NLRP3炎癥小體在抑郁癥中的作用及中藥干預研究進展
2021-06-11 08:01:48曾九僧紀雅菲胡靖文
中草藥 2021年11期
曾九僧,紀雅菲,方 洋,胡靖文,劉 蓉,曾 南
成都中醫藥大學藥學院,四川 成都 611137
抑郁癥又稱抑郁障礙,是常見的精神疾病之一,臨床主要特征為顯著持久的心境低落、興趣缺失、意志減退,全球約有3.5 億抑郁癥患者,中國終身患病率約為3.3%[1]。抑郁癥發病機制復雜,包括神經遞質及其受體紊亂、神經內分泌失調、神經營養與神經發生、免疫反應與神經炎癥、腸道菌群改變等眾多假說。尚未闡明的發病機制可能是部分患者應用臨床常規抗抑郁藥治療效果不佳甚至無效及抑郁癥易復發的原因之一[2]。中醫學中,依據抑郁癥的一些行為和癥狀,將其歸屬于百合病、郁證、臟燥等病證[3],并發現中藥治療能改善快感缺失等核心癥狀,且對并發癥亦有治療作用,安全性相對較高,因此受到國內外學者的廣泛關注。
20 世紀90年代,免疫反應和神經炎癥與抑郁癥的生物學關聯受到學者們的關注[4]。研究發現,重度抑郁癥患者可見巨噬細胞、小膠質細胞顯著激活[5],白細胞介素(interleukin,IL)-1β、腫瘤壞死因子(tumor necrosis factor,TNF)-α 等炎癥因子表達顯著升高[6],血清炎癥因子水平與抑郁癥狀程度、自殺意念呈正相關[7-8],抗炎治療則能減輕患者相關情緒癥狀[9]。固有免疫作為人體抵抗外界感染和內源性影響的第一道屏障,模式識別受體是固有免疫的重要組成部分,核苷酸結合寡聚化結構域樣受體家族 pyrin 結構域蛋白 3(nucleotide binding oligomerization domain-like receptor family pyrin domain protein 3,NLRP3)是NLR 中衍生的多域蛋白復合物,是固有免疫的重要組成因子,由22 個人類基因編碼,是固有免疫系統中加工和處理炎癥因子的主要分子平臺[10]。NLRP3 炎癥小體包括含有特征性的熱蛋白結構域作為中心結構的結構蛋白NLRP3,以及具有熱蛋白結構域、半胱氨酸蛋白水解酶(cysteine aspartate-specific protease,Caspase)募集域(Caspase activation and recruitment domain,CARD)的接頭蛋白凋亡相關斑點樣蛋白(apoptosisassociated speck-like protein,ASC)和效應蛋白Caspase-1[11-12],是迄今為止研究最多最廣泛的炎癥小體,其主要在外周巨噬細胞、單核細胞、常規樹突狀細胞和中樞小膠質細胞中表達[13]。
NLRP3 炎癥小體被激活發揮致炎作用前,首先需要接受啟動信號(信號1),即免疫細胞需要接受引發刺激,如細胞膜上的TNF 受體、IL-1 受體(IL-1R)、Toll 樣受體(Toll-like receptors,TLR)的配體經對應的受體及通路激活核因子κB(nuclear factor κB,NF-κB),活化的NF-κB 進一步上調NLRP3、pro-IL-1β的mRNA 轉錄水平,高水平的NLRP3、pro-IL-1βmRNA 是形成有效炎癥小體的關鍵[14-15]。
接受啟動信號后,NLRP3 炎癥小體會因趨化因子、三磷酸腺苷(adenosine triphosphate,ATP)、K+載體、血紅素、溶酶體損傷釋放的顆粒物[16],各種菌群/病原體釋放的有毒成分[17]、線粒體紊亂及活性氧(reactive oxygen species,ROS)等多種因素而被激活(信號2),活化狀態下的炎癥小體誘導炎癥因子的成熟與釋放并啟動下游炎癥反應。多項研究已證實NLRP3 炎癥小體的激活在抑郁癥的發生發展中具有重要作用[18-20]。本文主要通過文獻回顧,對NLRP3 炎癥小體相關通路及關鍵靶點在抑郁癥中的作用,以及中藥干預該通路治療抑郁癥的研究現狀進行歸納總結。
1 與抑郁癥關聯的NLRP3 炎癥小體相關通路
1.1 NLRP3/Caspase-1/細胞因子/神經炎癥與抑郁癥
精神疾病與固有免疫系統激活之間的關系已被廣泛討論,神經炎癥與腦功能之間的相互關系也很密切[21]。受到心理應激釋放的病原相關分子模式(pathogen-associated molecular patterns,PAMPs)或損傷相關分子模式(damage-associated molecular patterns,DAMPs)激活后,NLRP3 結構蛋白的寡聚化作用使其與接頭蛋白ASC 的熱蛋白結構域相結合,然后ASC 的CARD 與pro-Caspase-1 上的CARD結合,形成完整且有活性的NLRP3 炎性小體[22],促使pro-Caspase-1 自我裂解,產生活性的效應蛋白Caspase-1,誘導IL-1β 和IL-18 從未成熟狀態轉化為活性狀態,并分泌到胞外從而誘發神經炎癥。
小膠質細胞是腦內主要分泌IL-1β 和IL-18 的細胞類型,小膠質細胞依賴性炎癥小體激活在大腦中發揮重要作用,尤其在神經炎癥狀態下[23]。抑郁癥發病前環境應激會增加激活的NLRP3 炎癥小體的穩態濃度,NLRP3 炎癥小體的活性及其嚴密的調節很大程度上決定了小膠質細胞的形態、神經炎癥反應的強度以及暴露于應激源和開始炎癥反應之間的時間[24],NLPR3基因敲除(NLRP3-/-)[25]或Caspase-1基因敲除(CASP1-/-)[26]均能減輕小鼠因慢性應激出現的抑郁樣行為,且NLRP3-/-小鼠的小膠質細胞形態偏向監測性M2 樣狀態[27],可見其在連接應激與神經炎癥狀態之間的關鍵作用。
1991年,Smith[28]提出的巨噬細胞抑郁理論中,細胞因子首次被認為是抑郁癥的病因根源,后續不斷發現炎癥細胞因子可介導抑郁樣行為,從此細胞因子在抑郁癥中的作用獲得進一步重視[29]。臨床發現抑郁癥患者腦脊液[30]和血清[31]中炎癥因子水平升高,外周單核巨噬細胞[32]中NLRP3、Caspase-1 轉錄水平均升高。IL-1β 或IL-18 通過自分泌或旁分泌機制所誘導的其他炎性細胞因子如IL-6 和TNF-α,在抑郁癥患者的血清中也持續增加[33-34]。抑郁癥患者血清IL-1β、IL-6、IL-8、IL-12 和TNF-α 水平升高,同時IL-10 水平下降[35];抑郁癥癥狀的緩解與血漿細胞因子水平的正常化有關[36]。有研究認為,細胞因子可被用作內源性生物標志物,以監測抗抑郁藥的療效[37]。綜上所述,由危險信號誘發的神經炎癥反應是抑郁癥發病機制中的重要環節,其中NLRP3 炎癥小體在神經炎癥反應發生中發揮著關鍵介導作用。
1.2 ATP/P2X 配體門控離子通道7 嘌呤能受體(P2X ligand-gated ion channel 7 purinergic receptor,P2X7R)/NLRP3 通路與抑郁癥
P2X 受體家族是ATP 門控陽離子通道,哺乳動物中迄今發現了7 個亞型,其中P2X7 亞型是目前已知與抑郁癥關系最為緊密的亞型之一。暴露于應激壓力后,神經元或星形膠質細胞釋放的高濃度ATP 能激活小膠質細胞中的P2X7R;P2X7R 激活引起K+外排,胞質中K+外排可引起NIMA 相關蛋白激酶7 與NLRP3 蛋白結合導致炎癥小體的組裝和激活,最終誘導炎癥細胞因子分泌[38]。ATP 在小膠質細胞NRLP3 激活中的作用可被氯化鉀預處理阻斷,表明細胞內環境中K+外流是NLRP3 炎癥小體激活的必需條件[39]。P2X7R 和NLRP3 在小膠質細胞中大量共表達是ATP 影響NLRP3 炎癥小體的結構特征[40]:ATP 刺激培養的小膠質細胞,可增強NLRP3、ASC 蛋白和Caspase-1 的表達,并呈NLRP3炎癥小體相關性IL-1β、IL-18 分泌增加;但對于培養的星形膠質細胞則未觀察到NLRP3 炎癥成分(ASC、Caspase-1 和細胞因子)表達,表明NLRP3表達與細胞類型的特異性有關[41]。
動物實驗研究亦證實了ATP 通過P2X7R 介導NLRP3 炎癥小體的激活。大鼠急性束縛應激期間,海馬體中細胞外ATP 大量增加,誘導NLRP3 炎癥小體激活引起細胞因子IL-1β、TNF-α 釋放增加,選擇性P2X7R 拮抗劑A804598 則阻斷上述表現,并逆轉抑郁樣行為[42]。慢性不可預見性溫和應激(chronic unpredictable mild stress,CUMS)大鼠模型中,海馬細胞外ATP 水平與CUMS 刺激時長呈正相關[43]。P2X7R 拮抗劑亮藍G 和A438079 能預防大鼠的抑郁樣行為[44];連續7 周ip 亮藍G 或氟西汀,可抑制CUMS 模型小鼠皮層、海馬和基底核部位小膠質細胞的活化,抑制P2X7R 的表達及調節神經內分泌,逆轉抑郁樣行為[45];P2X7R基因敲除(P2X7R-/-)小鼠對CUS 誘導的抑郁樣行為效應表現出較弱反應[43],且基因敲除小鼠在脂多糖(lipopolysaccharide,LPS)刺激后其抑郁樣行為表現較少,與其不能釋放IL-1β 有關[46]。綜上所述,P2X7R作為ATP 門控陽離子通道,能通過調節ATP/P2X7R/ NLRP3 軸的活性來影響抑郁樣癥狀的發生,是與炎癥小體密切相關的潛在抗抑郁作用的重要靶標。
1.3 線粒體紊亂/ROS/氧硫蛋白相互作用蛋白(thioredoxin-interacting protein,TXNIP)/NLRP3/ 細胞焦亡與抑郁癥
抑郁癥帶來的心理應激會使腦區細胞耗氧量增加,ROS 主要是由線粒體氧化磷酸化產生的不穩定、高活性分子,ROS 同樣被認為是NLRP3 炎癥小體激活的觸發因子之一[47]。體外實驗證實,線粒體功能紊亂所產生的ROS 介導了由LPS 和ATP 引起的炎癥小體激活,線粒體DNA(mitochondrial DNA,mtDNA)作為連接線粒體功能障礙與Caspase-1 激活的信使分子[48],被釋放到胞質中的氧化mtDNA 對于NLRP3 炎癥小體的激活亦是必要的[49]。
ROS 誘導TXNIP 與NLRP3 結構蛋白結合可活化炎癥小體[50],NLRP3 炎癥小體激活則可誘發免疫細胞呈現Caspase-1 相關性的細胞焦亡[51]:活性Caspase-1 裂解gasdermin 家族蛋白D(gasdermin D,GSDMD),gasdermin D 引起小膠質細胞腫脹、溶解,致使促炎細胞因子IL-1β 和IL-18 釋放到腦區并延長免疫應答,加重抑郁癥患者的神經炎癥狀態[52]。
臨床發現,抑郁癥患者外周單核細胞中線粒體的ROS 水平顯著升高,并伴隨NLRP3 炎癥小體的激活[32]。研究表明,CUMS 模型大鼠海馬中ROS 與TXNIP 蛋白水平顯著升高,同時炎癥小體被激活[53];小鼠ip LPS誘導的急性抑郁樣模型中亦觀察到同樣結果[54];用 LPS +MSU 刺激解偶聯蛋白 2(uncoupling protein 2,UCP2,線粒體內膜上能控制ROS 產生的蛋白質)基因敲除小鼠(UCP2-/-)的原代星形膠質細胞和野生型小鼠原代星形膠質細胞,發現 UCP2-/-小鼠原代星形膠質細胞的 ROSTXNIP-NLRP3 通路激活處于更高水平[55]。綜上所述,ROS 在NLRP3 炎癥小體激活中發揮重要作用,線粒體功能紊亂及其ROS 的過度產生能特異性地致使NLRP3 炎癥小體激活和細胞焦亡,通過觀察NLRP3 與線粒體的相互作用將有助于理解應激所致神經炎癥反應的機制。
1.4 NLRP3/下丘腦-垂體-腎上腺軸(hypothalamic- pituitary-adrenal,HPA)軸與抑郁癥
細胞因子是HPA 軸的強刺激物[56],由于免疫系統的激活與炎癥反應,可見抑郁癥患者血漿細胞因子處于高水平狀態,而NLRP3 炎癥小體激活所分泌的細胞因子亦參與到神經內分泌功能調控中。研究表明,CUMS 模型小鼠物血清糖皮質激素的水平隨著NLRP3 炎癥小體的激活而升高[57];IL-1R 敲除小鼠(IL-1R-/-)及雙側腎上腺切除的小鼠在CUMS 程序刺激下無抑郁樣行為表現,且皮質酮水平處于正常狀態[58]。且發現抗抑郁藥的治療抗性與HPA軸功能障礙有關,但其潛在機制有待深入了解[59]。
1.5 腸道菌群/NLRP3 與抑郁癥
腸道菌群在影響行為,及其與大腦的相互聯系,可能潛在地參與了抑郁癥病理生理過程 ,已成為近年來新興的抑郁癥假說之一。微生物-炎癥小體-腦軸被認為是雙向交流系統,可將心理應激反應、免疫系統功能和腸道微生物組聯系起來。一方面,心理應激將導致NLRP3 炎癥小體激活,產生的IL-1β 和IL-18 經血漿對腸壁產生刺激作用,應激介導的腸屏障功能改變導致菌群移位,促進了炎癥信號的傳遞[60];另一方面,應激誘導的炎癥反應可介導腸道菌群變化及其代謝產物的紊亂,加強全身和中樞NLRP3 介導的促炎信號傳導,進一步影響腦功能和加重抑郁樣行為[61]。
動物實驗研究表明,中樞神經炎癥信號與腸道菌群之間關系密切。多種動物模型中發現炎癥信號的激活能影響腸道菌群的改變[62];將NLRP3-/-的腸道菌群移植到野生型小鼠腸道中,可有效抑制CUMS 誘導的抑郁樣行為,并減輕大腦功能障礙[63];CASP1-/-或Caspase-1 抑制劑米諾環素,能通過調節壓力與腸道菌群組成之間的關系來改善慢性束縛應激引起的小鼠抑郁樣行為[64]。綜上所述,腸道菌群與炎癥小體相關性強,但NLRP3 介導的通路是如何參與到腸道菌群-中樞功能通路中的潛在因果關系尚需進一步研究。
綜上所述,NLRP3 炎癥小體及其相關通路與抑郁癥聯系密切,涉及到神經炎癥、氧化應激、神經內分泌、腸道菌群等眾多抑郁癥發病機制假說。深入研究NLRP3 炎癥小體及其相關通路,不僅有助于深化抑郁癥發病機制中的應激信號轉化、免疫調節與炎癥反應表現等科學認識,而且能為抑郁癥治療切入點及創新藥物研發找到突破口。NLRP3 炎癥小體及其相關通路與抑郁癥的關系示意圖見圖1。
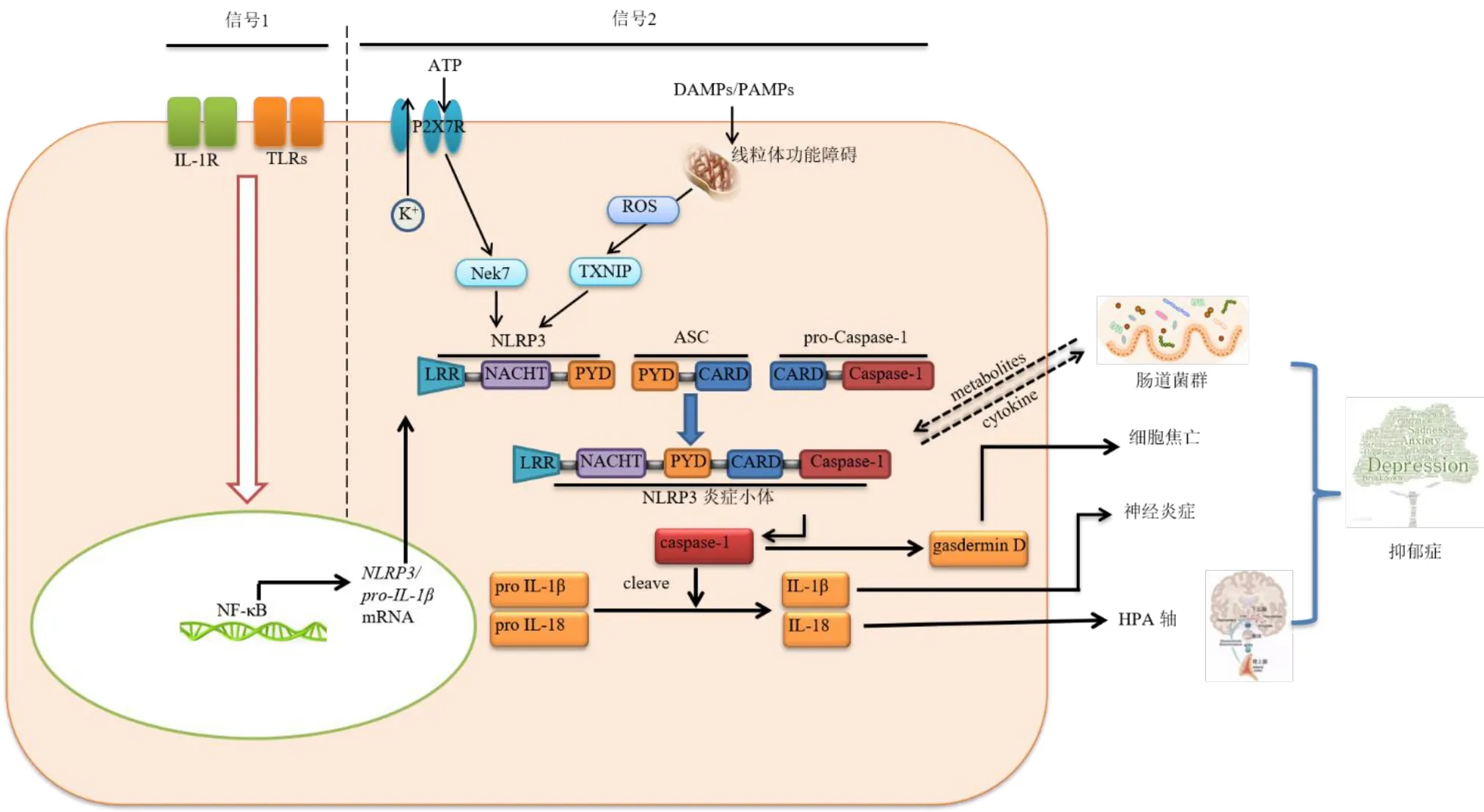
圖1 NLRP3 炎癥小體及其相關通路與抑郁癥的關系示意圖Fig.1 Relation diagram of NLRP3 inflammasome and related pathways in depression
2 中藥干預NLRP3 炎癥小體的抗抑郁作用
基于NLRP3 炎癥小體相關通路與抑郁癥發生關聯的研究現狀,本文主要綜述了多種中藥復方、中藥提取物及中藥單體成分通過抑制NLRP3 炎癥小體激活及減輕中樞神經炎癥反應的抗抑郁作用。
2.1 中藥復方
中藥復方是傳統中藥臨床應用最多的形式,目前已證實一些中藥復方能通過影響NLRP3 炎癥小體及相關蛋白表達來發揮抗抑郁作用。研究表明,對CUMS模型大鼠ig 柴胡疏肝散0.925、1.85 g/kg[65]及ig 半夏厚樸湯3.29、6.58 g/kg[66]均能不同程度降低血清中IL-1β 水平,抑制腦區NLRP3 炎癥小體的激活,并改善模型大鼠外周胰島素信號傳導。對CUMS 模型大鼠ig 黃連溫膽湯5.69、11.35 g/kg 抗抑郁作用的發揮與抑制大鼠海馬腦區NF-κB 蛋白表達,抑制前額葉皮質NLRP3 炎癥小體激活及減少炎癥因子的釋放等有關[67]。麻黃附子細辛湯12.5 g/kg 對LPS 誘導的小鼠有拮抗抑郁樣行為的作用,與其抑制海馬TXNIP 的增加、NLRP3 炎癥小體的激活,及上調腦源性神經營養因子(brain-derived neurotrophic factor,BDNF)、酪氨酸激酶受體B(tyrosine kinase receptor B,TrkB)表達增強神經發生等有關[68]。
2.2 中藥提取物
中藥提取物通常指用特定方法提取獲得的具有多種藥理活性物質組成的中藥有效部位,保留了中藥整體性和多成分的特點。目前中藥提取物通過抑制NLRP3/Caspase-1/IL-1β 軸緩解抑郁樣行為的研究報道相對較少。研究發現,銀杏酮酯50、100 mg/kg 能顯著改善持續注射LPS 誘導的抑郁樣行為,與抑制海馬NLRP3/Caspase-1/IL-1β 通路有關,能抑制NLRP3 炎癥小體的激活及減少炎癥因子的分泌[69]。金銀花多糖100 mg/kg 能顯著抑制CUMS模型小鼠海馬中NLRP3、Caspase-1 和IL-1β 的表達上調,從而減弱模型小鼠的抑郁樣行為[70]。
2.3 中藥單體成分
單體成分是中藥發揮功效的直接物質基礎,其抗抑郁作用的研究報道最為豐富,目前發現很多中藥單體成分能通過影響與NLRP3 炎癥小體相關的神經炎癥、氧化應激、神經發生等信號通路的激活,產生抗抑郁作用。
基于經典的CUMS 抑郁樣動物模型,研究發現,萜類化合物左旋薄荷酮15、30 mg/kg[71]和百里香酚15、30 mg/kg[72]均能逆轉CUMS 誘導的小鼠抑郁樣行為,上調海馬單胺類神經遞質水平,且抑制NLRP3 炎癥小體活化從而降低Caspase-1 表達及IL-1β、TNF-α 等促炎細胞因子的水平;阿魏酸40、80 mg/kg[73]及人參皂苷Rg120、40 mg/kg[74]能顯著抑制 CUMS 誘導的小膠質細胞活化及 NFκB/NLRP3 炎癥信號的激活;青藤堿30、100、300 mg/kg 能通過抑制p38 絲裂原活化蛋白激酶(p38 mitogen-activated protein kinase,p38 MAPK)/NF-κB通路和NLRP3 炎癥小體的激活,降低腦區IL-1β、IL-6、TNF-α 等多種炎癥因子的表達來減少CUMS小鼠的抑郁樣行為發生[75];黃芩苷20、40 mg/kg 能抑制模型大鼠皮層糖原合成酶激酶3β(glycogen synthase kinase 3β,GSK3β)、NF-κB磷酸化及NLRP3炎癥小體的激活[76],降低前額葉皮層Caspase-1(p20)和促炎細胞因子(IL-1β、IL-6)的水平,保護受到神經炎癥影響的神經元細胞從而發揮抗抑郁作用[77];穿心蓮內酯2.5、5 mg/kg 能以促進自噬的方式抑制CUMS 小鼠前額葉皮質NF-κB 信號通路和NLRP3炎癥小體(NLRP3、ASC 和Caspase-1)激活,降低前額葉皮質促炎介質和細胞因子水平[78];遠志皂苷元4、8 mg/kg 能下調NF-κB/NLRP3 炎癥途徑相關蛋白激活,抑制IL-1β 分泌,并提高BDNF 和神經營養因子3(neurotrophin-3,NT-3)的蛋白表達水平來顯著改善CUMS 小鼠的抑郁樣行為[79];五味子甲素20、40 mg/kg 能抑制Toll 樣受體4(Toll-like receptor 4,TLR4)/NF-κB/NLRP3 神經炎癥通路和促進谷氨酸受體(glutamate receptors,GluR)/突觸后致密蛋白95(postsynaptic density protein 95,PSD95)級聯反應,改善樹突棘密度,發揮對CUMS模型小鼠的抗抑郁作用[80];水飛薊素200 mg/kg 或水飛薊素納米顆粒10 mg/kg 能通過促進神經發生,提高神經遞質水平,抑制前額葉皮質和海馬NLRP3炎癥小體活化導致的神經炎癥而產生抗抑郁作用[81];紫蘇醛20、40 mg/kg 能不同程度抑制CUMS 模型大鼠海馬中TXNIP、NLRP3、Caspase-1 和p65 的蛋白表達上調,調控TXNIP/TRX/NLRP3 通路減輕CUMS 誘導的大鼠抑郁樣行為[53];丹酚酸B 20、40 mg/kg 能以抑制氧化應激和減輕神經炎癥反應的方式逆轉CUMS 模型大鼠的抑郁樣行為[82];姜黃素100 mg/kg 能通過抑制應激誘導的P2X7R/NLRP3 軸活化,減少IL-1β 的分泌來緩解CUMS 模型動物的抑郁樣行為,并能抑制NF-κB 的活化,降低吲哚胺- 2,3-二加氧酶的活化和犬尿氨酸/色氨酸比值的增加發揮神經保護作用[83]。
基于LPS 誘導的行為異常抑郁樣動物模型,研究表明,西紅花苷20、40 mg/kg 能抑制LPS 誘導的小鼠海馬NLRP3 炎癥小體活化和Caspase-1 的表達升高,降低海馬中多種炎癥因子水平來發揮抗抑郁樣行為作用,西紅花苷5~20 μmol/L 亦能抑制LPS誘導的小膠質BV-2 細胞炎性標記物和ROS 的產生,顯著下調誘導型一氧化氮合酶(inducible nitric oxide synthase,iNOS)、NF-κB p65 的表達,并促進小膠質細胞從M1 向M2 表型轉化[84];丹酚酸B 20 mg/kg 能以促進自噬的方式抑制NLRP3 炎癥小體的激活和促炎細胞因子的表達,有效緩解持續注射LPS 誘導的大鼠抑郁樣行為[85]。
基于反復束縛應激(repeated restraint stress,RRS)/LPS 刺激小鼠抑郁模型,黃芪甲苷IV 20、40 mg/kg 能增加過氧化物酶體增殖物激活受體γ(peroxisome proliferators-activated receptor γ,PPARγ)表達和GSK3β 磷酸化,降低海馬NF-κB 磷酸化、NLRP3 炎癥小體的激活,及Caspase-1(p20)、炎癥因子(IL-1β、IL-6)的水平,發揮抗抑郁作用[86]。
基于社會失敗抑郁模型(chronic social defeat stress,CSDS)小鼠,大蒜素50 mg/kg 能下調模型小鼠海馬中小膠質細胞的活化及炎性細胞因子的升高,抑制ROS 產生,并上調SOD 和核因子E2 相關因子2(nuclear factor erythroid 2-related factor 2,Nrf2)/血紅素加氧酶1(heme oxygenase 1,HO-1)通路的活性,同時抑制NLRP3 炎性小體相關蛋白(ASC、Caspase-1 和IL-1β)的表達[87],發揮抗抑郁作用。
綜上所述,目前通過干預NLRP3 炎癥小體及其相關通路而發揮抗抑郁作用的中藥見表1。
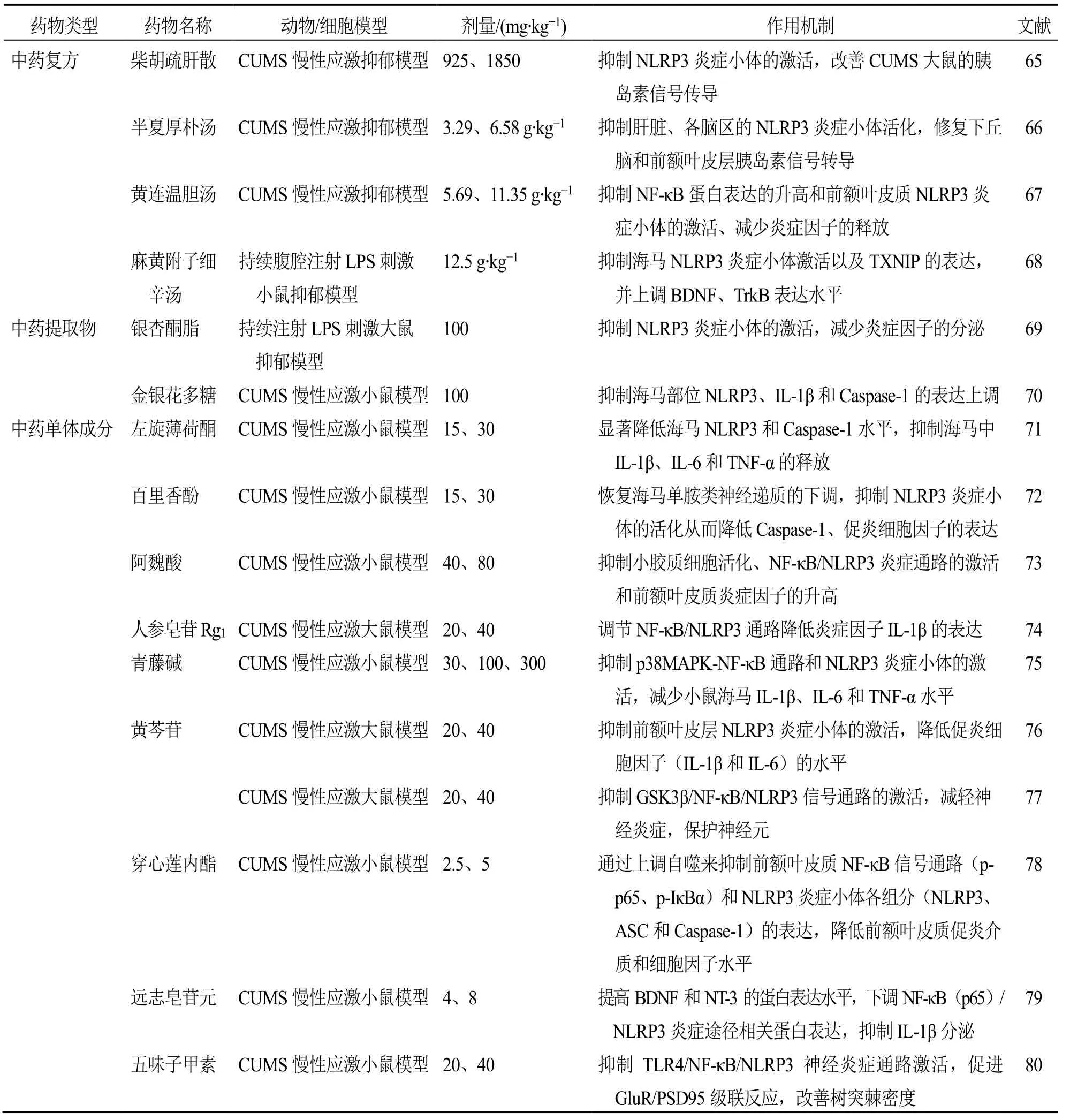
表1 中藥對NLRP3 炎癥小體及相關途徑的抗抑郁作用Table 1 Antidepressant effects of traditional Chinese medicine on NLRP3 inflammasome and related pathways
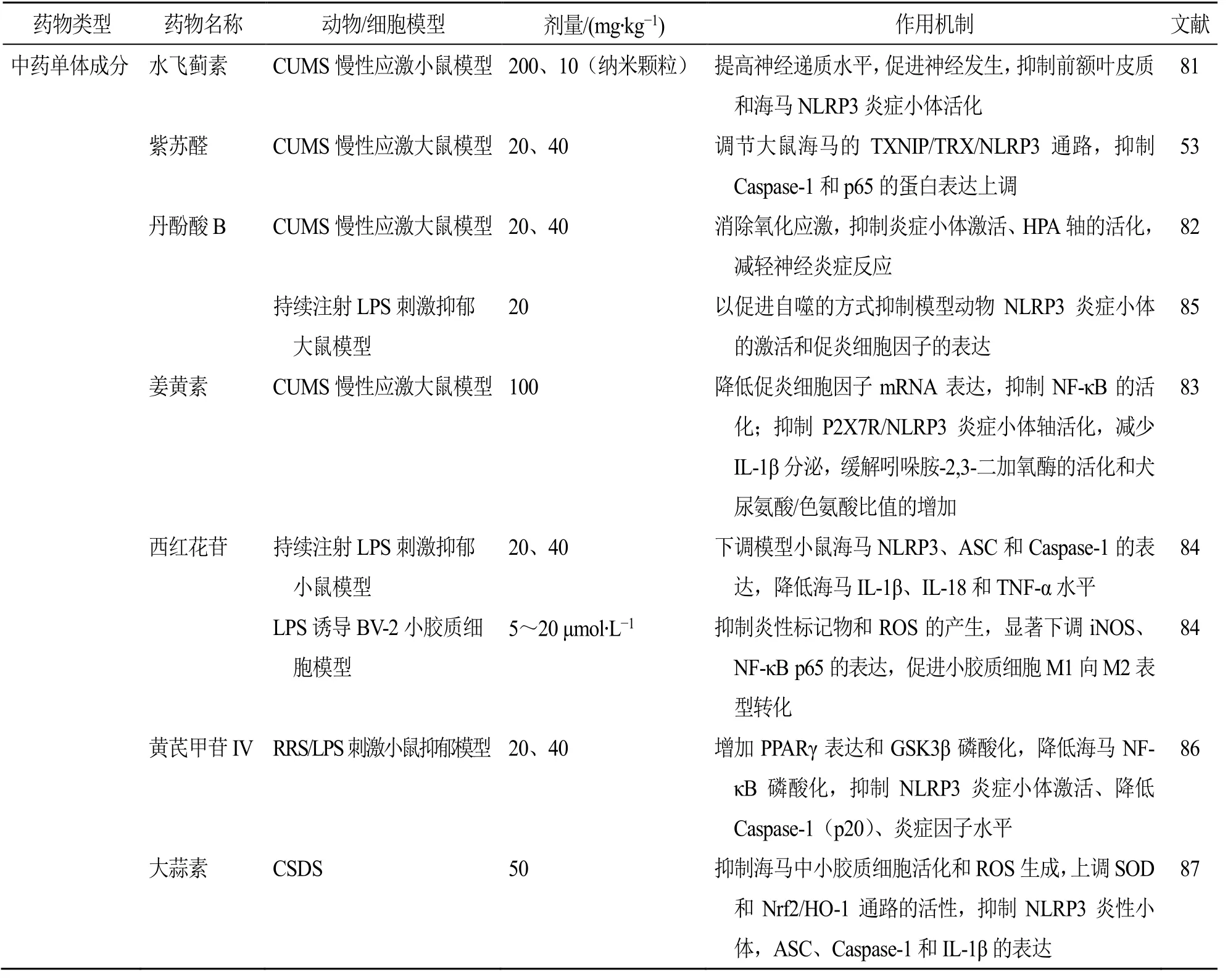
續表1
3 結語與展望
NLRP3 炎癥小體作為模式識別受體的重要組成部分之一,不僅是固有免疫系統中感應心理壓力應激的關鍵節點,而且是啟動下游炎癥級聯反應的重要環節。眾多臨床前與臨床研究均表明,炎癥小體介導了不同抑郁樣動物模型外周和中樞炎癥反應及抑郁樣行為表現[57,88];抑郁癥患者呈現免疫細胞中炎癥小體的激活[32]和血清高水平炎癥因子表達[89]的病理表現,降低炎癥因子水平有助于患者抗抑郁治療療效提高[90]。因此,抑郁癥的發生發展病理機制與炎癥、免疫反應的過度密切相關,NLRP3炎癥小體則代表了抑郁癥中壓力與炎癥傳感器的功能,為抑郁癥的免疫炎性假說提供了重要依據。
目前,關于中藥抗抑郁癥作用機制的研究已表明,通過影響啟動信號[73]或嘌呤能受體[83]、氧化應激[53,87]從而抑制NLRP3 炎癥小體激活、降低炎癥因子水平來減輕神經炎癥反應發揮抗抑郁作用是許多中藥的作用機制表現之一,同時通過抑制HPA 軸的活化[82]、上調神經營養因子水平[79]、促進神經發生[80]、改善胰島素信號[65-66]等其他途徑來調控抑郁癥所致的機體穩態失調亦是中藥的作用機制表現,呈現出多靶點特點,展現了中藥活性成分的多樣性,而其多靶點的藥理作用表現與抑郁癥發病及治療多靶標表現之間呈現良好契合度。此外,臨床研究表明,口服加味甘麥大棗湯4 周能降低圍絕經期伴初發重度抑郁患者漢密爾頓抑郁量表、Kupperman 量表評分,同時下調患者血清IL-1β、IL-6、TNF-α 等炎癥因子的水平[91];選擇性5-羥色胺再攝取抑制劑聯合甘草酸(干預組)治療與選擇性5-羥色胺再攝取抑制劑藥物聯合安慰劑(對照組)治療4 周后,干預組患者血清TNF-α 和IL-1β 的累積降低值更高,聯合中藥抗炎治療能產生更佳療效[92],足見中藥在基于免疫炎癥假說治療抑郁癥方面擁有巨大潛力。
盡管NLRP3 炎癥小體是抑郁癥免疫炎癥假說中研究最為深入的模式識別受體,但隨著對該假說研究的深入,包括TLR4、晚期糖基化終產物受體等模式識別受體與抑郁癥的關系也不斷明確[93],如作為啟動信號的TLR4 與NLRP3 在某些抑郁癥動物模型中呈現一致的高表達[94],但針對這些模式識別受體(pattern recognition receptors,PPRs)及其配體與抑郁癥的研究則主要集中在發病機制或與其他通路交互關系研究方面,而抗抑郁中藥能否靶向上述PPRs 的機制研究報道則較少,僅有黃芩苷能通過磷脂酰肌醇激酶/蛋白激酶B/插頭箱轉錄因子O1途徑抑制TLR4 表達,從而改善CUMS 及連續腹腔注射LPS 誘導的動物抑郁樣行為的報道[95]。因此,基于抑郁癥免疫炎癥假說開展中藥抗抑郁作用的研究,筆者認為尚可從以下路徑進行完善、深入:(1)神經炎癥方面,抗抑郁中藥(復方、提取物及有效成分)的研究可著重于NLRP3 炎癥小體與其他PPRs 的協同/拮抗性關系研究,以闡釋其相互之間的關聯性,有利于中藥整體性的表征;(2)探究中藥對固有免疫系統相關的上、下游通路及通路中不同關鍵因子的影響,并從基因組學、代謝組學等角度歸納、整合抗抑郁中藥對與NLRP3 炎癥小體相關的不同靶標的影響,尋找有效的靶標;(3)基于抑郁癥免疫炎癥假說,開展中藥臨床治療抑郁癥的研究,為該假說和抑郁癥的中藥治療提供更充分的科學依據。
綜上所述,免疫炎癥假說作為抑郁癥發病機制中的重要假說之一,結合抑郁癥患者伴有中樞或外周高水平炎癥因子的病理變化共識,且NLRP3 炎癥小體與上述假說及病理表現密不可分,由此,充分理解NLRP3 炎癥小體及其相關通路在抑郁癥發生發展中的作用,結合網絡藥理學、分子生物學等研究手段開展中藥抗抑郁作用機制及物質基礎的研究,將有助于為中醫藥現代化、為廣大抑郁癥患者提供更安全有效的治療選擇提供強有力的支撐,為人類健康服務。
利益沖突所有作者均聲明不存在利益沖突
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中老年保健(2021年4期)2021-12-01 11:19:40
中老年保健(2021年4期)2021-08-22 07:08:32
學苑創造·A版(2020年9期)2020-10-13 09:41:02
金橋(2020年7期)2020-08-13 03:07:00
基層中醫藥(2020年12期)2020-07-22 06:34:38
基層中醫藥(2018年6期)2018-08-29 01:20:20
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
肝博士(2015年2期)2015-02-27 10:49:49
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
