致癭昆蟲對寄主植物生理和代謝的影響
2021-05-25 04:07:46楊孟可喬海莉徐常青
昆蟲學報 2021年4期
關鍵詞:植物
楊孟可, 劉 賽, 喬海莉, 郭 昆, 徐 榮, 徐常青, 陳 君
(中國醫學科學院北京協和醫學院藥用植物研究所, 北京 100193)
癭(gall)在植物病理學上是指植物受刺激導致非正常發育的細胞、組織或器官,在病毒、細菌、真菌、線蟲、螨類及昆蟲等生物影響下,植物細胞非正常增殖和分化而形成,是自然界極其常見的生物學現象(Mani, 1964)。由于昆蟲是最主要的致癭類群(gall-inducers/galler),因此,狹義上把植物上的癭普遍稱為蟲癭,這類昆蟲稱為致癭昆蟲(gall-inducing insects)。據統計,全世界約有130 000種昆蟲可誘導植物形成蟲癭(Espírito-Santoetal., 2007),主要集中在纓翅目、半翅目、鞘翅目、鱗翅目、雙翅目和膜翅目(Ramanetal., 2009);植物幾乎每個部位都可形成蟲癭,如根、莖、葉、花、果實等,其中75%以上的蟲癭產生于葉緣、葉片、葉脈和葉柄(Inbaretal., 2004) (表1)。大部分致癭昆蟲的寄主植物種類單一,一種昆蟲只能在某種特定植物、特定器官或親緣關系相近的植物上形成蟲癭(Abrahamsonetal., 1998)。綿蚜亞科(Eriosomatinae)主要在榆屬Ulmus植物上形成蟲癭,癭綿蚜亞科(Pemphiginae)主要在楊屬Populus植物上形成蟲癭,五節根蚜亞科(Fordinae)主要在黃連木屬Pistacia和鹽膚木屬Rhus植物上形成蟲癭(申潔等, 2016)。

表1 主要致癭昆蟲Table 1 Main gall-inducing insects
致癭昆蟲幾乎整個生活史都在蟲癭內完成,與寄主植物關系十分密切。致癭昆蟲對植物的影響主要體現在兩個方面:(1)影響寄主植物細胞和組織結構,如引起植物細胞空泡化、染色質和細胞器增加、細胞壁變薄、胞間連絲變大和氣孔導度降低等(Florentineetal., 2005; Gironetal., 2016; 林晨等, 2016),造成植物葉片卷曲、斑狀突起或畸形膨大, 甚至形成更為復雜的蟲癭結構以保護致癭昆蟲(Hartley, 1998; Stone and Sch?nrogge, 2003)。(2)影響寄主植物的生理和代謝,如影響葉片光合作用 (Patankaretal., 2011),匯聚更多營養物質及次生物質(Martinsonetal., 2015),而有利于蟲癭內致癭昆蟲的生長發育(Nabityetal., 2013)。蟲癭是研究昆蟲與植物協同進化的最佳模式材料,明確致癭昆蟲對寄主植物的影響不僅有助于進一步認識致癭昆蟲與植物的關系,為致癭昆蟲與植物協同進化提供理論支撐,而且有助于了解植物對生物脅迫的響應過程,從而為生物因子與植物互作研究提供指導和借鑒。
1 致癭昆蟲對寄主植物光合作用的影響
綠色植物通過光合作用為各種生理活動提供能量來源,也為植物抵御不良環境和生物脅迫提供物質基礎,致癭昆蟲可影響植物葉綠素含量(Carneiroetal., 2014),調節植物光合速率(Patankaretal., 2011),甚至影響寄主的代謝產物分布和防御響應(Bagattoetal., 1996; Larson, 1998)。致癭昆蟲對寄主植物光合作用的影響,主要表現在光合色素、光合速率和葉綠素熒光參數3個方面(表2)。

表2 致癭昆蟲對寄主植物光合作用的影響Table 2 Influences of gall-inducing insects on the photosynthesis of host plants
光合色素是植物光合作用中參與光能吸收、傳遞以及引起原初光化學反應的色素,包括葉綠素和類胡蘿卜素兩大類,均排列在葉綠體的類囊體膜上(潘瑞熾等, 2004)。葉綠素是高等植物參與光合作用的重要色素,與光合能力以及發育階段有較好的相關性,類胡蘿卜素則是植物光合作用的重要輔助色素(張金恒等, 2003)。致癭昆蟲能夠影響植物葉綠素含量及其生物合成,幾乎所有蟲癭都存在葉綠素含量減少的現象,如木虱科昆蟲Nothotriozamyrtoidis在桃金娘科植物Psidiummyrtoides上誘導形成的蟲癭,其葉綠素含量顯著低于健康葉片(Carneiroetal., 2014)。而且,一些蟲癭不同部位的葉綠素降解代謝也有所差異,如紅楠Machilusthunbergii非蟲癭部位葉綠素降解途徑為:葉綠素(chlorophyll)→葉黃素(pheophytin)→脫鎂葉綠酸(pheophorbide),而蟲癭部位為:葉綠素→脫植基葉綠素(chlorophyllide)→脫鎂葉綠酸(Yangetal., 2003)。蟲癭部位類胡蘿卜素也有減少的現象,如木虱科昆蟲Pseudophacopteronsp.在白堅木屬植物Aspidospermaaustrale上形成的蟲癭中類胡蘿卜素含量顯著低于健康葉片(Oliveiraetal., 2011)。
絕大多數致癭昆蟲會導致植物凈光合速率降低,如亞洲栗癭蜂Dryocosmuskuriphilus使蟲癭葉片CO2同化率降低30%(Ugolinietal., 2014)。櫻桃植羽癭螨Phytoptuscerasicrumena和北美五倍子蚜Melaphisrhois在黑野櫻Prunusserotina和光葉漆Rhusglabra上形成蟲癭后,兩種蟲癭葉片光合速率分別比健康株葉片降低了47%和50%;櫻桃植羽癭螨致癭株系統葉片光合速率比健康株葉片降低了24%,五倍子蚜致癭株系統葉片光合速率與健康株相比無顯著變化(Larson, 1998)。然而,也有些致癭昆蟲對寄主植物的光合速率并無顯著影響,如橄欖星室木虱Pseudophacopteronaspidospermi在白堅木屬植物A.australe上誘導的蟲癭葉片和健康葉片相比,光合速率無顯著差異(Malenovskyetal., 2015),甚至某些致癭昆蟲還會為植物提供“好處”,即增強植物光合作用,如癭蜂Antistrophussilphii在松香草屬植物Silphiumspeciosum上形成蟲癭后可顯著提高木質部水勢,保證葉片在干旱條件下的含水量,進而提高光合速率(Fayetal., 1993)。金小蜂Trichilogastersigniventris在密花相思Acaciapycnantha的葉狀柄上形成蟲癭后通過提高寄主光合速率以滿足自身營養需求,改變與相鄰組織間的“源-庫”關系(Dorchinetal., 2006)。
葉綠素熒光參數具有反映植物光合作用“內在性”的特點,借助葉綠素熒光技術發現蟲癭部位的實際光合效率YII、最大光合效率Fv/Fm等光合參數也受到了影響(Oliveiraetal., 2011),如致癭昆蟲氈蚧Bystracoccusmataybae誘導的蟲癭部位實際光合效率低于健康葉片(Oliveiraetal., 2017),紅楠M.thunbergii上兩種癭蚊Daphnephilasueyenae和D.taiwanensis誘導的蟲癭部位最大光合效率均低于健康部位(Huangetal., 2011),一種癭蚊Bruggmanniellasp.在長葉木姜子Litseaacuminata上誘導的蟲癭部位最大光合效率較同一葉片的非蟲癭部位和健康葉片顯著降低(Huangetal., 2015)。
2 致癭昆蟲對寄主植物初生代謝的影響
致癭昆蟲生活史中大部分時間在蟲癭內部,因此致癭昆蟲對寄主植物營養物質產生特異性依賴(Florentineetal., 2005)。蟲癭形成的“營養假說”(nutrition hypothesis)也認為蟲癭的形成是為了給致癭昆蟲提供營養物質(Stone and Sch?nrogge, 2003)。蟲癭富含可溶性糖、蛋白質、脂類等營養物質(Raman, 2010)(表3)。
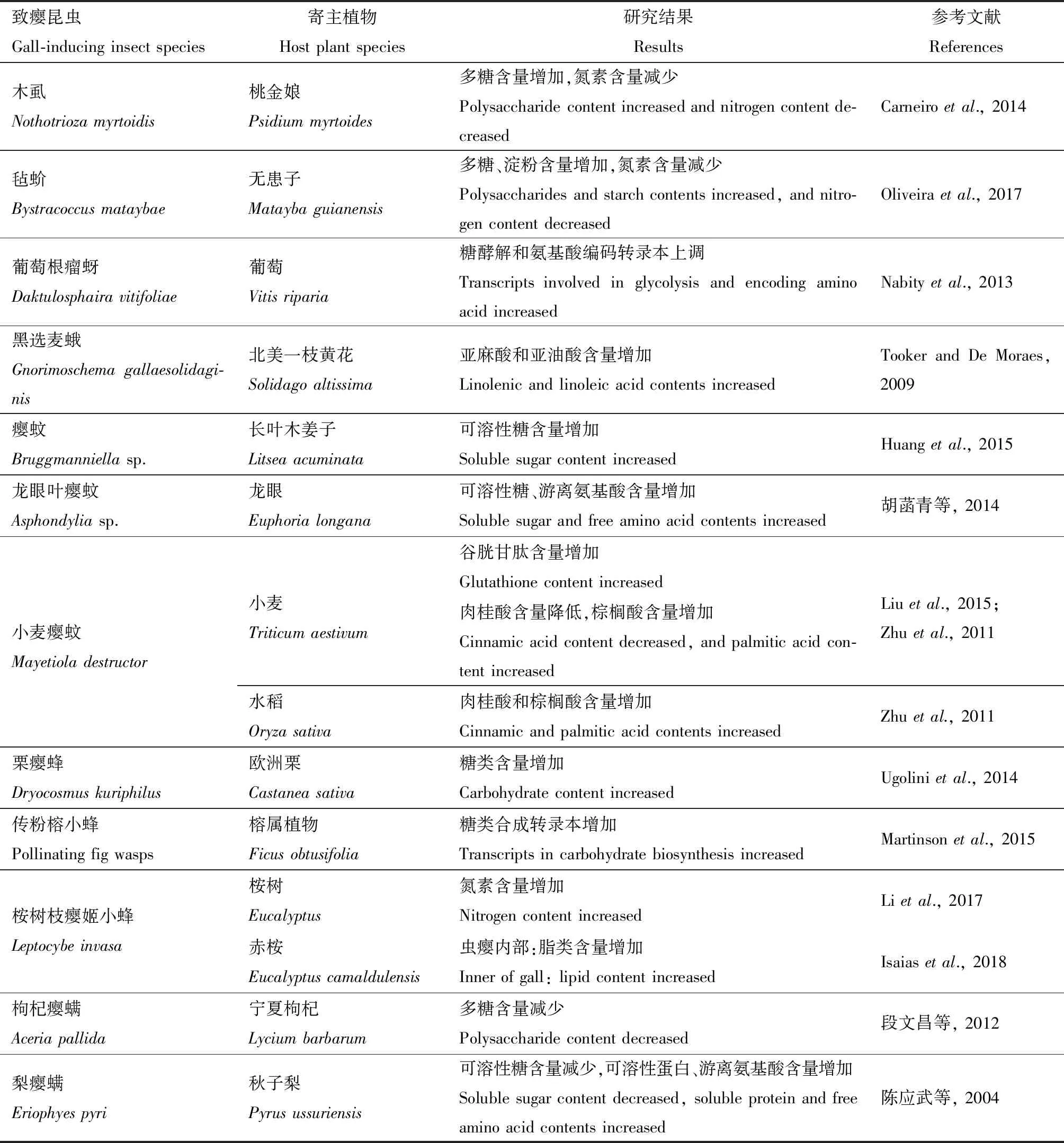
表3 致癭昆蟲對寄主植物初生代謝的影響Table 3 Influences of gall-inducing insects on the primary metabolism of host plants
2.1 對糖類物質的影響
糖類是致癭昆蟲的主要能源物質,致癭昆蟲普遍引起被害葉片可溶性糖、淀粉等糖類物質增加,如長葉木姜子L.acuminata上的一種癭蚊Bruggmanniellasp.誘導形成的蟲癭葉片可溶性糖含量高于健康葉片(Huangetal., 2015);龍眼葉癭蚊Asphondyliasp.在龍眼Dimocarpuslongana葉片致癭后可溶性總糖和蔗糖含量顯著高于正常葉片(胡菡青等, 2014)。蟲癭組織和非蟲癭組織的糖類含量也有差異,如小麥癭蚊Mayetioladestructor在小麥葉片致癭后還原糖、總可溶性糖、淀粉等營養物質表現為從蟲癭周邊組織向蟲癭部位轉移(Ranietal., 2017)。同時,致癭昆蟲還會影響寄主植物體內糖類代謝路徑基因表達,如葡萄根瘤蚜Daktulosphairavitifoliae為害葡萄Vitisriparia葉片后可引起寄主體內與糖類轉運和降解相關基因普遍上調表達(Nabityetal., 2013),這些現象為蟲癭形成的“營養假說”提供了有力證據。
2.2 對蛋白質和氨基酸的影響
蛋白質和氨基酸是一切生命體及生命活動的物質基礎,蟲癭為致癭昆蟲提供了豐富的蛋白質和氨基酸營養, 如梨癭螨Eriophyespyri致癭后可引起秋子梨Pyrusussuriensis葉片中可溶性蛋白質含量上升34.14%,游離氨基酸含量上升29.95%,游離脯氨酸含量上升84.08%(陳應武等, 2004)。葡萄根瘤蚜D.vitifoliae為害葡萄V.riparia葉片后可引起編碼氨基酸和寡肽合成的基因上調表達(Nabityetal., 2013)。此外,蟲癭的某些蛋白還有特異性分布的現象,2種癭蜂Andricusquercuscalicis和Diplolepisspinosa分別在橡樹Quercusrobur和犬薔薇Rosacanina上誘導形成的蟲癭中檢測出了兩個含量非常豐富的蛋白序列,相對分子質量分別為62和43 kD(Sch?nroggeetal., 2000)。植物體內某些抗性相關蛋白含量的增加還與其發揮防御功能有關,如小麥癭蚊M.destructor持續侵染小麥72 h后,葉片中谷胱甘肽含量顯著增加,谷胱甘肽能夠在酸性條件、滲透壓異常情況下保護細胞免受損傷,同時還參與植物防御生物和非生物脅迫時的信號轉導(Ghanta and Chattopadhyay, 2011; Liuetal., 2015)。
2.3 對脂類物質的影響
脂類是致癭昆蟲的重要儲能物質,但絕大多數致癭昆蟲自身無法合成脂肪酸,致癭后通常可引起蟲癭部位脂類物質含量增加,如致癭昆蟲黑選麥蛾Gnorimoschemagallaesolidaginis在北美一枝黃花Solidagoaltissima上誘導的蟲癭中,亞麻酸和亞油酸含量顯著高于健康葉片,這兩種游離脂肪酸均是鱗翅目昆蟲幼蟲發育和成蟲羽化的必需脂肪酸(Tooker and De Moraes, 2009)。此外,脂類還可作為植物的抗性物質發揮防御作用,如小麥癭蚊M.destructor為害可快速引起水稻葉片中軟脂酸、硬脂酸和油酸3種脂肪酸含量增加,其中軟脂酸和硬脂酸是植物表面蠟質合成的前體物質(Zhuetal., 2011)。黃連木角癭綿蚜Baizongiapistaciae在黃連木屬植物Pistaciapalaestina上誘導的蟲癭中富含大量的油性樹脂,該成分還能夠保護葉片免受動物取食和微生物侵染(Randetal., 2014)。
3 致癭昆蟲對寄主植物次生代謝的影響
次生代謝物質是植物適應各種環境脅迫(生物的和非生物的)的重要手段,致癭昆蟲作為一種特殊的生物脅迫因子,與植物有著特殊的關系。一方面植物產生大量的次生物質以防御致癭昆蟲為害,另一方面致癭昆蟲利用這些次生物質以保護蟲癭不被其他植食者取食。目前對于蟲癭中次生代謝產物的研究以非揮發性物質酚類和揮發性物質萜類兩類成分為主(表4),主要從兩個層面進行了研究:(1)宏觀層面將葉片分為蟲癭組織和非蟲癭組織,普遍認為蟲癭組織比非蟲癭組織含有更多的次生代謝物質(Halletal., 2016);(2)微觀層面將蟲癭結構分為內部營養組織和外部防御組織,營養組織富含營養物質,防御組織富含生物堿、黃酮類、酚類等次生物質,保障致癭昆蟲免受其他生物的寄生或捕食(Bragan?aetal., 2016)。
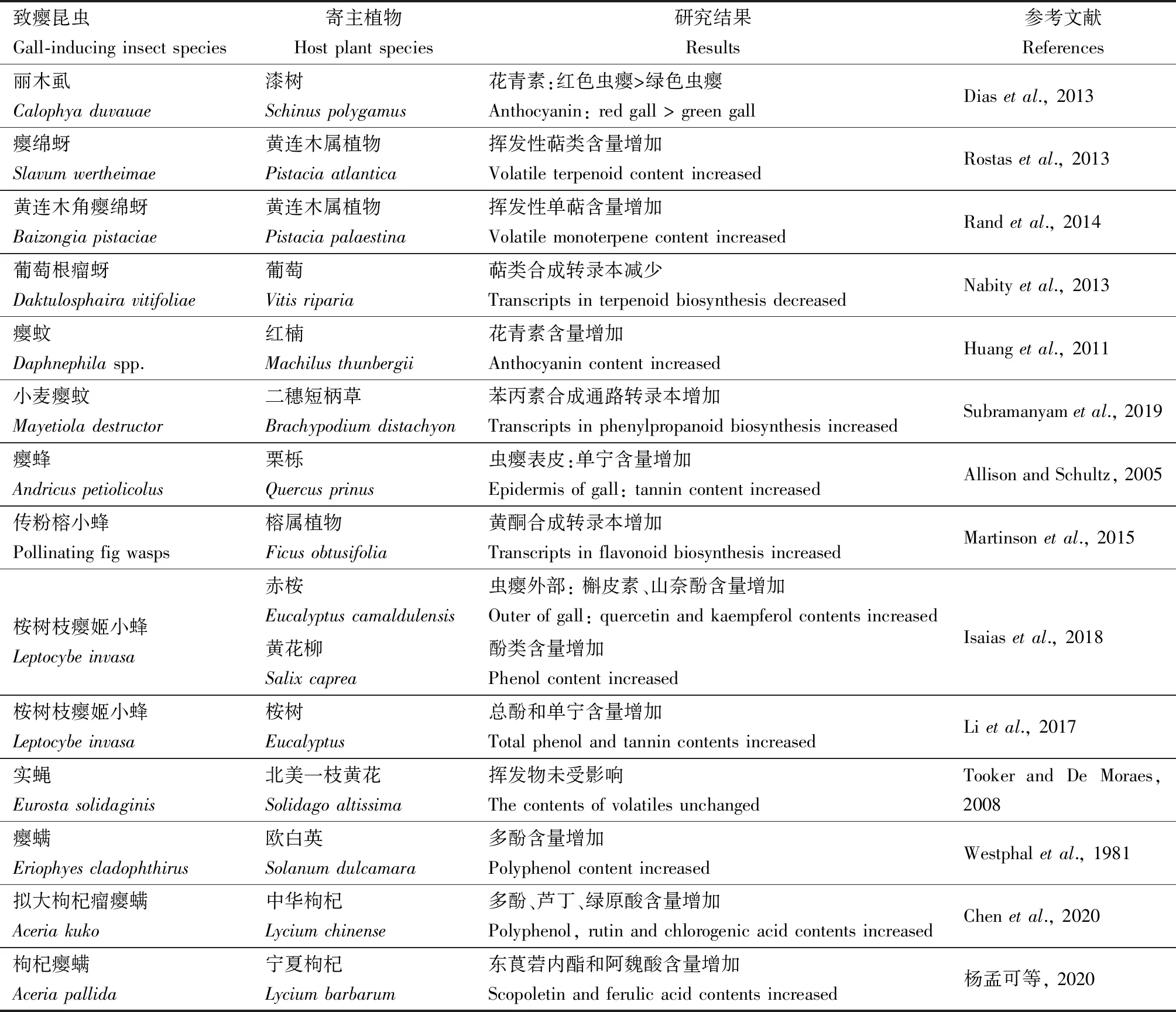
表4 致癭昆蟲對寄主植物次生代謝的影響Table 4 Influences of gall-inducing insects on the secondary metabolism of host plants
3.1 對非揮發性次生代謝物質的影響
非揮發性次生代謝物質通常被視為植物的直接防御手段,許多植物遭受植食性昆蟲為害后會積累大量次生代謝物質。致癭昆蟲常誘導寄主植物合成大量酚類物質,且隨蟲癭的生長而變化,如兩種葉蜂Pontaniaproxima和Pontaniapedunculi分別誘導白柳Salixalba和黃花柳Salixcaprea葉片致癭并合成酚類物質,蟲癭中酚類含量隨蟲癭成熟而降低,且始終高于正常葉片,但在致癭初期除去幼蟲后,蟲癭中酚類含量迅速降至正常水平(Hartley, 1998)。多項研究表明酚類在致癭昆蟲與植物互作中發揮了重要作用,主要體現在:(1)植物對致癭昆蟲的抗性與酚類化合物含量呈正相關,致癭昆蟲的存活率與蟲癭中酚類含量呈負相關(Westphaletal., 1981)。(2)酚類化合物參與寄主植物的基因表達和生長發育的調控,酚類已被證實在根瘤菌誘導寄主根部形成蟲癭的過程中充當入侵植物組織的信號物質(Ranade and David, 1985; Dakoraetal., 1993)。(3)酚類化合物可充當植物體內吲哚-3-乙酸氧化酶抑制劑,高濃度酚類抑制吲哚-3-乙酸(IAA)降解從而調控致癭部位的細胞增大和分化(Carneiroetal., 2014)。單寧(又稱鞣酸、鞣質)是一類重要的酚類物質,蟲癭組織單寧含量通常高于非蟲癭組織,甚至達某些蟲癭干重的60%~70%(Howes, 1953)。中藥五倍子來源于癭綿蚜科五倍子蚜Melaphischinensis寄生在鹽膚木Rhuschinensis及其同屬其他植物形成的蟲癭,其單寧含量平均在40%左右,最高可達70%以上(李秀萍等, 2002)。
致癭昆蟲通常還會引起蟲癭部位顏色的變化,其中起主導作用的是酚類中的花青素,它是植物體內一種重要的保護性色素,能夠保護植物組織免受致癭昆蟲引起的氧化應激損傷(Vanderauweraetal., 2009)。致癭昆蟲麗木虱Calophyaduvauae誘導的紅色蟲癭中花青素含量顯著高于綠色蟲癭及正常葉片,Dias等(2013)推斷這是由于紅色蟲癭內有致癭昆蟲的持續刺激,而綠色蟲癭內致癭昆蟲被天敵寄生而無法持續刺激。由于致癭昆蟲移動能力有限,只能通過對寄主植物的調控降低生存風險,一方面操控寄主組織結構(蟲癭)進行物理防御,另一方面操控寄主次生代謝進行化學防御。蟲癭內部組織作為致癭昆蟲賴以生存的營養供給站,而外部組織則是致癭昆蟲抵抗風險的保護傘。因此,蟲癭內部組織通常富含較多的營養物質,蟲癭外部組織則含有較多的次生物質,如桉樹枝癭姬小蜂L.invasa在赤桉Eucalyptuscamaldulensis葉片上誘導的蟲癭中,外部組織比內部組織含有更高濃度的槲皮素和山奈酚衍生物,黃酮類化合物有助于植物抵抗紫外輻射和調節細胞氧化還原穩態,從而保護蟲癭(Isaiasetal., 2018)。
3.2 對揮發性次生代謝物質的影響
揮發性有機化合物(volatile organic compounds, VOCs)是植物的一種間接防御手段,許多植物受植食性昆蟲取食后會釋放萜類等揮發性有機物,這些揮發性有機物可作為信號物質吸引天敵或驅避競爭者(Turlings and Ton, 2006)。高等植物中萜類物質的生物合成需消耗大量能量,蟲癭內積累的大量萜類物質可能是由植物葉片或其他組織合成后轉運而來(Gershenzon, 1994; Larson and Whitham, 1997)。致癭昆蟲能夠誘導植物產生VOCs,如黃連木角癭綿蚜B.pistaciae誘導黃連木屬植物P.palaestina形成的蟲癭中萜類含量超出健康葉片10~60倍,且蟲癭葉片和非蟲癭葉片的萜類組成有所不同,蟲癭中的萜類是以α-蒎烯和檸檬烯為主的單萜類型,葉片中的萜類是以E-石竹烯、吉瑪烯D和δ-卡丁烯為主的倍半萜類型(Randetal., 2014)。但也有研究發現致癭昆蟲對寄主植物產生的VOCs無顯著影響或抑制寄主植物產生VOCs,如致癭昆蟲實蠅Eurostasolidaginis和黑選麥蛾G.gallaesolidaginis幼蟲均未引起寄主北美一枝黃花S.altissima揮發物含量發生顯著變化,而且還可抑制其他昆蟲煙芽夜蛾Heliothisvirescens誘導產生VOCs(Tookeretal., 2008)。此外,蟲癭揮發物還可抑制其他植食者取食,如癭綿蚜Slavumwertheimae誘導黃連木屬植物Pistaciaatlantica形成的蟲癭后,其揮發物能對山羊產生明顯驅避作用(Rostasetal., 2013)。
4 致癭昆蟲對寄主植物保護酶的影響
抗氧化酶系統是植物遭受環境脅迫時的重要防御系統,致癭昆蟲作為一種生物脅迫因子,會破壞寄主細胞內的穩態環境,引起細胞內活性氧含量增加,使植物細胞中一系列保護酶活性發生變化,包括超氧化物歧化酶(superoxide dismutase, SOD)、過氧化氫酶(catalase, CAT)、過氧化物酶(peroxidase, POD)、多酚氧化酶(polyphenol oxidase, PPO)等組成的蛋白酶系統,以減輕對植物的損傷(楊舒貽等, 2016)(表5)。刺桐姬小蜂Quadrastichuserythrinae為害引起刺桐Erythrinavariegate葉片尤其是蟲癭組織中SOD, CAT和PPO活性顯著提高(汪少妃等, 2014)。癭蜂Andricuspetiolicolus在栗櫟Quercusprinus上誘導的蟲癭中皮層和表皮的POD含量高于內部營養組織,由于POD可激活和聚合植物體內的多酚,顯示蟲癭不僅可以為致癭昆蟲提供生長發育所需營養,而且蟲癭外部高活性的POD可抵御其他昆蟲和病原菌,以保護蟲癭內部組織(Allison and Schultz, 2005)。
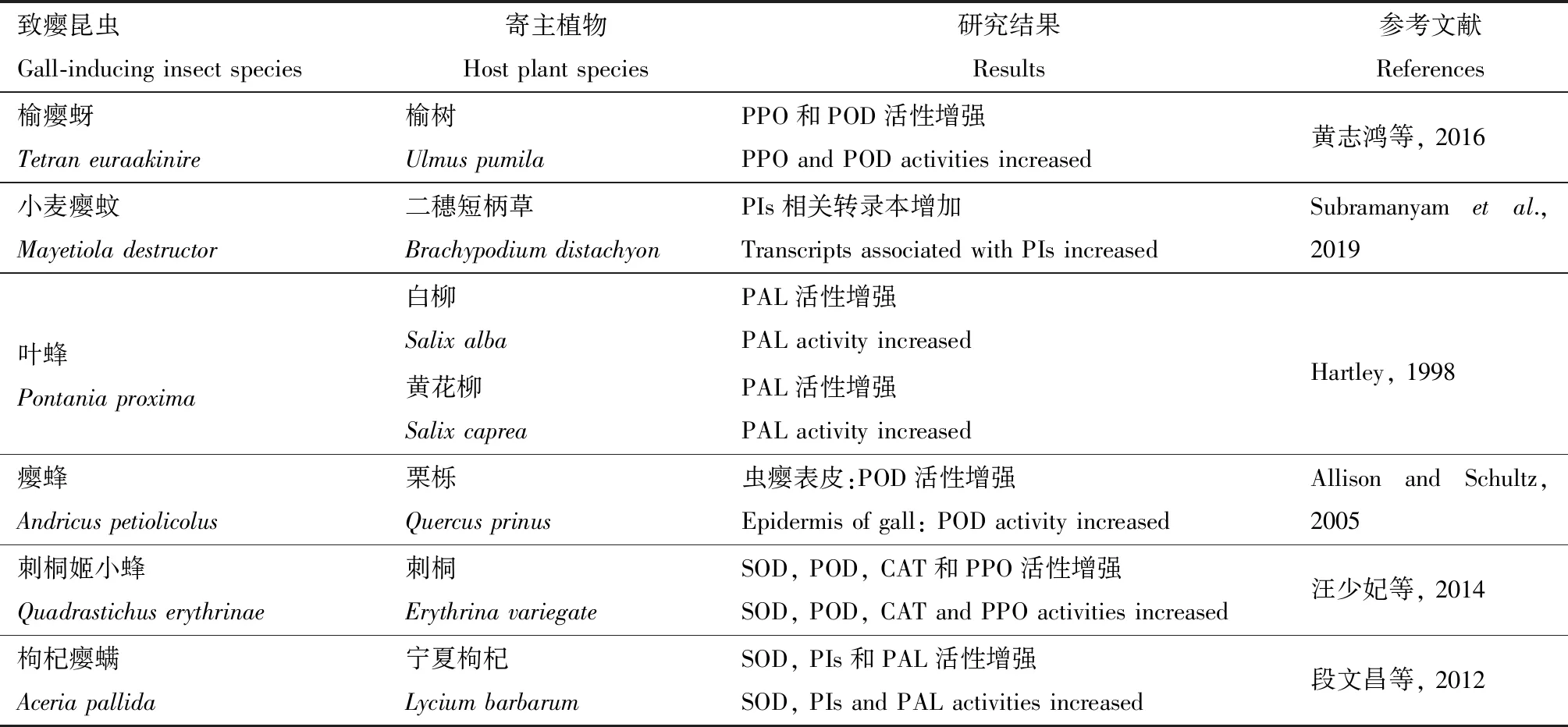
表5 致癭昆蟲對寄主植物保護酶的影響Table 5 Influences of gall-inducing insects on the defensive enzymes in host plants
蛋白酶抑制劑(protease inhibitors, PIs)和苯丙氨酸解氨酶(phenylalanine ammonia lyase, PAL)也在植物抗逆過程發揮了重要作用。PIs可與昆蟲腸道內蛋白水解酶結合,抑制腸道對蛋白質的消化,導致昆蟲營養不良和生長發育受阻(曹鑫等, 2011); PAL是植物體內連接初生代謝和次生代謝的橋梁物質,在細胞分化、木質化、黃酮類、生物堿類等次生代謝物質合成方面發揮重要作用(黃小貞和趙德剛, 2017)。已有研究顯示致癭昆蟲普遍引起植物PIs和PAL活性升高,如兩種葉蜂P.proxima和P.pedunculi侵染早期,蟲癭組織中PAL活性高于非蟲癭組織(Hartley, 1998),枸杞癭螨Aceriapallida為害可引起寧夏枸杞Lyciumbarbarum葉片中的PIs和PAL活性提高(段文昌等, 2012),在小麥癭蚊M.destructor為害1, 3和5 d后,二穗短柄草Brachypodiumdistachyon葉片中18個PIs合成相關的差異表達基因(differentially expressed genes, DEGs)逐漸上調表達(Subramanyametal., 2019)。蟲癭部位PIs和PAL酶活增強預示該部位次生代謝物質合成活躍,一方面用于防御致癭昆蟲,同時還可抵御其他病蟲為害。
5 致癭昆蟲對寄主植物內源激素的影響
植物內源激素不僅是植物生長發育的重要調節物質,還與蟲癭形成及發育密切相關(表6),目前生長素(indol-3-acetic acid, IAA)和細胞分裂素(cytokinin, CTK)被廣泛認為在蟲癭形成過程中起主導作用,包括刺激寄主植物細胞生長和分化(Tooker and Helms, 2014)。如致癭昆蟲黑選麥蛾G.gallaesolidaginis為害引起寄主北美一枝黃花S.altissima莖中IAA含量顯著增加,且隨著蟲癭成熟而逐漸減少(Tooker and De Moraes, 2011)。除了在植物組織中檢測到IAA,在致癭昆蟲體內也存在IAA,如一種葉蜂Pontaniasp.的幼蟲已被證實可以利用色氨酸合成IAA,致癭后的寄主柳屬植物Salixjaponica轉錄組結果顯示,蟲癭部位的生長素合成相關基因較其他組織顯著上調表達(Yamaguchietal., 2012)。某些蟲癭形態存在性別二態性也與雌雄幼蟲體內IAA含量差異有關,如一種金小蜂Trichilogasteracaciaelongifoliae在長葉相思樹Acacialongifolia上誘導的蟲癭結構與金小蜂的性別相關,雌蜂誘導的蟲室體積、貯藏組織和維管組織分別是雄蜂誘導的3.3, 1.5和3.5倍,這與雌性幼蟲體內IAA含量高活性強而雄性幼蟲體內IAA活性相對較弱有關(Dorchinetal., 2009)。赤霉素(gibberellin, GA)、脫落酸(abscisic acid, ABA)等激素也與蟲癭的生長發育有關,如桉樹枝癭姬小蜂L.invasa在桉樹Eucalyptusexserta葉片上誘導的蟲癭中積累大量的GA(Lietal., 2017),而一種葉蟬Cicadulinabipunctata在玉米上誘導的蟲癭中GA1和GA2含量減少,移除C.bipunctata后GA1和GA2含量與對照組相當,顯示致癭昆蟲是誘導寄主植物內源激素變化的誘因(Tokudaetal., 2013)。樸奶頭癭木虱Pachypsyllaceltidismamma致癭后引起美洲樸Celtisoccidentalis葉片組織中ABA含量增加(Strakaetal., 2010),而小麥癭蚊M.destructor不同侵染階段均引起兩種寄主小麥和水稻體內ABA含量減少(Zhuetal., 2011)。
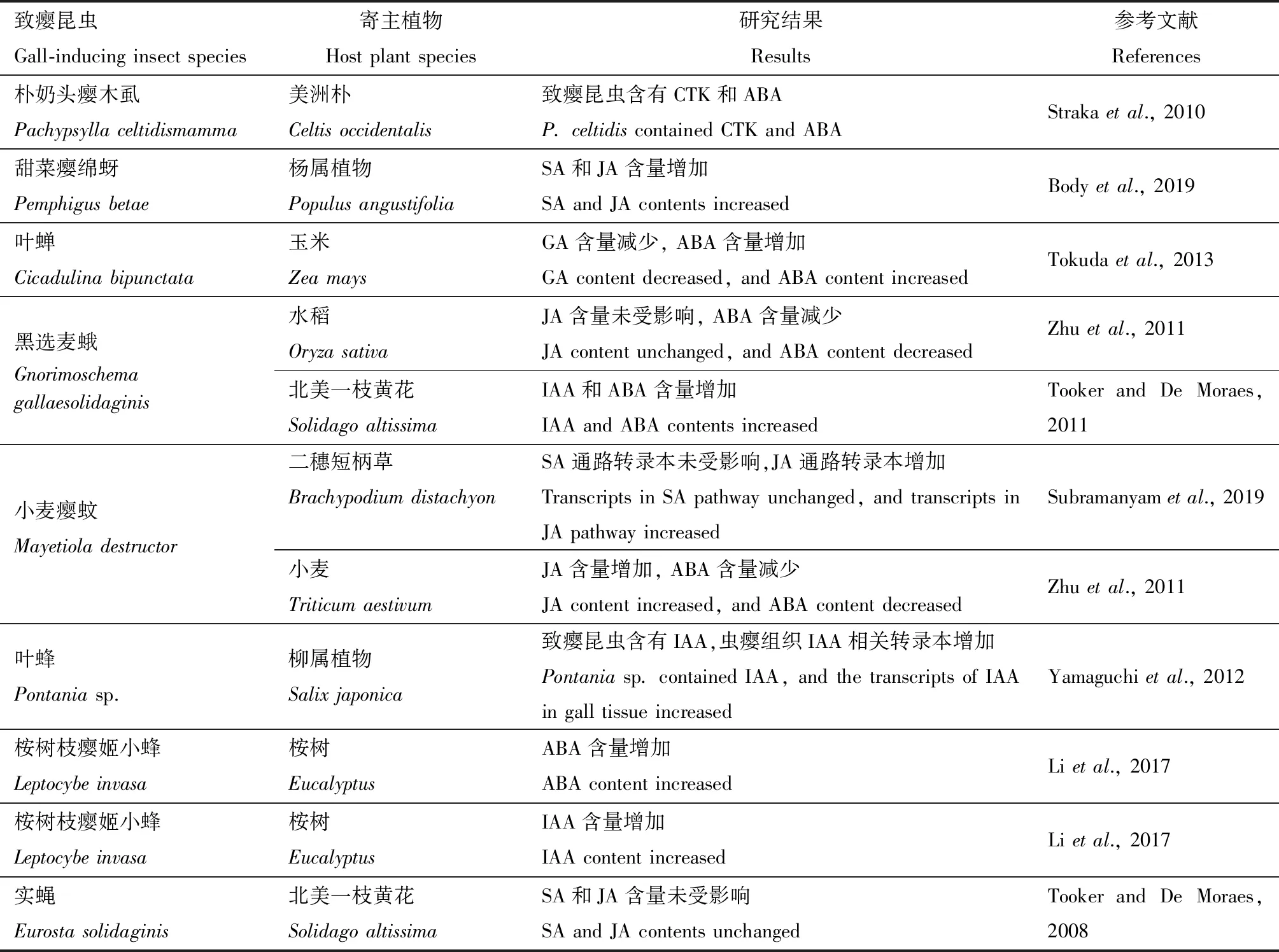
表6 致癭昆蟲對寄主植物內源激素的影響Table 6 Influences of gall-inducing insects on phytohormones in host plants
植物內源激素不僅與蟲癭生長密切相關,還是介導寄主植物防御反應的重要信號分子,主要包括水楊酸(salicylic acid, SA)、茉莉酸(jasmonic acid, JA)和乙烯(ethylene, ET)3種激素。一般情況下,咀嚼式口器昆蟲誘導植物體內SA途徑,刺吸式口器昆蟲誘導植物體內JA途徑,而致癭昆蟲包括咀嚼式口器和刺吸式口器兩類昆蟲,因此這一類昆蟲可誘導JA途徑和SA途徑(Tooker and Helms, 2014)。致癭昆蟲致癭可引起植物SA或JA含量單獨增加,如小麥癭蚊M.destructor分別侵染水稻6, 24和48 h后均導致SA含量顯著增加,但對JA含量無顯著影響(Zhuetal., 2011);榆癭蚜Tetraneuraakinire為害不同時期均引起榆樹Ulmuspumila葉片中JA含量顯著增加,但對SA含量無顯著影響(黃智鴻等, 2016)。也有研究顯示致癭昆蟲可誘導SA和JA含量同時增加,如致癭昆蟲甜菜癭綿蚜Pemphigus
betae為害引起寄主楊屬植物Populusangustifolia葉片中SA和JA含量均顯著增加,且在5種楊樹品種中,品種抗性越強則SA和JA含量越高(Bodyetal., 2019)。然而,也有不少研究顯示致癭昆蟲還可抑制植物抗性反應,對防御激素的影響不明顯,如致癭昆蟲實蠅E.solidaginis和黑選麥蛾G.gallaesolidaginis為害均未引起寄主北美一枝黃花S.altissima體內的JA含量發生明顯變化(Tooker and De Moraes, 2008)。
植物內源激素不僅能夠單獨發揮效應,不同激素組合還能發揮協同或拮抗效應。已有研究顯示,IAA與SA/JA存在負向交互作用,即IAA增加可抑制SA/JA的合成(Erbetal., 2012),在蟲癭形成的過程中,寄主植物體內(尤其是蟲癭部位)IAA含量普遍增加,推測這有利于抑制寄主植物產生SA/JA抗性,從而利于致癭昆蟲。
6 小結與展望
蟲癭是自然界極為常見的生物現象,形態多樣結構復雜,致癭生物類別眾多,其中昆蟲是最主要的致癭生物。致癭昆蟲一方面可改變寄主植物細胞結構和細胞器數目,引起寄主植物組織增生和分化,將自己保護在蟲癭內部,另一方面,致癭昆蟲還可調控寄主植物光合作用實現對初生和次生代謝產物的調配,從而讓蟲癭成為營養洼地(nutrient sink),匯聚周圍組織營養以滿足自身需求,并通過對寄主體內保護酶和防御激素的調控,降低植物對自身的防御響應,同時在蟲癭內外分別構建營養組織和防御組織,利用植物防御產生的大量次生代謝物質“保護自己”,致癭昆蟲的“生存智慧”可見一斑。
蟲癭通常富含某些特定成分,自然界蟲癭種類極為豐富,將來如何系統性研究不同蟲癭化學成分差異,以及某些特征化合物與致癭昆蟲或寄主植物科屬之間的關系具有重要意義。同時,蟲癭中豐富的代謝產物開發利用還值得繼續深入研究,尤其是在醫藥衛生領域的開發。如五倍子蚜形成的蟲癭,不僅含有豐富的單寧酸、沒食子酸和焦性沒食子酸,其極性溶劑提取物還具有強抗氧化活性和抗菌活性(Tianetal., 2009; 查玉平等, 2016)。枸杞癭螨A.pallida蟲癭葉片中具有抗炎、降壓活性的東莨菪內酯等物質含量顯著提高(楊孟可等, 2020)。擬大枸杞瘤癭螨Aceriakuko在中華枸杞Lyciumchinense上致癭后,蟲癭葉片中綠原酸含量較非蟲癭葉片提高了36%,且90%甲醇提取液也顯示蟲癭葉片具有更高的抗氧化活性和清除1,1-二苯基-2-吡啶酰肼(DPPH)自由基能力(Chenetal., 2020)。
目前對于蟲癭的研究多集中在健康葉片和蟲癭葉片的整體比較,較少關注蟲癭部位及蟲癭周邊組織的局部比較,以及致癭過程中的動態變化,在未來的研究中應更加關注致癭過程在空間及時間上的轉錄組、代謝組等多組學的動態變化,以更深入了解致癭過程的特點及其分子機制。最后,致癭昆蟲作為生態系統工程師(ecosystem engineers),對寄主植物各個層面的影響是否會對生態系統中的其他共棲生物產生深遠的影響,致癭昆蟲的致癭行為除滿足自身所需外,是否還會延伸影響到其他生物(競爭者、天敵、植物病原菌)也是值得關注的領域。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
