角倍蚜各蟲態蠟腺的分布與結構
2021-05-31 01:53:42魏洪媛豐國蕊邵淑霞陳曉鳴楊子祥
昆蟲學報 2021年4期
魏洪媛, 豐國蕊, 許 鑫, 邵淑霞, 陳曉鳴, 楊子祥
(中國林業科學研究院資源昆蟲研究所, 國家林業和草原局資源昆蟲培育與利用重點實驗室, 昆明 650224)
角倍蚜Schlechtendaliachinensis屬半翅目(Hemiptera)蚜科(Aphididae)癭綿蚜亞科(Eriosomatinae)五節根蚜族(Fordini)倍蚜屬Schlechtendalia(張廣學和陳小琳, 1999; Blackman and Eastop, 2007),是五倍子的主要生產種,其在鹽膚木Rhuschinensis上寄生形成的蟲癭稱為角倍,約占五倍子總產量的70%,具有重要的經濟價值(張宗和, 1991; 楊子祥, 2011)。角倍蚜具有多種蟲型和復雜的生活史,包括有性生殖與孤雌生殖的繁殖方式及兩種寄主植物的轉主交替寄生。每年3月初,越冬若蚜在側枝匐燈蘚Plagiomniummaximoviczii等冬寄主上羽化為春遷蚜,遷飛到附近的鹽膚木樹干上,產雌雄性蚜,交配后的雌性蚜產干母,1齡干母沿著樹干爬到梢頭嫩葉上取食,刺激葉片細胞增生包裹自身形成蟲癭;4~5 d后干母在蟲癭內蛻皮,變成2齡干母,繼續發育并經歷3次蛻皮后發育成4齡成蚜。于6月中下旬產生干雌,蟲癭在干母和干雌的取食刺激下繼續生長;秋季,干雌發育成有翅秋遷蚜,蟲癭爆裂后遷飛到附近的冬寄主蘚上產若蚜越冬;次年春季,越冬若蚜發育成有翅春遷蚜,開始新的生活史循環(楊子祥, 2011; 邵淑霞等, 2012)。角倍蚜各蟲態均分泌蠟粉,以適合復雜多變的生活微環境,其中干雌和越冬若蚜分泌的蠟粉最多,蠟粉具有防水、保溫和防止病原菌寄生等功能(Weietal., 2020)。
昆蟲綱中半翅目、鱗翅目、膜翅目和鞘翅目等的部分種類也分泌蠟粉,其主要作用是在蟲體或卵的表面形成保護層,保護昆蟲免受雨水、病原微生物和天敵等危害(Chen and Qiao, 2012; 李嘉樂等, 2018)。蠟粉變化最豐富的是半翅目的蚜總科、蚧總科和蠟蟬總科,不同種類的蠟腺分布、發育程度、形狀和結構等各不相同(Pope, 1983; Andrea and Emanuele, 2004; Qietal., 2019);同一種類蠟腺的結構和分布也隨著生存環境的不同而存在差異,如蘋果綿蚜Eriosomalanigerum的蠟粉分泌量隨著齡期的增加而增多,其寄生天敵的發生率隨著齡期增加而降低(Suetal., 2016)。
前人對角倍蚜的研究主要包括角倍蚜的生物學特性、生活史及其與寄主植物的相互作用等,但關于角倍蚜蠟腺的研究較少。本研究利用光學和電子顯微技術對角倍蚜各蟲態蠟腺的分布和超微結構進行觀察,分析各蟲態蠟腺的差異,探討蠟腺在角倍蚜生態環境適應中的功能,為角倍蚜基礎生物學研究和人工培育技術的改進提供依據。
1 材料與方法
1.1 試蟲
角倍蚜于2019年采集自云南省鹽津縣五倍子蚜培育基地(28°06′N, 104°22′E, 海拔980 m), 10月初從成熟爆裂的蟲癭中采集有翅秋遷蚜,轉移到側枝匐燈蘚蘚圃上培育,讓秋遷蚜在蘚枝上產下若蚜;次年2-3月從蘚層中收集越冬若蚜和有翅春遷蚜,并將春遷蚜置于室內培養,獲得雌雄性蚜,待雌雄性蚜交配后,收集產下的1齡干母。讓干母在鹽膚木葉片上取食形成蟲癭,收集癭內的各齡干母和干雌。
1.2 光學顯微鏡觀察
采用Keyence VHX-1000數碼顯微系統或Olympus SZX16體式顯微鏡觀察癭內外各蟲態的蠟粉、蘚枝上蠟球的結構和自然形態等,并拍照。
將各蟲態角倍蚜(每個蟲態15頭),置于2.5%戊二醛磷酸緩沖液(0.1 mol/L, pH 7.2)中固定12 h,轉至1%的四氧化鋨中固定2 h,經丙酮分級脫水后,包埋在環氧樹脂Araldite中;用Leica EM UC6超薄切片機制作成1 μm的半薄切片,用甲基苯胺藍和勞氏紫Thionin染色,采用Olympus BX-51顯微鏡觀察蠟腺結構并拍照。
1.3 透射電子顯微鏡觀察
將各蟲態角倍蚜(每個蟲態15頭)頭部切下,取蟲體部分置于2.5%戊二醛和磷酸鈉緩沖液中固定12 h,轉移至1%四氧化鋨冷溶液中固定12 h,經丙酮分級脫水后,包埋于環氧樹脂Araldite中;用Leica EM UC6超薄切片機制作成70 nm的切片,用檸檬酸鉛和醋酸鈾酰雙重染色,采用Zeiss EM900透射電子顯微鏡觀察蠟腺結構并拍照。
1.4 掃描電子顯微鏡觀察
在Olympus SZ61體視顯微鏡下對各蟲態蚜蟲表面的附著物進行清理,將清理干凈的蚜蟲分為2組,每組15頭。 第1組直接置于Hitachi TM3000掃描電子顯微鏡下,觀察蟲體上蠟腺的數量和排列方式;第2組用70%, 80%, 90%, 95%和100%的乙醇依次進行脫水,濾紙吸干;用導電膠將蟲體粘在樣品臺上,用Hitachi JS-1600離子濺射儀噴金,置于Hitachi TM3000電子顯微鏡下觀察蠟腺的結構并拍照。
2 結果
2.1 角倍蚜各蟲態蠟腺的數量和分布
角倍蚜所有6種蟲態(1齡干母除外)均有蠟腺,蠟腺縱向排列于蟲體背部,沿背部中線左右對稱分布,其中頭部2列2排、胸部4列1排、腹部6列8排(圖1: A, B, C),共56個蠟腺。干母比較特殊,1齡干母沒有蠟腺(圖2: A),2齡干母開始具有蠟腺,在腹部背面沿邊緣排列,有2列8排,共16個蠟腺,頭部和胸部背面沒有蠟腺(圖2: B)。
2.2 角倍蚜各蟲態蠟腺的結構
進一步觀察蠟腺的顯微結構,發現每個蠟腺由數量不等(2~22個)的多邊形凹陷組成,每個蠟腺附近通常伴生有1~2根感覺毛(圖2: B, C, D)。在所有6個蟲態中,干雌、春遷蚜和秋遷蚜的蠟腺復雜,每個蠟腺由10~14個凹陷組成,且蟲體各部位的蠟腺結構沒有明顯差異(圖2: C, D);越冬若蚜蠟腺的復雜程度次之,每個蠟腺由7~10個凹陷組成,蟲體各部位的蠟腺結構沒有明顯差異(圖2: E);性蚜各部位的蠟腺結構差異顯著,靠近背板中線的脊柱板和胸膜板的蠟腺結構簡單,由3~5個凹陷組成,而靠近邊緣線上的蠟腺結構復雜,由10~22個凹陷組成(圖2: F);1齡干母沒有蠟腺(圖2: A),2齡干母蟲體邊緣有蠟腺,由10多個凹陷組成,但蠟腺較其他蟲態小(圖2: B)。
2.3 角倍蚜蠟腺細胞的超微結構
超薄切片顯示,組成蠟腺的每個凹陷下對應一個分泌細胞,由直接特化的表皮覆蓋,沒有導管或者導管細胞。分泌細胞的頂端分化出微絨毛,在微絨毛與特化的表皮之間形成儲液囊,蠟腺細胞間通過間隙連接(圖3: A)。分泌細胞中含有大量的線粒體與粗面內質網,頂端微絨毛里含有大量的光面內質網(圖3: B, C)。

圖1 角倍蚜蠟腺的分布Fig. 1 The distribution of wax glands in the horned gall aphid, Schlechtendalia chinensisA: 角倍蚜蠟腺分布圖(蟲體圖仿張宗和, 1991)Wax gland distribution in the horned gall aphids (Aphid body diagram was imitated from Zhang, 1991); B: 蟲體背面的蠟腺縱切圖Wax glands of dorsal line by longitudinal section; C: 蟲體側面橫切圖Wax gland of lateral line by transverse section; D: 單個蠟腺的半薄切片縱切圖Single wax gland by semithin longitudinal section; E: 單個蠟腺的半薄切片橫切圖Single wax gland by semithin transverse section. mv: 微絨毛Microvilli; pd: 多邊形凹陷Polygonal depression; c: 單個蠟腺細胞Single wax gland cell.箭頭指蠟腺Arrows indicate wax glands.
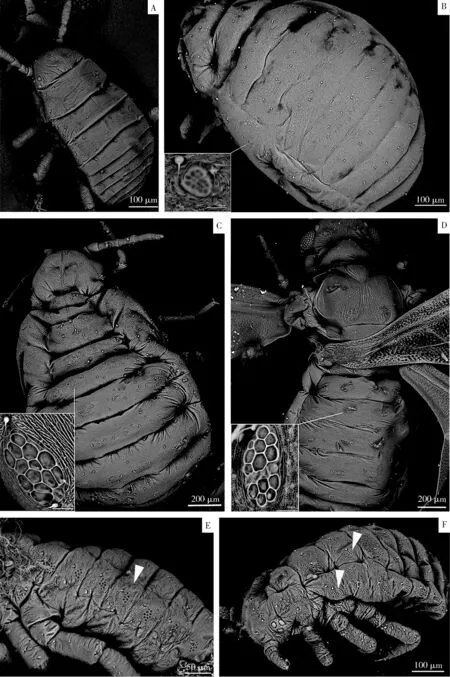
圖2 掃描電鏡觀察的角倍蚜各蟲態蠟腺的形態和結構Fig. 2 Morphology and structure of wax glands in various body forms of the horned gall aphid,Schlechtendalia chinensis observed using scanning electron microscopeA: 1齡干母1st instar fundatrix; B: 2齡干母2nd instar fundatrix; C: 干雌Fundatrigeniae; D: 秋遷蚜Fundatrispuriae; E: 越冬若蚜Apterae; F: 雄性蚜Sexual male. 箭頭示蠟腺。Arrows indicate the wax glands.
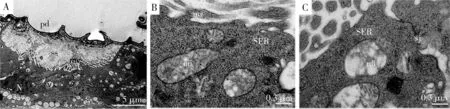
圖3 透射電鏡觀察的角倍蚜蠟腺的超微結構Fig. 3 Ultrastructure of wax glands of the horned gall aphid, Schlechtendalia chinensisobserved using transmission electron microscopeA: 蠟腺Wax gland; B: 角倍蚜蠟腺細胞內微絨毛(mv)中有大量的光面內質網(SER)Apical microvilli (mv) of wax gland cell with lots of smooth endoplasmic reticulum (SER); C: 蠟腺細胞間的連接方式——間隔連接Intercellular junction of wax gland by separate junction. pd: 多邊形凹陷Polygonal depression; ss: 表皮下空間Subcuticular space; mv: 微絨毛Microvilli; N: 細胞核Nucleus; v: 分泌物囊泡Secretion vesicles; mi: 線粒體Mitochondria; sj: 間隔連接Separate junction.
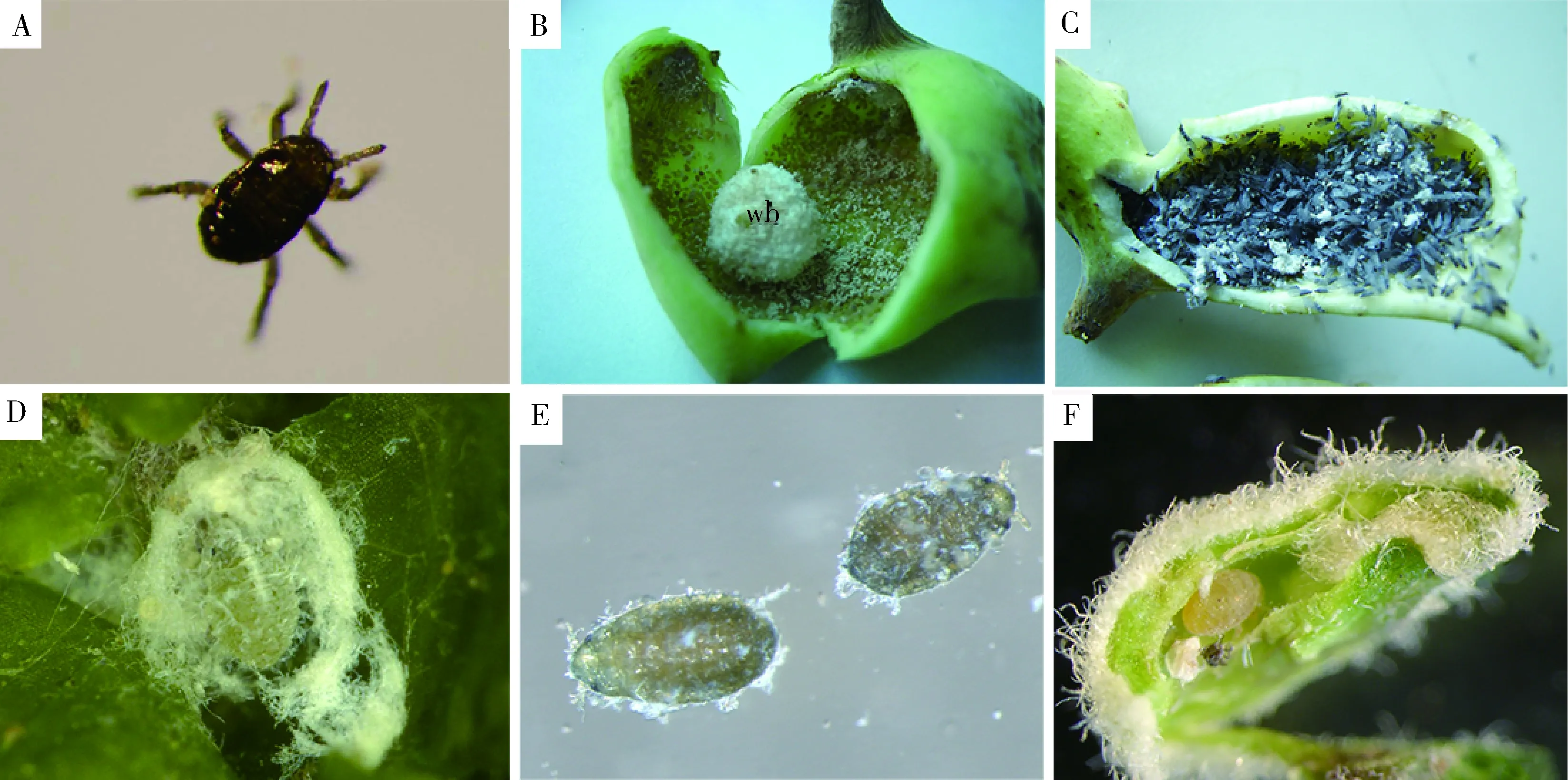
圖4 不同生態環境中的角倍蚜各蟲態的照片和蠟粉分布Fig. 4 Photographs and wax distribution of various body forms of Schlechtendalia chinensisliving in different ecological environmentsA: 1齡干母1st instar fundatrix; B: 干雌,癭內的干雌和蠟粉包裹蜜露形成的蠟球(wb)Fundatrigeniae and a large wax ball (wb) formed by wax wrapping honeydew in a gall; C: 秋遷蚜,蟲癭內的有翅秋遷蚜和蠟粉Fundatrispuriae with wax in a gall; D: 越冬若蚜,蘚上被蠟粉包裹的越冬若蚜Apterae with wax in mosses; E: 性蚜,體表有蠟粉Sexual forms with wax on the body surface; F: 2齡干母,蟲癭內蛻皮后的干母、蠟粉和蛻2nd instar fundatrix after molting, wax and exuviae in a gall.
2.4 角倍蚜各蟲態蠟粉的數量和分布
除1齡干母外(圖4: A),角倍蚜各蟲態的體表均有蠟粉,其中干雌、春遷蚜和秋遷蚜分泌的蠟粉最多,癭內干雌分泌大量蠟粉,包裹蟲體,脫落的蠟粉包裹蜜露、蛻等,有時甚至在癭內形成大的蠟粉球(圖4: B),遷飛前的秋遷蚜,蟲體間有大量的蠟粉(圖4: C);越冬若蚜分泌蠟粉次之,蠟粉包裹自身,在蘚枝上形成松散的蠟球(圖4: D);性蚜的蠟粉比越冬若蚜少(圖4: E),沒有包裹蟲體;干母比較特殊,1齡干母(癭外)蟲體黑色、細長,體表光滑,沒有蠟粉(圖4: A),2齡干母(癭內)蟲體淡黃色、短粗,體表有少量蠟粉(圖4: F)。
3 討論
3.1 角倍蚜各蟲態蠟腺的結構變化及其對生活微環境的適應
蚜蟲分泌的蠟粉具有良好的防水功能,對于在潮濕或寒冷環境中生活的蚜蟲來說更加重要。綠色云杉蚜Elatobiumabietinum可以在冬季經歷幾次霜凍,是因為其表面豐富的蠟粉可以防止水分在蟲體表面凝固從而起到保護作用(Powell and Parry, 1976)。角倍蚜蠟腺結構、形態和凹陷數量等在不同蟲態及同一蟲態的不同部位間差異顯著,其復雜程度與生活微環境密切相關,是角倍蚜對環境條件長期適應的結果。角倍蚜干雌生活在完全封閉的蟲癭內,相對濕度接近100%(劉平, 2014),發達的蠟腺分泌大量的蠟粉,可以有效防止水汽在體表凝結和蟲體粘連。春遷蚜需要從貼近地面的潮濕蘚層中遷飛到鹽膚木樹干上,秋遷蚜需要從蟲癭裂口處遷飛到潮濕的蘚層中(邱明生和趙志模, 1995; 賴永祺等, 1986),發達而均勻的蠟腺分泌蠟粉(圖4: C),可以幫助保持體表干燥,有利于成功遷飛。越冬若蚜生活在貼近地面的高濕度的蘚層中,發達的蠟腺分泌大量蠟粉包裹蟲體(圖4: D),可以有效隔離雨水和防止水汽在體表凝結,度過寒冷的冬季。有研究表明,與形成蠟球的角倍蚜個體相比較,未形成蠟球的個體死亡率較高(賴永祺等, 1990)。
性蚜由遷飛到鹽膚木樹干的春遷蚜在3月中上旬繁殖產生,在濕度較高的樹皮縫內爬行、交配并隱蔽生活(唐覺和張傳溪, 1987),蟲體背部邊緣線上蠟腺的復雜程度要遠遠高于脊柱板和胸膜板上的蠟腺,因此蟲體的邊緣較背面分布有更多的蠟粉(圖4: E),這可能是因為性蚜在潮濕的樹干表面爬行,邊緣線上發達的蠟腺分泌較多的蠟粉可以隔離樹干表面的水。交配后的雌性蚜在鹽膚木樹干上繁殖產干母,1齡干母必須在1~2 d內沿樹干爬到新萌發的嫩葉上取食,刺激葉片組織增生并形成蟲癭包裹自身,在癭內繼續取食并蛻皮,完成從癭外到癭內的轉變;1齡干母需要快速爬行到梢頭尋找嫩葉取食(唐翊峰, 2014),蠟粉會影響其快速爬行,因此其蠟腺缺失或退化;當干母被包裹進封閉的蟲癭內后,為適應癭內高濕度的微環境,干母蛻皮后出現蠟腺,分泌蠟粉防止水汽在體表凝結,保證了其正常的生存和繁殖。因此,干母在個體發育中蠟腺的從無到有,正是干母對從癭外到癭內的微環境變化的適應。
可以看出,經過長期的進化,角倍蚜各蟲態均具有與其生活微環境相適應的蠟腺結構,保證了種群的繁衍,但仍存在一些薄弱環節。如1齡干母沒有蠟腺,不能分泌蠟粉,如遇到初春的連續陰雨,將會由于缺乏蠟粉的保護而大量死亡;此外,秋季在蘚上初產的越冬若蚜,如遭遇陰雨,也會由于蠟粉分泌不足和蠟球未完成形成而死亡。因此,在角倍蚜培育過程中,可以有針對性地研發或改進培育技術,如通過積溫調控干母發生時間,在蘚圃上覆蓋薄膜等,避開早春和晚秋的陰雨,可以有效降低初產干母和越冬若蚜的死亡率,提高五倍子產量。
3.2 角倍蚜蠟腺細胞的超微結構與功能
昆蟲的外分泌腺細胞根據表皮結構和分泌物的分泌方式可以分為class 1和class 3,屬于class 1的腺體細胞被表皮細胞覆蓋,分泌物需要穿過表皮;而屬于class 3的腺體細胞與導管相連,分泌物通過導管到達細胞外(Noirot and Quennedey, 1974)。角倍蚜蠟腺細胞被表皮細胞覆蓋,蠟的分泌必須穿過表皮這個屏障,表明角倍蚜蠟腺分泌細胞屬于class 1(圖3: A);蠟腺細胞中和頂端微絨毛中存在大量的光面內質網,其作用是將脂類分泌物帶到微絨毛,然后通過微絨毛分泌,微絨毛增加了蠟腺細胞的面積,提高了分泌效率(Wangetal., 2018)。蠟腺細胞中存在大量的線粒體,可能是為蠟的分泌提供能量(Billenetal., 2017)。
3.3 角倍蚜蠟粉在癭內封閉微環境中的保護功能
蚜蟲通過刺吸式口器取食植物汁液,食物中多余的大量含糖液體以蜜露的形式被排出。自由生長的蚜蟲通常排放大量的蜜露(Stadler and Müller, 1996),一些生活在半封閉蟲癭內的蚜蟲可以將蜜露、蠟粉和蛻等從開口處排出(Smith, 1999)。但角倍蚜大量的干雌(通常每個蟲癭內5 000~8 000頭)生活在完全封閉的蟲癭內長達150多天(Shaoetal., 2013; 陸沁等, 2018),其排泄的大量蜜露是如何處理的呢?最新的研究表明角倍蚜蟲癭內壁可以將蜜露吸收和再利用,成為蟲癭生長的營養來源之一(Chenetal., 2020)。除此之外,角倍蚜癭內干雌發達的蠟腺分泌大量的蠟粉,可以將多余的蜜露和蛻等包裹,形成較大的蠟球(圖4: B),有效地防止蜜露的污染和蟲體間的粘連。蟲癭的生長期為每年5-10月,跨越最熱的夏季,癭內的高濕、高溫、封閉的環境和高糖的蜜露,具備了病原菌發生的良好條件,癭內大量的蠟粉,還可以有效防止病原菌孢子萌發和菌絲生長(Pikeetal., 2002),保證了癭內角倍蚜種群的正常生長和繁殖。前人的研究也證明山毛櫸蚜Phyllaphisfagi分泌的蠟粉可以防止黑色霉菌的發生(Smith, 1999)。
綜上所述,角倍蚜蠟粉的主要功能是防水和包裹蜜露等,各蟲態蠟腺的數量、排列和發達程度與其生活微環境和各蟲態的生物學特性密切相關,這些特征可能是角倍蚜對環境條件長期適應的結果。隨著角倍蚜全基因組的發布,聯合基因組、轉錄組和蛋白質組學數據鑒定角倍蚜的泌蠟基因,分析泌蠟基因在各個蟲態間的表達差異,將有助于從分子層面揭示角倍蚜蠟腺在環境適應和進化中的生態學意義。
