乳酸菌胞外多糖生物學功能及其在畜牧生產中的應用前景
2021-05-20 01:41:12李金澤李丘軻單安山
動物營養學報 2021年4期
李金澤 李丘軻 單安山
(東北農業大學動物營養研究所,哈爾濱 2150030)
乳酸菌(lactic acid bacteria,LAB)是一類能利用可發酵碳水化合物產生大量乳酸的細菌總稱[1],主要包括乳酸桿菌、雙歧桿菌、乳酸乳球菌和鏈球菌等菌屬。在動物機體內,LAB除了可以調節腸道菌群、促進腸道蠕動、保護腸道生態平衡以及改善腸道功能外,還能提高機體免疫能力,抑制腐敗菌滋生,提高飼料轉化率[2],在畜牧業等領域中有重要的應用價值,有望成為抗生素的替代品之一[3]。大量研究顯示,LAB的上述功能可能與其細胞壁的不同成分——肽聚糖、磷壁酸、多糖、蛋白質及其次級代謝產物胞外多糖(expolysaccharides,EPS)有關(圖1)[4-5]。其中,EPS是一種分子量較高的長鏈聚合物,主要由藍藻、真菌和細菌等微生物代謝產生[6],存在形式包括構成細菌細胞壁成分的莢膜多糖,以及分泌到周圍環境中的黏多糖[7]。雖然有研究表明植物乳桿菌磷壁酸能夠抑制細胞外調節蛋白激酶(extracellular regulated protein kinases,ERK)和p38激酶的磷酸化以及抑制核因子-κB(nuclear factor-kappa B,NF-κB)的活化,從而降低細胞因子白細胞介素(interleukin,IL)中IL-8的產生[3],但是國內外相關研究主要集中在EPS上,因為EPS能夠為機體提供能量,參與機體多種生命活動,是生命有機體的重要組成成分,并具有多種生物學功能。所以,有關EPS的研究逐漸成為相關學科的前沿研究領域。EPS作為LAB的重要次級代謝產物[8],隨著研究的不斷深入,其抗腫瘤、抗氧化、抗病毒、免疫調節和腸道菌群調節等生物學功能(圖2)不斷被發現,并得到廣泛關注和認可[9-12]。本文將就乳酸菌胞外多糖(LAB-EPS)的分類、結構、生物學功能及其在生產中的應用進行綜述,為LAB-EPS能早日在生產中得到廣泛應用提供理論依據。
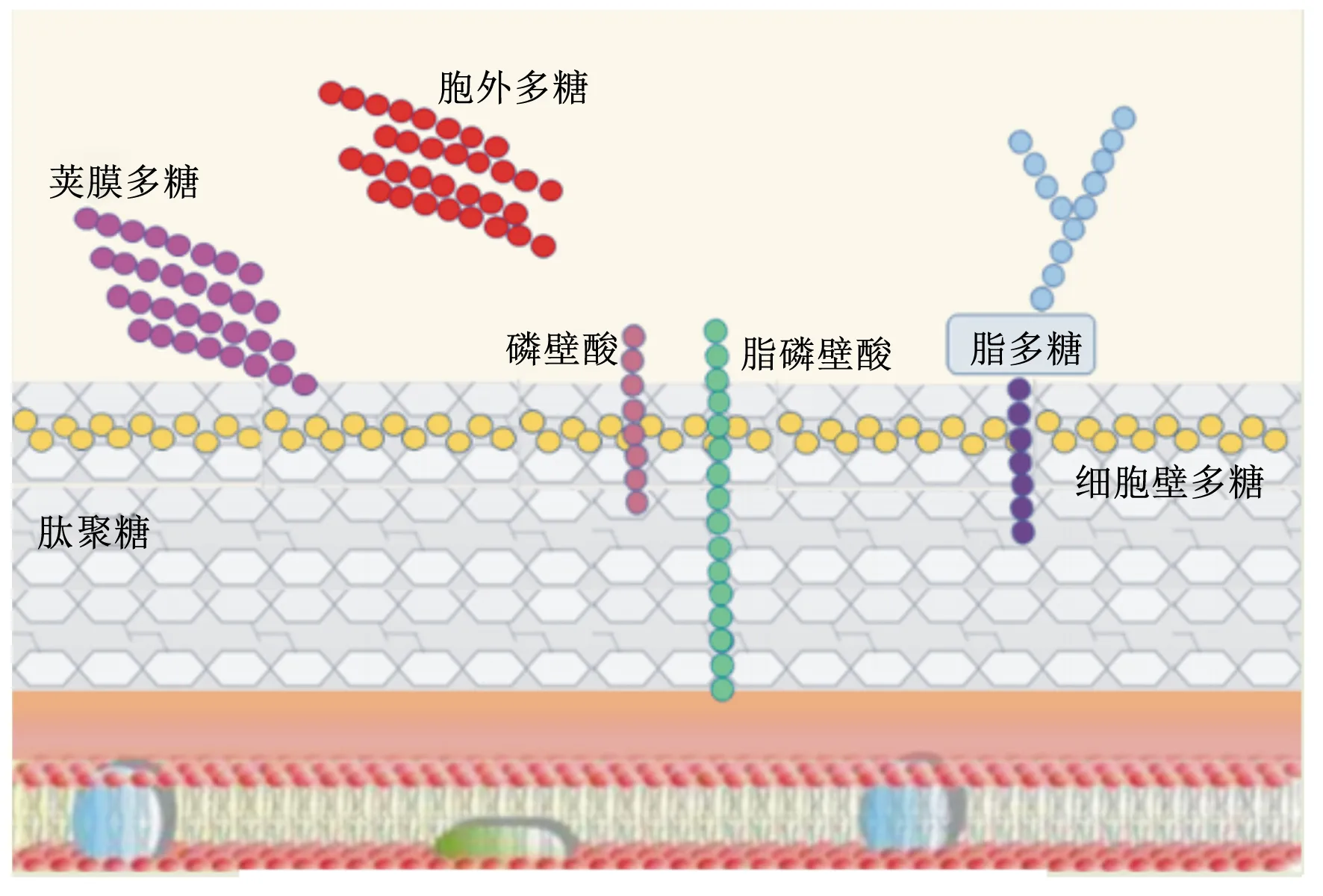
圖1 LAB細胞表面相關多糖
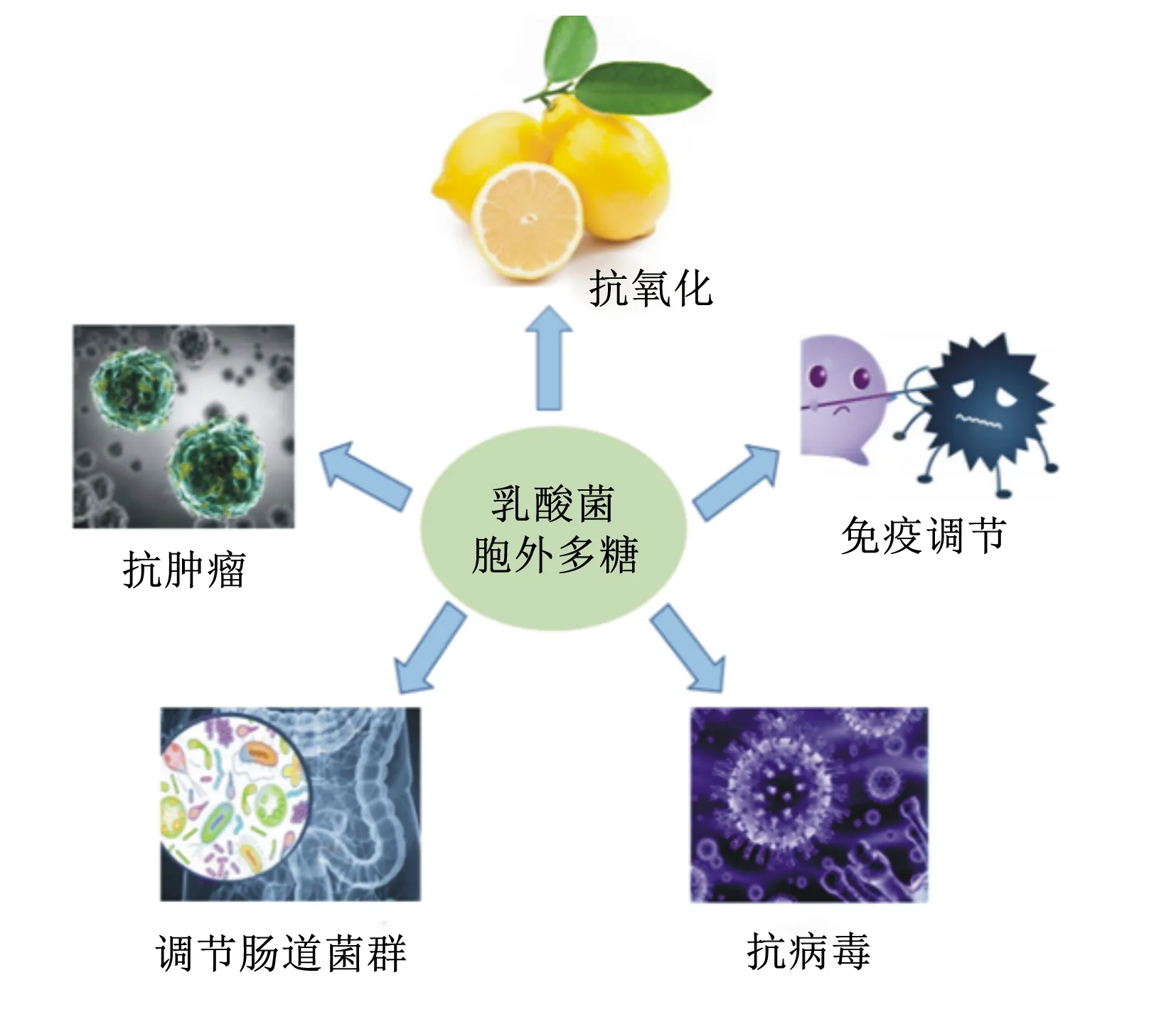
圖2 LAB-EPS主要生物學功能
1 LAB-EPS的分類及結構
與大多數其他細菌一樣,LAB能夠產生多種類型的多糖或聚糖,具有豐富的結構多樣性。EPS作為LAB的重要次級代謝產物,根據位置不同,可被分為結合在細胞表面的“莢膜多糖”和釋放到周圍環境中的“黏多糖”,由于2種多糖經常混合在一起難以分開,所以統稱為EPS。根據其化學成分的差異,EPS可分為同型多糖(homopolysaccharides,HoPS)和異型多糖(heteropolysaccharides,HePS)。HoPS是指由1種單糖脫水縮合而成的多糖;HePS是指2種及2種以上單糖通過脫水縮合形成3~8個重復單元所構成的多糖[13]。與HoPS相比,HePS包含不同單糖的重復單元(表1),主要包括不同比例的葡萄糖、半乳糖和鼠李糖,有的還包含N-乙酰基-D-氨基葡萄糖、N-乙酰基-半乳糖胺、糖醛酸或一些非碳水化合物取代基,例如丙酮酸、乙酸鹽、磷酸鹽和琥珀酸鹽等,它們在EPS發揮生物學功能時起關鍵作用[14-24],因此,HePS的生物合成及其結構更為復雜。

表1 LAB分泌的中性或酸性異型多糖的結構
2 LAB-EPS的生物學功能
2.1 抗腫瘤
抗腫瘤是LAB-EPS的主要生物學功能之一,1982年Shiomi等[25]初次提出LAB-EPS具有抗腫瘤作用之后,EPS的抗腫瘤功能便成為科研工作者的研究熱點。研究表明,LAB-EPS發揮其抗腫瘤的活性主要是通過2種方式:一是直接抑制腫瘤細胞生長;二是調動機體的免疫系統,激發免疫細胞通過先天性免疫或適應性免疫對腫瘤細胞做出反應。用嗜酸乳桿菌EPS刺激人結腸癌細胞Caco-2和小鼠,激活了關鍵轉錄蛋白NF-κB和p38絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)啟動,此外,還顯著上調了IL-1α、單核細胞趨化蛋白-1、腫瘤壞死因子-α(tumor necrosis factor α,TNF-α)和重組人穿透素-3免疫相關基因的表達,且呈現劑量依賴性變化[26]。El-Deeb等[27]發現嗜酸乳桿菌20079的EPS通過對凋亡和NF-κB通路的調節實現對Caco-2的抑制作用;嗜酸乳桿菌20079的EPS處理Caco-2后,G0/G1期凋亡細胞比例增加,細胞凋亡率可高達80.65%,同時還具有上調NF-κB抑制蛋白α、p53和轉化生長因子基因表達的潛力。Sun等[28]通過對流式細胞儀結果的分析也得到了相似的結論,嗜熱鏈球菌CH9的EPS能夠將人肝癌細胞HepG-2周期阻滯于G0/G1期。Di等[6]研究發現,干酪乳桿菌SB27的EPS可直接抑制結腸癌細胞HT-29的增殖,并上調B淋巴細胞瘤-2相關死亡啟動子(Bcl-2-associated death promoter,Bad)、B淋巴細胞瘤-2相關X蛋白(Bax)、半胱氨酸天冬氨酸蛋白酶-3(caspase-3)和半胱氨酸天冬氨酸蛋白酶-8(caspase-8)基因的表達,誘導HT-29細胞凋亡引起形態學改變。此外,副干酪乳桿菌M5的EPS也可以激活氧化應激和內質網應激通路,進而誘導HT-29細胞凋亡[29]。
2.2 抗氧化
在正常情況下,體內活性氧物質(reactive oxygen species,ROS)是有氧代謝的副產物,其中包括羥基自由基、超氧陰離子自由基和過氧化氫等[30],ROS的產生和消除維持著氧化-抗氧化平衡,這在調節信號通路的傳導和細胞增殖中起著重要作用[31]。當平衡被破壞時,ROS水平升高,導致自由基的產生,這些自由基可能對蛋白質、脂質和DNA造成有害影響[32],導致機體氧化應激,引起細胞氧化損傷并發展為多系統疾病[33]。雖然人工合成的抗氧化劑能有效減緩氧化過程,但其安全性卻受到質疑[34]。
因此,尋找具有高抗氧化活性和低細胞毒性的抗氧化劑成為研究熱點,LAB-EPS也因此備受科研人員關注[35]。目前研究顯示,大多數LAB-EPS無論在體內還是體外均具有抗氧化功能,能夠參與自由基清除,從而作為天然的安全抗氧化劑發揮作用,但是抗氧化的效果普遍低于抗壞血酸。Tang等[36]在德氏乳桿菌SRFM-1中分離出3種EPS,具有較強的超氧陰離子自由基、羥基自由基、1,1-二苯基-2-三硝基苯肼(DPPH)自由基清除活性和亞鐵離子螯合活性。植物乳桿菌KX041的EPS可以充當電子供體直接與自由基反應[37],植物乳桿菌JLAU103的EPS還能夠通過與過渡金屬離子催化劑螯合來發揮抗氧化功能[35]。劉煜珺等[38]還發現植物乳桿菌Y42的EPS發揮抗氧化功能是通過上調HT-29細胞中過氧化氫酶(CAT)、超氧化物歧化酶(SOD)和谷胱甘肽過氧化物酶(GSH-Px)等抗氧化酶系的表達量和活性實現的。此外,通過衰老模型小鼠體內試驗發現,瑞士乳桿菌的EPS在上調小鼠血清、腦組織和肝臟中多種抗氧化酶的活性和總抗氧化能力(T-AOC)的同時,還能夠下調丙二醛的水平,證實了瑞士乳桿菌的EPS在體內同樣具有抗氧化功能[39]。由此可見,LAB-EPS主要是通過清除氧自由基和提高機體內相關抗氧化酶活性實現其抗氧化功能,使其可以作為功能性飼料添加劑進行開發利用。
但是大量研究表明,菌株、培養基和培養條件等對LAB-EPS抗氧化功能均有影響[40](表2)。因此,篩選具有能夠分泌高抗氧化活性EPS的LAB菌株,通過優化培養條件等提高EPS抗氧化功能的研究也逐漸受到科研工作者的關注。

表2 部分LAB氧自由基清除能力
2.3 抗病毒
病毒的有效消除依賴于被感染細胞產生促炎性免疫反應的能力,并發展為能夠限制病毒復制的Th1型免疫。這種反應的特征是促炎細胞因子和趨化因子的產生,包括干擾素(interferon,IFN)、TNF-α和各種IL(如IL-12、IL-18及IL-23等)以及單核/巨噬細胞、自然殺傷(NK)細胞、T淋巴細胞的激活。研究表明,益生菌及其代謝物可以通過改善先天性免疫和適應性免疫來保護機體免受病毒感染,從而縮短病程,減少發病次數,減少病毒脫落,使腸道通透性正常化并增加病毒特異性抗體的產生[49]。Sirichokchatchawan等[10]研究證實,植物乳桿菌培養液上清液和活菌均可以在非洲綠猴腎細胞(Vero細胞)上抑制豬流行性腹瀉病毒感染和復制,并且上清液抗病毒活性具有明顯劑量和細菌數量依賴性。Kim等[50]證明了植物乳桿菌LRCC5310的EPS通過減少腹瀉持續時間,抑制腸上皮病變,減少輪狀病毒在腸道的復制,縮短哺乳小鼠的恢復時間,表現出明顯的抗輪狀病毒活性。Kanmani等[49]研究發現,德氏乳桿菌的EPS具有改善腸道抗病毒反應和防止腸道病毒(如輪狀病毒)感染的潛力。此外,德氏乳桿菌TUA4408L的EPS還可以通過減少病毒復制來提高豬腸上皮細胞對輪狀病毒感染的抵抗力,并通過調節Toll樣受體3(Toll-like receptor 3,TLR3)激活抗病毒先天免疫應答[51]。王瑩瑩[52]發現,嗜酸乳桿菌的EPS可以促進豬睪丸細胞的免疫活性,抑制豬傳染性胃腸炎病毒感染,在作用早期EPS可以協同豬傳染性胃腸炎病毒誘導豬睪丸細胞的TLR3表達和NF-κB的活化,上調IL-6、IL-8和IFN-γ的表達,并調節細胞的先天性免疫反應,提高細胞的抗病毒能力。由此可見,LAB-EPS不僅能夠抑制病毒的感染和復制,還可以通過激活動物機體先天性免疫應答和適應性免疫應答發揮抗病毒功能,在畜牧生產中具有重要的研究價值。
2.4 免疫調節
LAB-EPS是一種天然的大分子活性物質,安全性高且來源的可追溯性強,在調節機體免疫過程中具有良好的免疫原性,是理想的天然免疫佐劑。研究發現,大多數LAB及其EPS可通過提高單核細胞吞噬能力,促進T、B淋巴細胞的增殖,提高NK細胞殺腫瘤活性和促有絲分裂活性,誘導細胞因子來調節機體免疫系統,從而提高機體的免疫防御能力來抵抗病原體。Zhu等[53]通過體外試驗證明,植物乳桿菌RS20D分泌的EPS可以刺激巨噬細胞釋放一氧化氮(NO),并在mRNA水平上調促炎細胞因子的基因表達。此外,EPS除了可以通過提高胞內酶活性及釋放活性氧激活巨噬細胞RAW264.7,發揮免疫增強作用[54],還能顯著提高巨噬細胞RAW264.7的增殖和吞噬活性,并誘導NO、TNF-α、IL-1β和IL-6的產生[5]。由副干酪乳桿菌DG產生的EPS還能夠增強人單核細胞系THP-1中TNF-α、IL-6、IL-8和巨噬細胞炎癥蛋白3α的表達[55]。綜上所述,大多數LAB-EPS主要是通過調節免疫相關基因,激活巨噬細胞,刺激淋巴細胞增殖并釋放NO、IL和TNF-α等來表現免疫調節活性[56]。因此,無論是體外試驗還是體內試驗均顯示出LAB-EPS具有良好的免疫調節作用,很多LAB-EPS發揮免疫調節作用的同時也具有一定的抗腫瘤和抗病毒作用。由此推測,LAB-EPS很可能是通過增強機體免疫力,促進淋巴細胞、巨噬細胞等增殖,合成分泌相應細胞因子,從而抑制腫瘤細胞生長,提高動物機體抵抗病毒感染能力,發揮抗腫瘤、抗病毒作用。
2.5 調節腸道菌群
動物腸道中聚居著1013~1014個微生物,是一個復雜的微生態體系。腸道微生物的組成與機體健康密切相關,在維持生理平衡中起著至關重要的作用。同時,腸道微生物的構建與腸道免疫系統的發育也有密不可分的聯系,大多數LAB-EPS能促進腸道有益微生物的生長,改善其多樣性[57]。EPS可以作為腸道微生物的碳源,因為腸道微生物的基因組編碼碳水化合物活性酶(carbohydrate active enzymes,CAZymes),而人和其他哺乳動物的基因組則沒有編碼足夠的CAZymes,因此,腸道微生物能夠將EPS降解為短鏈脂肪酸,如乙酸、丙酸和丁酸等[58]。這些短鏈脂肪酸對于維持腸上皮功能、控制腸上皮生長、刺激免疫系統、預防結直腸癌并減少各種炎癥性疾病有重要的作用[59](圖3)。有研究顯示,LAB-EPS可能通過抑制細菌細胞表面的特異性黏附因子來減少有害菌對腸上皮細胞的黏附[48],選擇性促進有益菌的增殖,抑制有害菌的增殖和有害物質的產生[60],進而改善腸道微生物的多樣性和平衡[57]。大量體外研究結果顯示,LAB-EPS可以有效抑制阪崎腸桿菌、大腸桿菌、單核細胞增生李斯特菌、金黃色葡萄球菌、白色念珠菌、鼠疫桿菌和鼠傷寒沙門氏菌等有害菌的生長[61-64],其中有些EPS可以同時抑制多種有害菌的增殖,具有一定的廣譜抗菌性。邵麗[65]研究表明,鼠李糖乳桿菌KF5的EPS可以部分被人的糞便菌群利用,促進雙歧桿菌的增殖,改變糞便菌群的組成,提高菌群的豐度,促進短鏈脂肪酸產生,具有潛在的益生特性。李勝杰[66]用雙歧桿菌WBIN03的EPS對小鼠進行灌胃,發現EPS可以促進乳桿菌和厭氧總菌的生長,并抑制腸道內腸桿菌、腸球菌及擬桿菌的生長,進一步證明EPS是一種潛在的益生物質。
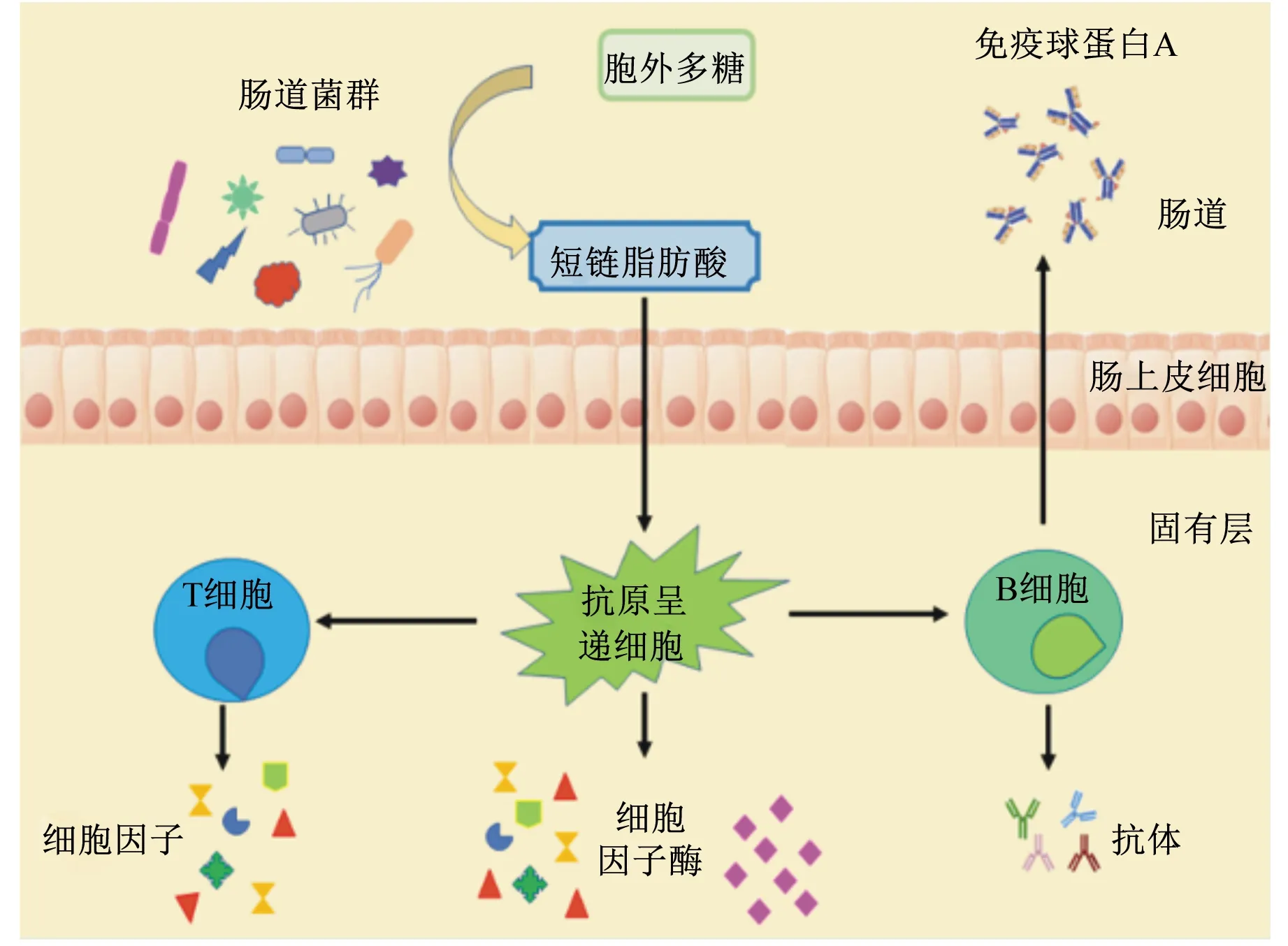
圖3 LAB-EPS與腸道微生物的相互作用
2.6 其他生物學功能
通過上述研究發現,每種LAB-EPS生物學功能并不是單一的,同一種EPS可能具有不同的功能,不同的EPS可能具有相同的作用機制。除了上述生物學功能以外,研究發現,EPS對細菌生物膜形成有明顯抑制作用,并通過減弱細胞表面修飾或減少細胞間相互作用以抑制細菌細胞的初始自動聚集和細胞附著,從而可以用于治療和預防由產生生物膜的致病細菌引起的傳染病[67]。植物乳桿菌BGCG11產生的EPS通過下調IL-1β和誘導型一氧化氮合酶(iNOS)mRNA以及增加具有抗炎活性的IL-6和IL-10細胞因子的水平,在大鼠中表現出較高抗炎活性[68]。EPS還能與病原菌細胞壁相互作用,降解細胞壁和質膜引起蛋白質溶解和必需物質的外露,從而導致病原菌死亡[69]。Ai等[70]發現干酪乳桿菌LC2W的EPS能顯著降低自發性高血壓大鼠的收縮壓,馬乳酒樣乳桿菌WT-2B產生的EPS可以通過抑制血管緊張素Ⅰ轉換酶的活性來降低血壓[71]。在體外試驗中,植物乳桿菌BR2的EPS可以降低45%的膽固醇水平[72]。
3 LAB-EPS在畜牧生產中的應用
EPS作為LAB的重要次級代謝產物之一,隨著研究不斷深入,其各種生物學功能逐漸被人們所熟知,所以,目前已經廣泛應用于農業、食品、醫藥和化妝品等研究領域。在食品工業中,EPS可以改善食品品質[73],提高酸奶的黏度[74]、奶酪產量[75]和冰淇淋的黏性及假塑性[76]。此外,EPS還能阻礙淀粉的凝沉并降低陳化速率,從而延長面包的保質期,提高面包質量[77]。因此,EPS具有非常可靠的安全性。
在過去幾十年中,為了追求較高的經濟效益,抗生素被大量廣泛地應用于畜牧生產中,正是由于抗生素的濫用,導致環境污染、畜禽產品藥物殘留及耐藥菌株出現[78-79]等問題越來越嚴重。2020年我國已全面禁止抗生素在飼料添加劑中使用,隨之可能會出現動物生產性能降低、病死率增加等問題,所以,開發抗生素替代品作為飼料添加劑以保證畜禽健康生長成為研究熱點[80]。EPS作為LAB的重要次級代謝產物,具有多種生物學功能,是非常有潛力的抗生素替代物之一[81]。研究發現,在飼糧中添加LAB能夠下調IL-6、IL-8及TNF-α的表達和分泌,顯著增加仔豬防御肽pBD2、PG1-5和pBD2的分泌表達,還能增強腸上皮細胞防御功能并通過調節腸道菌群來增強腸道屏障作用[81]。而LAB可以通過EPS與腸上皮細胞的黏附定殖在腸道中,EPS還可以促進LAB生長,提高其抗逆性及益生潛力[82],并形成保護層幫助LAB逃避免疫監測,促進腸道內LAB的定殖,同時還能降低腸道致病菌檸檬酸桿菌的定植水平[83]。此外,EPS還能夠通過競爭性抑制機制干擾細菌表面上特定黏附因子與腸道黏膜表面受體結合,從而降低細菌致病性[26]。據報道,飼喂能夠產生EPS的羅氏乳桿菌發酵的飼糧,可降低斷奶仔豬回腸、盲腸和結腸中產腸毒素的大腸桿菌的定殖水平[84],有效緩解因腸道菌群平衡破壞導致的疾病。此外,Ashfaq等[85]將EPS添加到雛雞飼糧中,發現EPS能夠顯著提高腸道內LAB的豐度,并抑制常見腸道病原體如大腸桿菌、沙門氏菌和腸球菌的生長,具有替代抗生素的潛力。Rajoka等[86]對從母雞盲腸內分離的LAB所產生的EPS進行體外試驗研究,EPS表現出較強的抗大腸桿菌和鼠傷寒沙門氏菌活性,對羥基自由基、DPPH自由基和超氧陰離子自由基有較強的清除能力,同時還能抑制Caco-2的生長,表現出一定的抗腫瘤活性。由此推斷,LAB-EPS通過調節腸道菌群結構、抑制有害菌定殖及調節免疫反應等作用,提高機體抗病能力,從而減少抗生素在生產中的應用。
4 小 結
與動物和植物來源的EPS相比,LAB產生的EPS具有許多優點,例如具有較強的可操作性、較高的繁殖能力和高級性能等,但到目前為止,其結構和活性關系尚未完全清楚。因此,我們需要對EPS的結構和活性關系進行深入研究,并構建其潛在生物學功能和工業應用之間的橋梁。目前,EPS應用研究主要集中于食品生產等領域,雖然也有研究顯示,在畜牧業中LAB-EPS可以通過其抑菌活性、免疫增強作用、促進營養物質消化吸收和調節腸道菌群等方面的作用進而維持動物腸道的微生態平衡,提高動物的免疫力及生產性能,但是相關研究多數集中在體外試驗及小鼠體內試驗。有關EPS應用于畜禽的研究并不多,其中主要原因集中在LAB-EPS產率較低,無法滿足畜禽體內試驗所需劑量。另外,不同LAB分泌的EPS生物學功能也存在較大差異,其作用機理等也均處于初步研究階段尚不明確。所以,關于如何提高LAB-EPS的產量和生物學功能,及其在生產實踐中的應用和作用機理等研究均有廣闊的前景,同時LAB-EPS也有望成為抗生素替代物之一。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
汽車工程學報(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年2期)2017-02-21 01:21:24
國際漢語學報(2016年1期)2017-01-20 08:21:20
