FDFT1基因重組慢病毒表達載體的構建及其表達測定
2021-05-13 01:14:26陳曉穎易雪麗陸飛燕曾怡
右江民族醫學院學報 2021年2期
陳曉穎,易雪麗,,陸飛燕,曾怡
(1. 右江民族醫學院附屬醫院,廣西 百色 533000;2. 右江民族醫學院病原微生物學與免疫學教研室,廣西 百色 533000)
法尼基二磷酸法尼基轉移酶1(farnesyl diphosphate farnesyl transferase 1,FDFT1) ,又稱鯊烯合成酶,是一種分子量為47 kDa的酶,由416個氨基酸組成。人類FDFT1啟動子包含兩個固醇調節元件(sterol regulatory element,SRE)SRE-1基序、一個倒置的SRE-3 (inverse SRE-3,Inv SRE-3)和一個核轉錄因子Y(nuclear factor,NF-Y),是SRE結合蛋白(binding protein,BP)SREBPs的結合位點。這三個SREBP結合位點已經被證實與甾醇介導的FDFT1啟動子的調控有關[1]。SREBPs以轉錄因子形式參與膽固醇生物合成途徑的許多基因的轉錄。SREBP2是FDFT1的關鍵轉錄因子之一。SREBP2位于內質網膜上,當SREBP2被激活后進入細胞核,與FDFT1啟動子中的SRE區域結合,激活FDFT1進入轉錄過程。FDFT1是膽固醇合成途徑的第一個酶,通過兩階段反應合成角鯊烯。兩個焦磷酸法尼酯(farnesyl pyrophosphate,FPP)分子先聚合形成前角鯊烯二磷酸(pre-squalene diphosphate,PSDP), PSDP再被NADPH還原,生成角鯊烯[2],角鯊烯通過氧化、還原等多步驟轉化為膽固醇。有研究結果表明膽固醇代謝異常與癌癥的發生有關[3],因此,與膽固醇代謝相關的酶和相關的生物分子被認為可能是抗腫瘤治療的靶點[4],靶向膽固醇代謝治療方法備受重視。FDFT1在膽固醇合成調節中起重要作用,因在肺癌、髓系白血病、乳腺癌等腫瘤中高表達[5]而備受關注。本實驗通過構建FDFT1基因重組慢病毒表達載體,為進一步探索FDFT1基因在腫瘤中的作用機制。
1 材料與方法
1.1 材料 包膜質粒pMD2.G、包裝質粒psPAX2、pCDH-GFP質粒載體為南京醫科大學盧春教授饋贈。PCR引物、PrimeScriptTMII 1st Strand cDNA Synthesis Kit、PCR試劑盒、DL 2,000 DNA Marker、 DL 10,000 DNA Marker、DL 15,000 DNA Marker、DNA切膠回收試劑盒、real-time PCR (qPCR)試劑盒為Takara生物公司(大連)產品;內切酶XbaⅠ、BamHⅠ購自NEB公司;質粒小量提取試劑盒、EasyGeno重組試劑盒、DH5α感受態細胞為北京天根生化科技有限公司產品;胎牛血清、DMEM購自Gibco公司; Trizol試劑為卓一生物技術有限公司(南寧)產品;lipofectamine2000購自Invitrogen公司。293T細胞株為本實驗室保存。
1.2 方法
1.2.1 FDFT1基因片段的擴增 根據NCBI登記的FDFT1基因序列設計PCR引物,由Takara生物公司合成,引物序列如下:
FDFT1-上游引物(FDFT1-F):GGACCGCAGAGGTGAGAGTCGC
FDFT1-下游引物(FDFT1-R):TTCCTAAAGGTCCCAGCCACAC
以293T細胞cDNA為模板,普通PCR進行擴增,得到FDFT1基因全長片段。50 μl PCR反應體系包括模板1 μl,TaKaRa LA Taq 0.5 μl,2× GC Buffer 25 μl,dNTP Mixture 8 μl,上游引物0.5 μl,下游引物0.5 μl,ddH2O 14.5 μl。PCR反應條件如下:94℃預變性3 min;94℃變性30 s、60℃退火30 s、72℃延伸150 s,共進行35個循環;之后72℃加長延伸5 min。PCR產物經瓊脂糖凝膠電泳確定擴增陽性后,-20℃保存。
設計FDFT1基因PCR引物,在上下游加入與pCDH-GFP載體同源序列(“”部分)及XbaⅠ、BamHⅠ限制性酶切位點(單下劃線斜體部分),并在上游引物起始密碼子前增加KOZAK序列(“”部分)增強目的基因表達,在下游引物終止密碼子前增加V5標簽蛋白編碼序列(雙下劃線斜體部分)。引物序列如下:
FDFT1-重組上游引物(FDFT1-重組F):5′-CCTCCATAGAAGATTCTAGAGCCACCATGGAG
TTCGTGAAATGCCTTGGCCACCCCGAAGAGTT
CTACAACCTGGTGCGCTTC-3′
FDFT1-重組下游引物(FDFT1-重組R):5′-TCCTTCGCGGCCGCGGATCCTTACGTAGAATCGAG
ACCGAGGAGAGGGTTAGGGATAGGCTTACCATGTTCTCCAGTCTGAACATAG-3′
以前述FDFT1基因PCR擴增產物為模板再次擴增,反應體系及反應條件同前述,使FDFT1基因片段上下游分別帶有與載體互補序列及限制性酶切位點。純化擴增產物。測定DNA濃度,-20℃保存備用。
1.2.2 pCDH-GFP線性化載體的制備 用限制性內切酶XbaⅠ、BamHⅠ酶切pCDH-GFP載體。雙酶切反應體系如下:pCDH-GFP質粒載體1 μg,XbaⅠ 1 μl,BamHⅠ 1 μl,Cutsmart Buffer 5 μl,ddH2O補足50 μl;雙酶切條件:37℃ 水浴15 min;65℃水浴20 min。回收酶切后的pCDH-GFP載體,并測定其濃度。純化的pCDH-GFP線性化載體-20℃保存備用。
1.2.3 FDFT1基因重組質粒的構建及鑒定 將線性化載體與目的基因通過同源重組反應連接。10 μl反應體系如下:線性化載體0.01 pmol,插入片段0.05 pmol,2×EasyGeno Assembly Mix 5 μl,以ddH2O補足10 μl;反應條件如下:50℃反應15 min,迅速冰上冷卻5 min。將上述反應液進行轉化,轉化體系如下:50 μl DH5α感受態細胞;5 μl上述重組產物。轉化條件如下:冰浴30 min;42℃熱激90 s后冰浴150 s;加入350 μl的 LB液體培養基37℃ 180 r/min搖45 min。取100 μl轉化液涂布于事先配好的含氨芐青霉素(Amp)的LB固體平板,37℃ 180 r/min倒置培養16~18 h。挑取單菌落接種至分裝好的Amp抗性的LB液體培養瓶并標記,37℃ 180 r/min振蕩培養60 min。取1 μl菌液進行菌落PCR鑒定,PCR反應體系及條件同1.2.1。將菌落PCR陽性的菌液繼續培養至16 h后提取質粒。將提取質粒進行雙酶切驗證,雙酶切體系及條件同1.2.2。對菌落PCR、雙酶切均符合的質粒(命名為pCDH-GFP-FDFT1)進行序列測定(廣州艾基生物有限公司)。根據實驗需要將序列比對正確的質粒進行批量提取,-20℃保存備用。
1.2.4 FDFT1慢病毒的包裝 用胰酶消化293T細胞,離心棄上清后用10%FBS的完培將細胞調整為1×105cells/ml,取2 ml接種至6孔板中,37℃、5% CO2培養24 h。轉染前吸棄上清用預冷PBS洗細胞2次并盡量吸凈,并用無血清DMEM潤洗1次備用。將提取的pCDH-GFP-FDFT1、pCDH-GFP分別與包裝質粒psPAX2 和包膜質粒pMD2.G 利用lipofectamine2000共轉染293T細胞。轉染8 h后給細胞換液,培養至48 h后觀察熒光蛋白GFP表達情況并收集上清4℃保存。加入新鮮完培繼續培養24 h后觀察熒光蛋白GFP表達情況并收集上清。將48 h和72 h的上清混合離心后用0.45 μm濾器過濾,分裝病毒液,-80℃保存。
1.2.5 FDFT1慢病毒的表達測定 按照實驗所需將293T細胞用10%FBS的完培調整為1×105細胞/孔,接種至6孔板,37℃、5% CO2培養24 h。將實驗所需的病毒液置于冰上并在4℃融化備用。感染前用預冷PBS洗細胞2次并盡可能吸干凈,隨機分組并標記,對應加入200 μl的FDFT1病毒液(FDFT1-VIR)及陰性對照病毒液(Ctrl-VIR),用無血清的DMEM補足2 ml。感染8 h后以10%的完全培養基給細胞換液。96 h觀察熒光蛋白GFP表達,并用Trizol試劑分別提取細胞RNA,以PrimeScriptTMⅡ 1st Strand cDNA Synthesis Kit逆轉錄為cDNA后進行qPCR,運用相對定量法比較兩組樣本FDFT1表達情況。
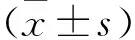
2 結果
2.1 FDFT1目的基因的擴增 以FDFT1-F、FDFT1-R為引物,普通PCR從人全基因組cDNA中擴增FDFT1全長基因并進行瓊脂糖核酸電泳,電泳結果顯示在約1200 bp處出現單一條帶,與實驗預期的FDFT1目的條帶1254 bp基本一致,見圖1。以該FDFT1 PCR產物為模板,以FDFT1-重組F、FDFT1-重組R引物再次進行PCR擴增,使FDFT1兩端加入載體同源序列及限制性酶切位點,經瓊脂糖核酸電泳,結果顯示在約1300 bp處出現目的條帶,與預期的1342 bp基本一致(圖片未顯示)。切膠回收純化帶載體同源序列的FDFT1 PCR擴增產物,得到濃度為23.30 ng/μl的FDFT1目的基因DNA。
注:M:DL2,000 DNA Marker;1:FDFT1全基因組片段擴增產物。
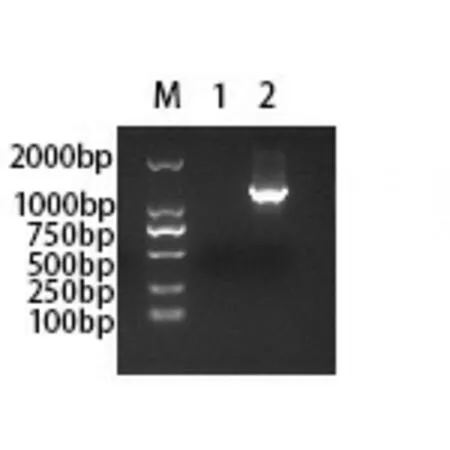
2.2 pCDH-GFP線性化載體制備 用限制性內切酶酶切pCDH-GFP質粒,酶切后進行電泳驗證酶切情況。如圖2所示,在約7500 bp處呈現單一條帶(預期條帶為7544 bp),說明線性化載體制備成功。切膠回收酶切后pCDH-GFP載體,得到濃度為19.04 ng/μl的線性載體DNA。
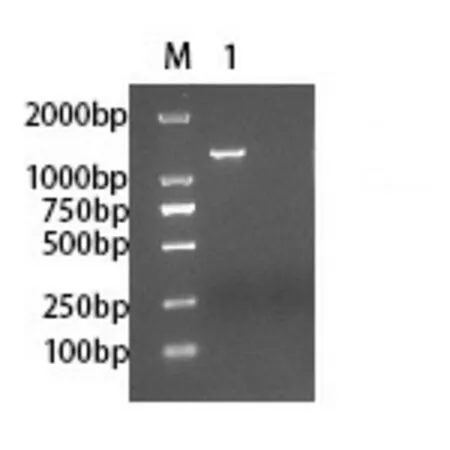
2.3 FDFT1基因重組質粒的構建及鑒定 利用同源重組技術將FDFT1目的基因片段連接到載體上,經轉化涂布接種含Amp的LB平板,培養16~18 h后挑2個陽性克隆,分別轉種至含Amp抗性的LB液體培養瓶。1 h后取菌液進行菌落PCR鑒定,如圖3所示,只有2號菌落在約1200 bp處出現條帶。培養至16~18 h,提取菌落PCR陽性菌液的質粒。質粒進行雙酶切驗證,如圖4所示,酶切后的質粒出現兩條條帶,在約1300 bp及7500 bp處,說明目的基因重組成功。將菌落PCR、雙酶切驗證均符合的重組質粒進行測序(廣州艾基生物有限公司),測序結果顯示重組質粒含有目的序列,與GenBank中登記的FDFT1(NM_004462)基因同源,表明FDFT1基因重組質粒連接成功。

注:M:DL15,000 DNA Marker;1:pCDH-GFP質粒雙酶切產物。
注:M:DL2,000 DNA Marker;1:1號菌落PCR擴增產物;2:2號菌落PCR擴增產物。
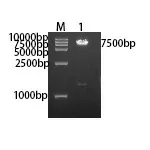
注:M:DL15,000 DNA Marker;1:pCDH-GFP-FDFT1質粒雙酶切產物。
2.4 FDFT1慢病毒包裝 轉染48 h后利用熒光顯微鏡觀察熒光蛋白GFP表達。如圖5所示,pCDH-GFP組及pCDH-GFP-FDFT1組均可看到綠色熒光,表明構建的質粒成功轉染293T細胞。
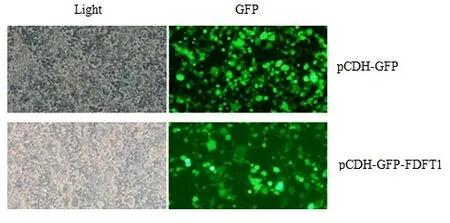
圖5 熒光顯微鏡觀察三質粒共轉染48 h的293T細胞(×100)
2.5 FDFT1慢病毒表達測定 同體積的Ctrl-VIR及FDFT1-VIR分別感染293T細胞96 h后,兩組均有綠色熒光表達,表明包裝的病毒顆粒成功感染293T細胞。分別提取細胞總RNA,以逆轉錄試劑進行逆轉錄為cDNA,利用qPCR檢測FDFT1表達,結果如表1所示,兩組間差異有統計學意義(P<0.001),FDFT1-VIR組相對Ctrl-VIR組上調15.13倍。

表1 FDFT1慢病毒感染293T細胞后FDFT1表達水平
3 討論
膽固醇是構成細胞質膜的重要部分,是維持細胞內穩態的必要脂質。膽固醇穩態[6]是一個復雜的調控過程,它在膽固醇的合成、酯化、代謝和運輸中起作用。近年來膽固醇生物合成的變化被認為是多種癌癥[7]的標志。當癌細胞快速增殖時,它們依賴于升高的膽固醇生物合成來提供足夠的膽固醇,以滿足膜生物生成和能量穩態及類固醇激素合成。膽固醇也可以通過脂質筏間接的發揮促癌作用,通過富膽固醇的脂筏[8]來傳導腫瘤細胞的增殖、遷移和生存信號。也有研究發現,膽固醇通過在TME中富集的膽固醇酯和氧甾醇等衍生物來調節其對腫瘤進展的影響[9]。FDFT1即法尼基二磷酸法尼基轉移酶1是一種膜結合酶,位于染色體8p.22-23.1區[10],參與膽固醇合成分支的調節。FDFT1可能作為癌癥的潛在靶點在癌癥的治療中發揮作用[11]。Hughes SJ等[12]通過Northern blot分析和免疫組化分析發現在8p22-23區的FDFT1 mRNA過表達,在食管腺癌中表達增加。但Weng ML等[13]研究發現在結直腸癌中FDFT1表達較低,FDFT1作為抑癌基因通過負性調控AKT/mTOR/HIF1α信號通路來實現腫瘤抑制功能。FDFT1既作為一種致癌基因又是一種抑制基因與癌癥密切相關,目前關于這種差異產生的原因的研究還很少。這種差異可能不僅是由于FDFT1表達的變化,而且是由于癌癥中各種膽固醇代謝物如何導致不同反應的差異,這也可能是TME對FDFT1表現出不同反應的結果。關于FDFT1影響癌癥特征的作用有很多報道,然而,關于與FDFT1相互作用的分子的研究卻很少。雖然已經有很多關于FDFT1抑制劑的研究,但大多數研究都集中在降低血液膽固醇上,似乎還需要更多的研究將其用作抗癌藥物。我們期望能夠通過構建FDFT1慢病毒穩轉株以研究FDFT1在各種腫瘤的發生發展中的相關機制,為癌癥的治療提供依據。
傳統的脂質體轉染方法存在一定的弊端,不適用于轉染效率不高的細胞,慢病毒載體雖然操作復雜,但從理論上講病毒顆粒能感染所有細胞,并能將外源基因整合到宿主染色體上,可以獲得穩定表達的細胞株,比傳統的脂質體轉染發更具優勢[14]。腺病毒載體雖然病毒滴度高,且易于轉染其他細胞,但仍不能克服表達沉默的自身缺陷[15]。慢病毒載體是以病原體HIV-1 為基礎發展起來的基因治療載體,該載體可以將外源基因隨機插入宿主基因組,具有持久性表達,感染效率高、安全性高和免疫原性低等優勢[16]。雖然與其他載體相比,慢病毒載體更勝一籌,但其生物安全性方面的局限性是我們所關注的。近年來,研究者們也主要著力于改進其基因結構,從而增加慢病毒載體的生物安全性[17]。在本實驗中,我們通過包裝產生的病毒為“自殺”性病毒,病毒在感染靶細胞后不會產生新的病毒顆粒,也不會再去感染其他細胞。本實驗成功構建FDFT1慢病毒載體,經轉染后獲得的病毒液能感染293T細胞,并表達FDFT1基因。本文結果為進一步研究FDFT1基因在腫瘤中的作用機制提供了實驗基礎。
