水稻苗期抗稻瘟病基因全基因組關聯分析
2021-05-15 09:27:02陳汝斌彭沙莎康厚祥
激光生物學報 2021年2期
關鍵詞:水稻
陳汝斌,彭沙莎,王 丹*,康厚祥
(1. 湖南農業大學農學院,長沙 410128;2. 中國農業科學院植物保護研究所/植物病蟲害生物學國家重點實驗室,北京 100193)
水稻(Oryzae sativaL.)是世界上最重要的糧食作物之一,養育了世界近一半以上的人口[1]。據研究,到2050 年,平均每年需要增加44 萬t 以上的糧食產量以滿足全球的糧食需求[2]。由稻瘟菌(Magnaporthe oryzae)引起的稻瘟病是一種長期侵害水稻的真菌病害,幾乎發生在水稻生長發育的整個時期[3]。根據侵染部位可將其分為苗瘟、葉瘟、穗頸瘟、節瘟以及谷粒瘟等類型[4]。每年因稻瘟病造成的水稻收成損失可達到10%~30%,嚴重時達50%以上,甚至顆粒無收,給我國的糧食生產及稻米品質造成了嚴重的損失。目前針對稻瘟病發生的防止措施是化學藥劑防治以及培育優良廣譜抗性品種。相比于前者,后者具有經濟、環保、高效等特點[5]。然而,由于田間稻瘟菌生理小種基數大、傳播和變異速度快等原因,已有抗病基因的水稻其抗性在推廣3~5 年后容易喪失,因此快速挖掘新的稻瘟病抗性基因對培育抗稻瘟病品種具有重大的意義[6]。
目前,定位復雜性狀主要有兩種方式,一是經典的連鎖分析,二是關聯分析。連鎖分析基于雙親雜交構建遺傳作圖群體,通過遺傳標記對個體進行基因分型,并對結果進行分析。但該方法存在構建群體耗費時間長且每個位點只能鑒定2 個等位基因等缺點,因此所鑒定數量性狀位點(quantitative trait locus,QTL)的數量受到限制[7]。而關聯分析相較于連鎖分析有以下幾個優點:1)目標群體為自然群體,不需要耗費大量時間構建雙親作圖群體,極大地節約了時間和降低了工作強度;2)可同時鑒定多個等位基因。全基因組測序技術的不斷發展推動了全基因組關聯分析(genome-wide association study,GWAS)技術用于遺傳作圖的進程。GWAS 又名連鎖不平衡作圖,可快速對QTL 進行精細定位,且分析通量較高[8]。該方法在研究水稻的農藝性狀改良中被廣泛應用。如Lu 等[9]利用469 份水稻種質資源定位了17 個水稻劍葉長度相關的QTL;Huang等[10]從950 份水稻品種中定位到10 個與產量性狀相關的QTL;Jiang 等[11]鑒定出350 份水稻品種中23 個與水稻分蘗相關的QTL。近幾年GWAS 也被應用于定位抗病相關位點。如Zhang 等[12]用6 個白葉枯小種對172 份群體材料進行接種,通過GWAS鑒定了12 個白葉枯抗性相關位點;Kang 等[13]用5 個稻瘟菌接種了390 份群體材料,鑒定出97 個稻瘟病抗性相關位點;Liu 等[14]通過GWAS 鑒定了27 個稻瘟病抗性相關位點,并成功克隆了一個稻瘟病抗性基因PiPR1。GWAS 方法的廣泛應用極大程度上提高了育種的效率及精度,為后續抗性品種選育提供了一定的科學基礎。
為進一步發掘新的稻瘟病抗性基因,本研究對水稻多樣性群體II(rice diversity group II,RDP-II)中470 份水稻種質資源進行了稻瘟病抗性評估,并使用高密度陣列[700 000 個單核苷酸多態性(single nucleotide polymorphism,SNP)]進行了基因分型[15],選擇最優的關聯分析模型,并對定位的稻瘟病抗性相關的QTL 進行了分析,為快速發掘新的稻瘟病抗性基因提供了科學依據。
1 材料與方法
1.1 試驗材料與田間種植
本研究所用的470 份水稻材料來自RDP-II,由廣東省農業科學院劉斌老師提供。于2018 年5 月將育好的秧苗種植于湖南省益陽市桃江縣的自然病圃中(N28°22',E112°03'),以每行20 株種植24 行,株行距為15 cm×24 cm。該病圃面積約0.133 km2,歷年來稻瘟病發生較為嚴重。此外,為保證發病質量,本研究在群體材料周圍種植了高感稻瘟病的品種進行自然誘發。在生長階段應注意水培管理,多施加氮肥以誘導稻瘟病的發生,待水稻苗三葉一心時期(2018年6月)進行葉瘟調查。
1.2 表型調查及分析
根據稻瘟病分級標準對自然病圃中群體材料進行調查[13],其中0 級(高抗)為無病斑;1 級(高抗)為僅有針尖大小病斑;2 級(抗)為出現較大褐點;3 級(中抗)為直徑1~2 mm 的小圓斑;4~6 級(中感)開始形成典型的病斑,占葉面積大小約6%~25%;7 級(感)的病斑面積超過25%;8~9 級(高感)的病斑面積達到50%以上,甚至整片葉片死亡。利用Excel 2010 進行表型數據的處理,過濾掉不正常數據后,以2 次重復數據的平均值對亞群表型及抗性品種分布等結果進行分析。
1.3 基因型檢測與分析
利用Mccouch 等[15]報道的700 000 個SNP 芯片對470 份水稻種質資源進行基因型分析,在Tassel 4.0 軟件中過濾掉低質量的SNP 位點,選擇最小等位基因頻率(minor allele frequency,MAF)>0.05 以及缺失率≥20%的SNP 位點,以進行下一步的關聯分析。
1.4 GWAS 數據分析
本研究利用700 000 個SNP 基因型數據以及來自RDP-II 群體的470 份水稻材料的抗病表型數據,選擇混合線性模型(mixed linear model,MLM),即同時加入親緣關系Kinship 以及群體結構Q 矩陣,在Tassel 4.0軟件中進行GWAS。利用R軟件(R version 3.6.2)的qqman包繪制曼哈頓圖。
1.5 抗病相關QTL的定位
通過GWAS,我們得到176 個與稻瘟病抗性相關聯的SNP[-log10(Pvalue)≥3.50]。為了降低假陽性,我們以在200 kb 的基因組間隔內具有至少2 個顯著相關的SNP 位點為標準定位抗病相關的QTL。在國家水稻數據中心(http://www.ricedata.cn/)及NCBI 數據庫(https://www.ncbi.nlm.nih.gov/)中尋找已克隆或定位的稻瘟病抗性相關基因的位置信息,與本研究所檢測的QTL進行共定位。
2 結果與分析
2.1 自然群體抗病表型分析
本研究根據國際水稻所稻瘟病0~9 級分級標準,對湖南省桃江縣自然病圃中(圖1)的470 份水稻品種進行了稻瘟病抗性的評估,其中0~1 級為高抗、2 級為抗、3 級為中抗、4~6 級為中感、7 級為感、8~9級為高感。
抗病表型的統計結果如表1 所示。抗病品種占自然群體的32.98%,而感病品種占自然群體67.02%,表明該群體總體趨勢上對稻瘟菌表現為感病。

圖1 湖南省桃江縣稻瘟病自然病圃Fig. 1 Natural blast nursery in Taojiang county, Hunan province
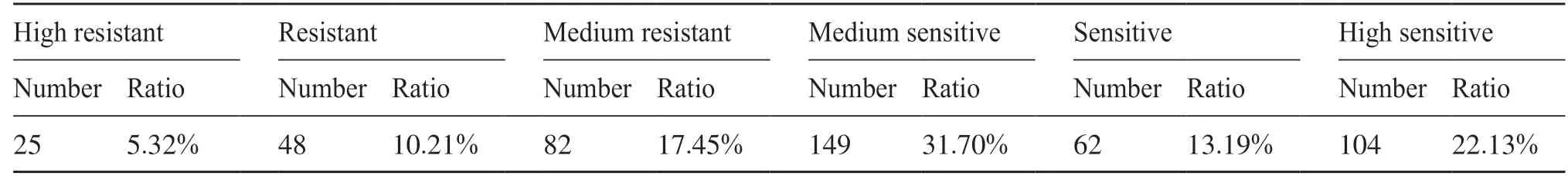
表1 RDP-II群體中抗病表型分布Tab. 1 The distribution of resistance phenotype in RDP-II population
2.2 群體中亞群抗病表型分析
本研究所用群體材料除6 份未定義品種外,可大致分為6 個亞群,其中秈稻(indica,IND)163 份、香型稻(aromatic,ARO)65份、熱帶粳稻(tropical ja-ponica,TRJ)102份、溫帶粳稻(temperate japonica,TEJ)35份、混合型水稻(admixture,ADM)28份,奧斯稻(aus,AUS)71 份。從亞群抗病表型直方圖中(圖2a)我們發現,熱帶粳稻亞群的平均抗性水平最高,達到4.0,而溫帶粳稻亞群的平均抗性水平最低,達到6.0,秈稻亞群的平均抗性水平居中,為5.3。氣泡圖如圖2b 所示。氣泡自上而下分別代表1~9 抗病等級,氣泡大小代表品種數目。通過分析發現,在抗病等級≤3(表型表現為抗病)時,熱帶粳稻亞群中含有56個符合條件的品種,而溫帶粳稻亞群僅含有8個品種。該結果進一步證明了以上結論。
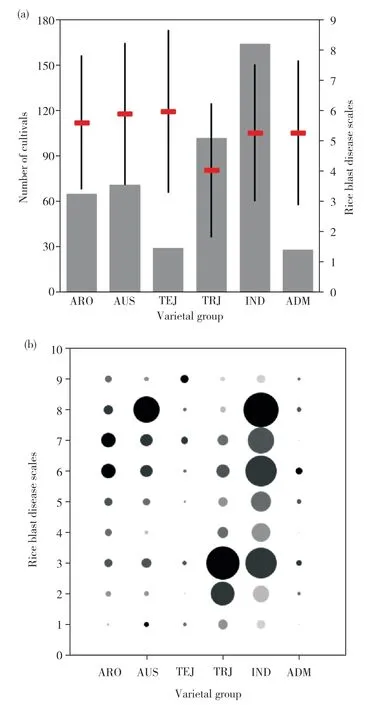
圖2 RDP-II群體的亞群抗病表型Fig. 2 Subpopulations resistance phenotype of RDP-II
通過對470 份水稻群體材料的表型數據進行統計發現,在73份抗性品種(0~2級)中,熱帶粳稻亞群的品種最多,占總數的38%,而溫帶粳稻亞群的品種僅占6%。該結果同群體材料中亞群稻瘟病抗性分布較為一致。此外,表2 特別列出了超高抗的25 個品種(0~1 級),其分別來自亞洲(56%)、非洲(24%)、北美洲(16%)以及歐洲(4%)這4個不同的大洲。這些超高抗材料為抗病育種提供了優良的種質資源。
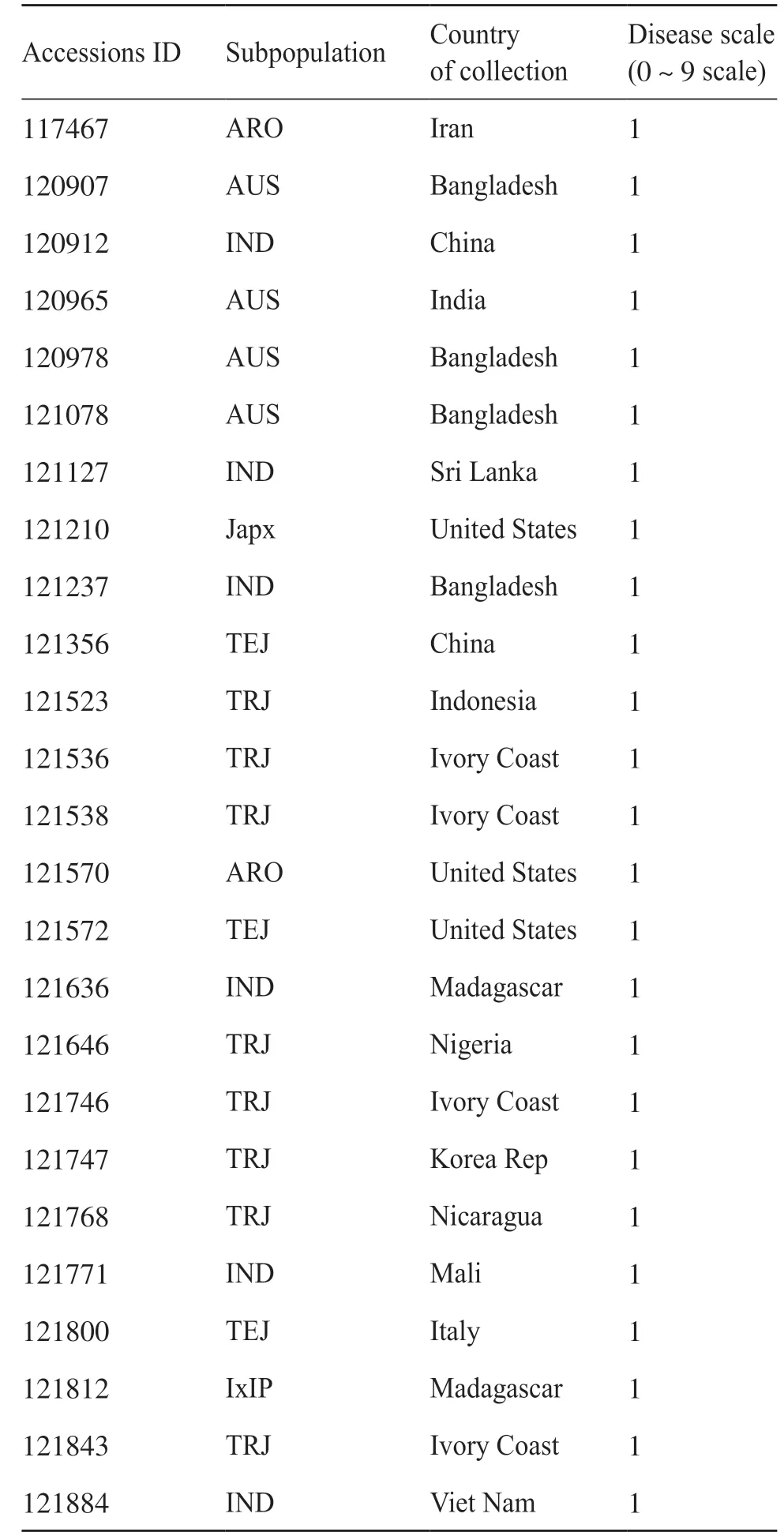
表2 高抗品種亞群及地區分布Tab. 2 Subpopulation and regional distribution of high resistant varieties
2.3 GWAS
本研究基于470 份水稻自然群體材料的抗病表型數據以及700 000 個SNP 基因型數據, 選擇MLM模型進行抗病表型數據和SNP 標記的GWAS。在閾值[-log10(Pvalue)≥3.50]時判定SNP 標記與抗病表型關聯的顯著性,共檢測到176 個相關聯的SNP位點,覆蓋于全部染色體。抗病表型關聯分析結果如圖3 所示,曼哈頓圖中紅色虛線以上代表抗病相關性顯著的位點。
2.4 稻瘟病抗性關聯位點鑒定
以SNP 上下游200 kb 范圍內至少2 個顯著相關SNP 位點作為1 個QTL,對GWAS 結果所檢測的176 個抗病相關SNP 進行QTL 分析。由表3 可知,在除了4 號及7 號外的10 條染色體上定位到24 個稻瘟病抗性相關位點(loci associated with rice blast resistance,LABRs)。6 號染色體上分布最多,為4 個,其中最關聯的SNP 標記(SNP-6.8792603)也位于6 號染色體上。將這些檢測的LABRs 同已克隆或定位的稻瘟病抗性相關基因進行共定位,結果共定位到4 個已克隆稻瘟病抗性基因和2 個已定位的基因,分別是1 號染色體的OsSGT1[16]、2 號染色體的OsCBSX3[17]和OsCUL3a[18]、11 號染色體的Pi-38[19]、12 號染色體的Pi-ta[20]和Pi-42[21]。分析發現,這4 個基因中除了OsCUL3a為負調控因子外,其余均為抗病反應的正調控因子。
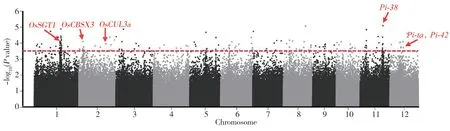
圖3 稻瘟病抗病表型全基因組關聯分析曼哈頓圖Fig. 3 Genome-wide association analysis manhattan plots of rice blast resistance
3 討論
稻瘟病抗性機制復雜,既受主效基因的控制,又受QTL 的影響。由主效基因介導的抗性是定性的,對稻瘟菌具有完全的抗性,而由多個QTL 介導的抗性是定量的,對稻瘟菌只表現部分抗性[22]。廣泛的實踐證明,單一主效抗性基因在3~5年內易失去抗性[23],因此快速挖掘新的稻瘟病抗性基因意義重大。本研究對470 份水稻種質資源進行了水稻苗期稻瘟病抗性評估,其中熱帶粳稻亞群的平均抗性水平最高,而溫帶粳稻亞群的平均抗性水平最低,秈稻亞群的平均抗性水平居中。該結論與之前對RDP-I 和RDP-II 群體的表型研究結果一致[13-14]。同時對高抗品種的亞群分布進行研究,發現來自熱帶粳稻亞群最多,這與群體分布結果相一致。此外,高抗品種中一半以上來自亞洲(56%),可能與亞洲地區的稻瘟病抗性選擇較高有關。本研究所獲得的高抗品種為育種家提供了新的抗稻瘟病種質資源。
本研究選擇混合線性模型MLM 對抗病表型數據及700 000 個SNP 基因型數據進行GWAS,共檢測到分布于12 條染色體上的176 個相關聯SNP位點。通過對這些SNP 位點進行QTL 分析發現,在除了4 號及7 號外的10 條染色體上定位到24 個LABRs,-log10(Pvalue)值介于3.72~5.10 之間,可以解釋3.37%~45.06%的表型變異。將這些LABRs同已克隆或定位的基因進行共定位,分析發現,5 個位點(LABR2、LABR4、LABR6、LABR22、LABR23)共定位了6 個稻瘟病抗性相關基因,包括4 個已克隆及2 個已定位的基因,分別位于1、2、11 及12 號染色體上。這5 個位點對應的峰值SNP 分別為SNP-1.25004091、SNP-2.3910977、SNP-2.31170935、SNP-11.21704502、SNP-12.10723430。除上述5 個位點與已報道的位點共定位外,其余19 個顯著關聯位點為首次發現。這些新位點的發現對后續稻瘟病抗性基因的發掘以及進一步的優良品種選育工作具有重要意義。基于以上研究結果,下一步的研究工作將主要集中于候選基因的選擇以及稻瘟病抗性基因的克隆與功能驗證,為培育廣譜抗性水稻品種提供理論基礎。

表3 24個稻瘟病抗性顯著相關位點Tab. 3 24 significant LABRs
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
