針對性地選擇靶動物的血細胞。
3.3 殺魚假交替單胞菌對中國明對蝦的潛在毒性
Sun等[6]將分離的一株殺魚假交替單胞菌(P.-SW-1)用于貝類養殖,發現該菌能夠減輕溶藻弧菌Vibrioalginolyticus對貝類的致病力,認為該菌株可以用作貝類的益生菌。而喬毅等[8]發現了一株與殺魚假交替單胞菌分離地位相近的假交替單胞菌屬細菌LSHD-1含有絲氨酸蛋白酶基因、溶血素基因、腸毒素基因,對黑鯛Acanthopagrusschlegel具有致病性。本研究中,菌株2515用于對蝦幼體培育,僅在N3至Z1期顯示出隨著添加劑量的增大幼體成活率呈現降低的趨勢,而在低濃度組(105CFU/mL)表現出能提高對蝦卵孵化率及N3至Z1期變態率的效果,表明該菌株對中國明對蝦幼體具有的潛在危害與使用濃度有關。有關殺魚假交替單胞菌對中國明對蝦的毒性評價及機制研究較少,但相關研究表明,殺魚假交替單胞菌能夠產生降解幾丁質的酶[23],若定殖在對蝦前腸中,可能會對中國明對蝦前腸幾丁質具有潛在的破壞作用[24]。而Z1期對蝦幼體消化道初步發育完成,為幼體開口期,需要單胞藻類為食物,試驗養殖水體中無添加餌料,幼體會濾食菌體導致菌體附著,使得前腸更易受到損傷,但該推測尚需進一步結合組織病理學研究加以證實。
3.4 殺魚假交替單胞菌熱脫毒后的抗菌效果
去除生物毒素有化學、物理及微生物等方法,其中,加熱脫毒是降解生物毒素的常用方法,加熱脫毒效果與毒素種類有關,過高的溫度處理會破壞產品中有價值的成分,因此,適宜的脫毒溫度選擇尤為重要。存在于糧食中的脫氧雪腐鐮刀菌烯醇和T-2毒素為耐高溫毒素,需105 ℃以上高溫處理方可達到部分去除效果[25]。而為提高牛奶的保質期同時保持其溶菌酶的抗菌活性,采用63 ℃ 30 min處理即可達到效果[26]。菌株2515的抗菌物質成分目前尚未闡明,但為了去除其對魚類紅細胞具有破壞作用的溶血素,滅活該菌,防止其在環境中定殖,同時維持細菌抗菌物質的活性,本研究中采用55 ℃熱處理1 h即可達到最佳效果。
4 結論
1)菌株2515直接用于對蝦育苗,影響對蝦蚤狀幼體成活率。該菌存在能夠分別溶解魚與羊紅細胞的溶血素,若用于魚類養殖存在潛在毒性。
2)菌株2515同時含有熱穩定性與熱不穩定性兩種溶血素,熱不穩定性溶血素對魚紅細胞產生溶血,而熱穩定性溶血素對羊紅細胞產生溶血,對魚紅細胞不產生溶血。
3)通過55 ℃ 1 h熱處理能夠去除菌株熱不穩定性溶血素活性,能夠滅活該菌,而對其抗菌活性無影響,熱處理可以作為該菌脫毒的一種備選方法。
