全轉(zhuǎn)錄組測序分析精子發(fā)生中RNA結(jié)合蛋白質(zhì)的動態(tài)表達
2021-06-17 06:56:48邙新雨鄒定峰李夢真繆時英王琳芳
基礎(chǔ)醫(yī)學與臨床 2021年6期
李 凱,邙新雨,鄒定峰,李夢真,繆時英,王琳芳,宋 偉
(中國醫(yī)學科學院基礎(chǔ)醫(yī)學研究所 北京協(xié)和醫(yī)學院基礎(chǔ)學院 生物化學與分子生物學系醫(yī)學分子生物學國家重點實驗室,北京 100005)
精子發(fā)生是從精原細胞發(fā)育成為成熟精子的一個復雜有序的連續(xù)細胞分化過程,主要分為3個時期:精原細胞有絲分裂期、精母細胞減數(shù)分裂期和精子形成期[1]。該過程眾多階段特異性基因的表達與調(diào)控是維持精子發(fā)生正常進行的分子基礎(chǔ)。目前,大量單細胞組學研究高精度解析了精子發(fā)生中生精細胞有序性發(fā)生的轉(zhuǎn)錄圖譜[2-6],然而,在轉(zhuǎn)錄后水平如何操控這些轉(zhuǎn)錄生成物的命運和功能進而影響生精細胞的增殖或分化仍未詳細闡明。
RNA結(jié)合蛋白質(zhì)(RNA-binding proteins,RBPs)是一類通過其功能結(jié)構(gòu)域與RNA互作并操控RNA命運和功能的蛋白質(zhì)[7]。RBPs在轉(zhuǎn)錄后水平以多種方式參與調(diào)控RNA命運,例如mRNA選擇性剪接、運輸、編輯和翻譯等,這些方式均會引起相應(yīng)的基因表達變化[8]。目前,在精子發(fā)生中已發(fā)現(xiàn)部分RBPs在轉(zhuǎn)錄后水平發(fā)揮關(guān)鍵的基因表達調(diào)控作用[9-10],然而有關(guān)RBPs在精子發(fā)生全程的動態(tài)表達圖譜仍缺乏完整認識。本研究整合小鼠6種類型生精細胞的全轉(zhuǎn)錄組測序數(shù)據(jù),系統(tǒng)分析RBPs在精子發(fā)生中的動態(tài)表達全貌、階段特異性及協(xié)同表達模式,并對其潛在功能進行預測,為闡釋精子發(fā)生的分子機制及診治男性不育相關(guān)疾病提供新的科學依據(jù)。
1 材料與方法
1.1 材料
小鼠睪丸組織6種類型生精細胞的全轉(zhuǎn)錄組測序數(shù)據(jù)(Bulk RNA-seq)來源于本實驗室前期的研究成果[11]。該數(shù)據(jù)可從美國國立生物信息中心的基因表達綜合數(shù)據(jù)庫GEO下載(https://www.ncbi. nlm.nih.gov/geo/),其登錄號為GSE145130。6種類型生精細胞分別為原始A型精原細胞(primitive type A spermatogonia, priSG-A)、B型精原細胞(type B spermatogonia, SG-B)、前細線期精母細胞(preleptotene spermatocytes, plpSC)、粗線期精母細胞(pachy-tene spermatocytes, pacSC)、圓形精子(round spermatids, rST)和長形精子(elongating spermatids, elST)。
1.2 方法
1.2.1 RNA-seq數(shù)據(jù)處理:6種類型生精細胞RNA-seq的文庫構(gòu)建、測序數(shù)據(jù)質(zhì)控、基因組比對及基因表達分析方法見參考文獻[11]。
1.2.2 基因表達熱圖:利用R語言pheatmap工具包展示RBPs在6種類型生精細胞中的基因表達水平(Fragments Per Kilobase of transcript sequence per Millions base pairs mapped, FPKM≥1)。
1.2.3 RBPs差異表達分析及功能預測:利用R語言DESeq2工具包分析6種類型生精細胞差異表達的RBPs,篩選標準為:|log2FoldChange|≥1.5且P.adjust<0.05。利用時間序列分析軟件(Short Time-series Expression Miner, STEM)分析差異表達RBPs動態(tài)表達模式,基因簇最大數(shù)目設(shè)置為50。利用ClusterProfiler工具包對6種類型生精細胞中差異表達的RBPs分別進行GO(Gene Ontology)功能富集分析(P.adjust<0.05)。
1.2.4 RBPs共表達調(diào)控網(wǎng)絡(luò)分析:WGCNA(Weighted Gene Co-Expression Network Analysis)稱為加權(quán)基因共表達網(wǎng)絡(luò)分析,通過計算基因間表達關(guān)系鑒定表達模式相似的基因模塊(Module,ME),位于同一模塊的基因共表達程度較高并且具有相似的調(diào)控作用。利用該方法分析RBPs在精子發(fā)生中的共表達網(wǎng)絡(luò),算法軟閾值設(shè)置為22,其他為默認參數(shù)。
2 結(jié)果
2.1 RBPs在精子發(fā)生中的階段特異性及動態(tài)表達模式
目前小鼠物種RBPs的數(shù)目預計為1 913個[7]。基于6種類型生精細胞的全轉(zhuǎn)錄組測序數(shù)據(jù)分析RBPs在精子發(fā)生過程的全局轉(zhuǎn)錄圖譜,結(jié)果顯示,在這6種類型生精細胞中共檢測到1 835個RBPs(FPKM≥1)(圖1A)。根據(jù)差異基因篩選標準進一步在精子發(fā)生中鑒定了519個階段相對特異表達的RBPs,其在priSG-A、SG-B、plpSC、pacSC、rST和elST中的數(shù)目分別為71、102、97、134、80和35個,其中減數(shù)分裂時期(plpSC與pacSC)的RBPs比例最高(44.5%),有絲分裂時期(priSG-A與SG-B)RBPs比例次之(33.3%),精子形成時期(rST與elST)的RBPs比例最低(22.2%)。STEM軟件分析結(jié)果顯示階段特異表達的RBPs在精子發(fā)生中主要具有7種動態(tài)表達模式(Cluster 1-7)(圖1B)。
2.2 RBPs在精子發(fā)生中的潛在調(diào)控作用
為了進一步預測RBPs在精子發(fā)生中的潛在調(diào)控作用,分別對階段特異表達的RBPs進行GO功能富集分析(P.adjust<0.01)。結(jié)果發(fā)現(xiàn)priSG-A特異表達RBPs主要富集在RNA剪接(RNA splicing)和mRNA加工(mRNA processing)等(圖2A);SG-B特異表達RBPs主要富集在核糖核蛋白復合物生成(ribonucleoprotein complex biogenesis)和核糖體生成(ribosome biogenesis)等(圖2B);plpSC特異表達RBPs主要富集在RNA剪接(RNA splicing)和mRNA加工(mRNA processing)等(圖2C);pacSC特異表達RBPs主要富集在核糖核蛋白復合物生成(ribonucleoprotein complex biogenesis)和核糖體生成(ribosome biogenesis)等(圖2D);rST特異表達RBPs主要富集在RNA代謝(mRNA metabolic process)和翻譯(regulation of translation)等(圖2E);elST特異表達RBPs主要富集在RNA翻譯(regulation of translation)和RNA代謝(mRNA metabolic process)(圖2F)。
2.3 RBPs在精子發(fā)生中的共表達網(wǎng)絡(luò)分析
RBPs在細胞發(fā)育過程中通常會形成互作復合體與RNA互作并操控RNA命運和功能,因此鑒定RBPs共表達模式將有利于進一步發(fā)現(xiàn)其在精子發(fā)生中的重要調(diào)控作用。利用WGCNA分析階段特異表達RBPs在精子發(fā)生中共表達調(diào)控網(wǎng)絡(luò),結(jié)果共獲得5個共表達基因模塊(ME 1-5),并計算各模塊間的關(guān)聯(lián)性(相關(guān)系數(shù)>0.9)(圖3A)。根據(jù)RBPs之間的表達量進行聚類繪制得到RBPs共表達調(diào)控網(wǎng)絡(luò), 這5個共表達基因模塊內(nèi)的RBPs數(shù)量分別為285、145、219、77和76個(圖3B)。將關(guān)聯(lián)性得分最高的RBPs作為某一RBPs的潛在共表達對象,取交集去掉重復后共獲得246個共表達RBPs。STEM軟件分析結(jié)果顯示這些共表達RBPs在精子發(fā)生中主要具有7種動態(tài)表達模式(Cluster 1-7),每一種表達模式內(nèi)的RBPs數(shù)目分別為19、28、45、36、21、30和26個,其中減數(shù)分裂時期(Cluster 3-5)共表達RBPs比例最高(49.8%),精子形成時期(Cluster 6-7)共表達RBPs比例次之(27.3%),有絲分裂時期(Cluster 1-2)共表達RBPs比例最低(22.9%)(圖3C)。

A.heat map showing global transcriptional profile of RBPs in spermatogenesis; B.STEM showing the stage specificity and dynamic expression pattern of RBPs in spermatogenesis
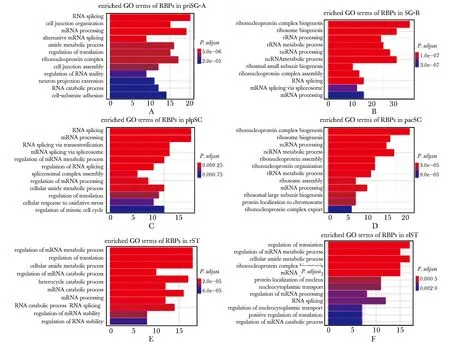
A-F.enriched GO terms of stage-specific RBPs in priSG-A, SG-B, plpSC, pacSC, rST and elST圖2 階段特異表達的RBPs在精子發(fā)生中的GO功能富集分析Fig 2 GO enrichment analysis of stage-specific RBPs in spermatogenesis

A.heatmap showing the correspondence between co-expression modules; B.heatmap showing WGCNA analysis of RBPs; C.heatmap showing the dynamic expression pattern of RBPs co-expression in spermatogenesis
2.4 共表達RBPs在精子發(fā)生中的潛在調(diào)控作用
利用GO功能富集分析預測共表達RBPs在精子發(fā)生中的潛在調(diào)控作用(P.adjust<0.05)。priSG-A共表達RBPs數(shù)量較少,并未富集到明顯的GO功能條目。SG-B共表達RBPs主要富集在核糖核蛋白復合物生成(ribonucleoprotein complex biogenesis)和核糖體RNA代謝(rRNA metabolic process)等(圖4A);plpSC共表達RBPs主要富集在RNA剪接(RNA splicing)和mRNA加工(mRNA processing)等(圖4B);pacSC共表達RBPs主要富集在蛋白質(zhì)定位(protein localization)和核糖核蛋白復合物生成(ribonucleoprotein complex biogenesis)等(圖4C);rST和elST共表達RBPs表達模式較為相似,作為一個基因集進行GO富集分析,主要富集在核糖體生成(ribosome biogenesis)和翻譯調(diào)控(regulation of translation)等(圖4D)。
3 討論
根據(jù)分子生物學中心法則,以往被視為遺傳信息傳遞中間站的信使RNA (mRNA)其實具有遠超人們所理解的轉(zhuǎn)錄后調(diào)控方式,例如mRNA選擇性剪接、編輯、運輸和翻譯等,而這些調(diào)控方式主要是由RBPs介導完成,最終引起相應(yīng)的基因表達變化。本研究系統(tǒng)描繪了RBPs在精子發(fā)生中的動態(tài)表達全貌、階段特異性及協(xié)同表達模式,并對其潛在功能進行了預測。
基因表達的轉(zhuǎn)錄后調(diào)控對于維持精子發(fā)生的正常進行至關(guān)重要[12]。精子發(fā)生早期階段基因轉(zhuǎn)錄異常活躍,大量mRNA被轉(zhuǎn)錄生成后,可與RBPs相互作用形成mRNP復合物進行存儲,在精子發(fā)生后期階段由于染色質(zhì)高度壓縮,基因轉(zhuǎn)錄活性逐漸降低。然而,為了維持生精細胞的正常發(fā)育,早期轉(zhuǎn)錄并存儲的mRNA在此時開始進行翻譯,該現(xiàn)象稱為“轉(zhuǎn)錄-翻譯”解偶聯(lián)[13]。目前,在精子發(fā)生中已發(fā)現(xiàn)部分RBPs在轉(zhuǎn)錄后水平發(fā)揮關(guān)鍵的基因調(diào)控作用[14-16]。中科院生化與細胞研究所劉默芳教授團隊發(fā)現(xiàn)MIWI/piRNA通過與翻譯起始因子eIF3f及AU-rich元件結(jié)合蛋白HuR等相互作用,激活生精細胞mRNA的翻譯并調(diào)控生精細胞發(fā)育[9]。本研究發(fā)現(xiàn)RBPs在精子發(fā)生過程呈現(xiàn)階段特異性表達,并且在早期階段(例如priSG-A、SG-B及plpSC)主要參與調(diào)控mRNA加工、選擇性剪接或穩(wěn)定等過程,而在后期階段(例如rST及elST)主要參與調(diào)控核糖體組裝或mRNA翻譯等過程,提示這些階段特異表達的RBPs在生精細胞發(fā)育過程發(fā)揮了重要調(diào)控作用,并為精子發(fā)生中的“轉(zhuǎn)錄-翻譯”解偶聯(lián)提供了潛在分子基礎(chǔ)。此外,RBPs在細胞內(nèi)經(jīng)常以共表達形式存在并能驅(qū)動靶標mRNA的協(xié)同表達,進而調(diào)控細胞分化或組織發(fā)育。本研究發(fā)現(xiàn)RBPs在精子發(fā)生過程具有明顯的共表達特征,并主要參與mRNA代謝、mRNP組裝或翻譯等過程,提示精子發(fā)生中共表達的RBPs是操控RNA命運和功能的潛在重要參與者。然而,這些共表達的RBPs需要在細胞和動物水平進行分子生物學驗證,深入解析RBPs與RNA的相互作用機制、生化特征及生理功能可為理解、診斷和治療男性不育相關(guān)疾病提供新的線索。
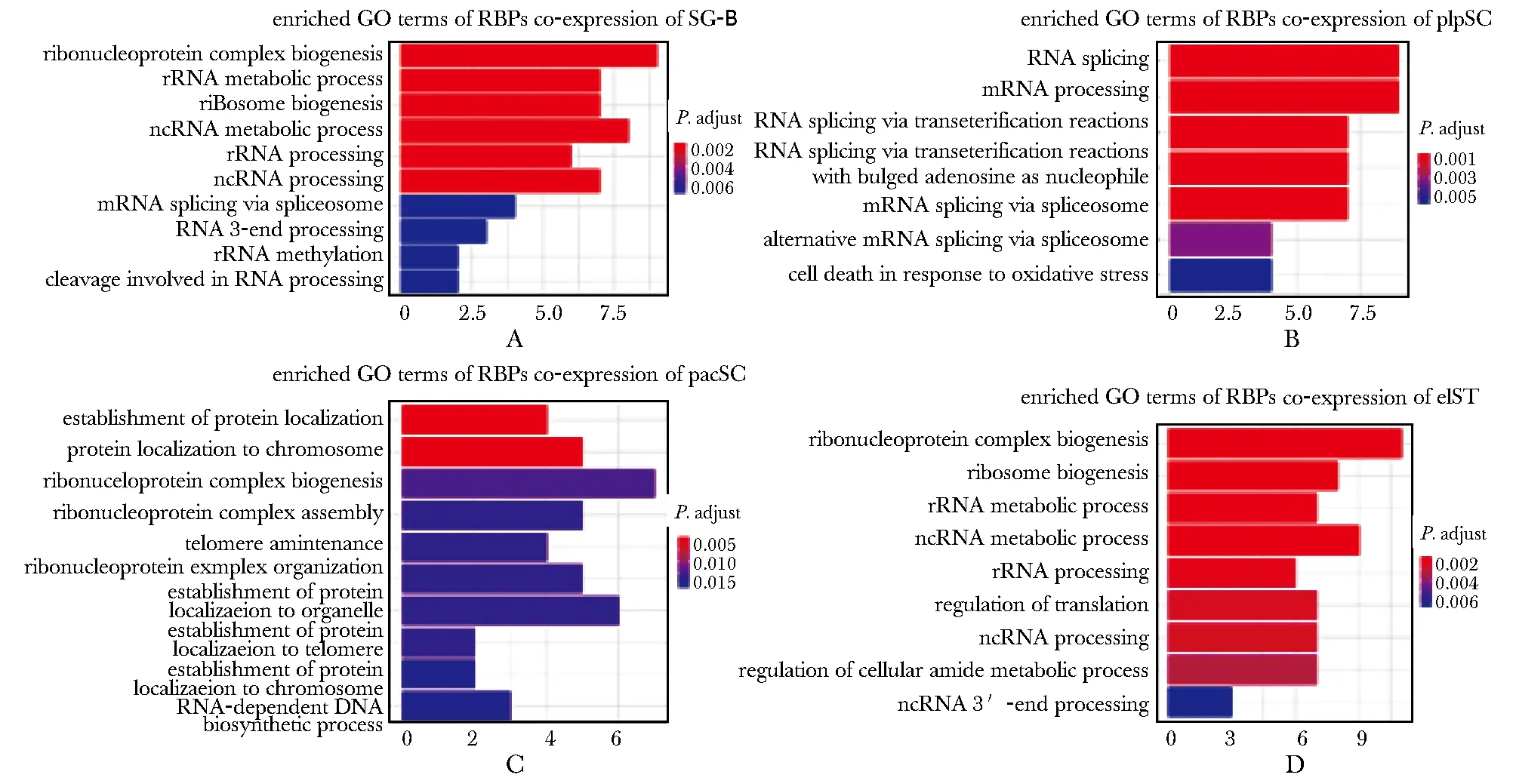
A-D.enriched GO terms of RBPs co-expression in SG-B, plpSC, pacSC, rST and elST圖4 共表達RBPs在精子發(fā)生中的GO功能富集分析Fig 4 GO enrichment analysis of RBPs co-expression in spermatogenesis
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
中國中醫(yī)藥現(xiàn)代遠程教育(2014年22期)2014-03-01 04:32:55
中國中醫(yī)藥現(xiàn)代遠程教育(2014年16期)2014-03-01 04:28:54
