碳基納米銅復(fù)合材料對普通小球藻脅迫作用的研究
2021-05-10 13:01:26吉喜燕唐靜懿葉璟吳世超黃賽花侯梅芳李鑫許文武
生態(tài)環(huán)境學(xué)報 2021年3期
關(guān)鍵詞:復(fù)合材料
吉喜燕,唐靜懿,葉璟,吳世超,黃賽花,侯梅芳, ,李鑫,許文武
1. 上海應(yīng)用技術(shù)大學(xué)生態(tài)技術(shù)與工程學(xué)院,上海 201418;2. 廣東省科學(xué)院生態(tài)環(huán)境與土壤研究所/華南土壤污染控制與修復(fù)國家地方聯(lián)合工程研究中心/廣東省農(nóng)業(yè)環(huán)境綜合治理重點實驗室,廣東 廣州 510650;3. 上海應(yīng)用技術(shù)大學(xué)化學(xué)與環(huán)境工程學(xué)院 上海 201418;4. 上海應(yīng)用技術(shù)大學(xué)軌道交通學(xué)院 上海 201418
納米材料在醫(yī)學(xué)、生物等領(lǐng)域的廣泛應(yīng)用使其不可避免地進(jìn)入各種環(huán)境介質(zhì)之中,其中水環(huán)境是納米類材料的主要受納介質(zhì)(Chen et al.,2017)。不僅單質(zhì)類納米材料對水環(huán)境中生物的影響備受人們關(guān)注,碳基納米復(fù)合材料對水生態(tài)的影響也成為了研究熱點之一,碳基納米銅就是其中的典型代表之一(Gao et al.,2020)。當(dāng)碳基納米銅進(jìn)入水環(huán)境介質(zhì)后會對水體中的藻類產(chǎn)生一定的影響(Windfeld et al.,2013)。由于普通小球藻具有普適性強、生長力旺盛等特點,因此常用于水生生物毒理學(xué)實驗(孫紅羽等,2019)。
據(jù)報道,碳基納米銅對藻類的毒性作用與其他納米單質(zhì)或納米氧化物相似,碳基納米銅復(fù)合材料暴露下的藻細(xì)胞會被納米材料包裹,形態(tài)會發(fā)生明顯變化(劉雅等,2016)。當(dāng)碳基納米銅濃度超過30 mg·L-1時,普通小球藻細(xì)胞中活性氧(ROS)的水平升高,超氧化物歧化酶(SOD)活性增強,藻細(xì)胞的生物量降低,細(xì)胞活性下降(Costa et al.,2020)。碳基納米銅懸浮液等水相碳基復(fù)合材料的毒性在不同水生生物體中存在差異,有研究者推斷,這是由于碳基納米銅對不同水生生物細(xì)胞表面的作用存在差異而導(dǎo)致的(Eltarahony et al.,2018)。不同濃度的碳基納米銅復(fù)合材料對藻細(xì)胞的數(shù)量、葉綠素 a含量等具有明顯的影響,當(dāng)碳基納米銅材料濃度范圍為5—10 mg·L-1時,碳基納米銅的存在對蛋白核小球藻(Chlorella pyrenoidosa)和斜生柵藻(Scenedesmus obliquus)的葉綠素a含量具有促進(jìn)作用;而同等濃度下的單一碳粉則對二者的葉綠素 a含量無明顯影響(Fazelian et al.,2019)。對于蛋白核小球藻而言,當(dāng)碳基納米銅的濃度高于60 mg·L-1時,其對藻細(xì)胞生物量的增長出現(xiàn)了嚴(yán)重的抑制現(xiàn)象,藻細(xì)胞的比增長速率幾乎為0(Mahawar et al.,2018)。據(jù)文獻(xiàn)報道,實際河、湖水體中碳基納米銅的水平最高可達(dá)約60 mg·L-1,水體中的碳基納米銅復(fù)合材料除了會對藻細(xì)胞的生物量、形態(tài)、葉綠素a含量等造成一定的影響,還會對藻細(xì)胞的生理代謝過程也會產(chǎn)生擾動作用(Dimkpa al.,2012;Lammel et al.,2019)。以往關(guān)于碳基納米氧化鋅對小球藻的生物毒性的研究中,發(fā)現(xiàn)低濃度的碳基納米氧化鋅對小球藻的生理活性具有抑制作用,小球藻基因的完整性會受到碳基氧化鋅破壞并出現(xiàn)氧化應(yīng)激反應(yīng)(Alho et al.,2020)。上述關(guān)于納米類、碳基納米類材料的藻細(xì)胞生理毒性研究中,并未涉及到藻細(xì)胞的代謝物豐度及相關(guān)代謝途徑的分析,尤其是碳基納米銅對普通小球藻代謝方面的脅迫作用的研究仍然較少。
為此,本文以普通小球藻為研究對象,從藻細(xì)胞葉綠素a含量和藻細(xì)胞代謝組學(xué)特性變化兩個方面入手,研究不同濃度碳基納米銅復(fù)合材料(0、0.05、0.5、5、50 mg·L-1)對普通小球藻的脅迫作用。本文旨在探究碳基納米銅復(fù)合材料脅迫下普通小球藻的差異代謝物豐度和對應(yīng)顯著富集通路的變化,豐富碳基納米材料對藻細(xì)胞脅迫作用相關(guān)研究的數(shù)據(jù)和資料,為碳基納米類復(fù)合材料與水生生態(tài)關(guān)系的研究提供科學(xué)參考。
1 材料與方法
1.1 實驗設(shè)計
本文實驗設(shè)計包括普通小球藻葉綠素a含量變化和藻細(xì)胞代謝組學(xué)變化兩部分,設(shè)置碳基納米銅的濃度為0、0.05、0.5、5、50 mg·L-1。實驗裝置如圖1所示。
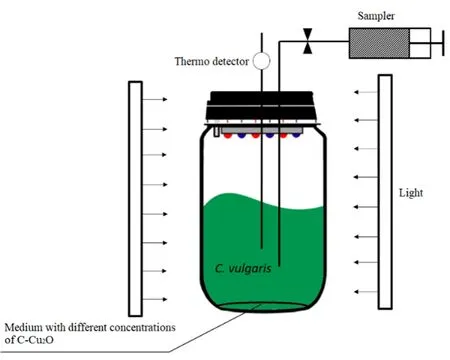
圖1 實驗裝置圖Fig. 1 The setup of experiment
如圖1所示,實驗裝置置于無菌培養(yǎng)箱中,由反應(yīng)器(250 mL反應(yīng)瓶)、光源、采樣管3個主要部分組成。錐形瓶中含有 150 mL人工成合成的具有不同濃度碳基納米銅的BG-11培養(yǎng)基。每個實驗組及對照組均設(shè)置平行組(n=3,n為每個平行組的數(shù)量)。調(diào)節(jié)光照為 120 μmol·(m2·s)-1,溫度為 28 ℃,藻細(xì)胞初始濃度為 1×105cell·mL-1。
1.2 藻種培養(yǎng)
本研究選用普通小球藻作為受試對象進(jìn)行實驗,所用的藻種購買自中國科學(xué)院武漢水生所淡水藻種庫,編號FACHB-8。藻種在光照培養(yǎng)箱中進(jìn)行培養(yǎng)及擴培,選取BG-11培養(yǎng)基,恒溫28 ℃,設(shè)置光暗比為 12 h∶12 h,光照強度維持在 120 μmol·(m2·s)-1。BG-11 培養(yǎng)基成分列于表1。

表1 BG-11培養(yǎng)基成分Table 1 The BG-11 medium components
將實驗室擴培的藻及菌用于實驗之前,需將普通小球藻和地衣芽孢桿菌分別以 8000×g離心 10 min并棄去上清液。用 0.1 mol·L-1的 PBS(PBS,Phosphate buffered solution)緩沖液(無菌超純水配置)重復(fù)離心、洗滌3次,以確保藻類粘連的培養(yǎng)基去除干凈才可正式接種于實驗中培養(yǎng)體系中。
1.3 主要指標(biāo)檢測與表征
1.3.1 普通小球藻葉綠素a濃度檢測
首先,將10 mL藻菌混合物樣品進(jìn)行離心處理,轉(zhuǎn)速為4000×g,離心時間10 min,離心后棄去上清液。將所得沉淀溶于 10 mL體積分?jǐn)?shù) 90%的丙酮中,將樣品置于旋渦震蕩儀混勻。然后置于4 ℃黑暗環(huán)境中處理24 h。將樣品取出后再次進(jìn)行離心處理,離心轉(zhuǎn)速為4000×g,離心時間15 min。離心后的上清液用于葉綠素a濃度測定。紫外分光光度法測定波長為630、645、663、750 nm的吸光度,以90%丙酮溶液作為空白對照。葉綠素 a質(zhì)量濃度(μg·L-1)通過式 1 計算(Ptushenko et al.,2017):

式中,Chl-a表示葉綠素質(zhì)量濃度,單位為μg·L-1;OD663、OD750、OD645、OD630分別為對應(yīng)波長下樣品測得的吸光度數(shù)值;V為樣品體積,單位為mL。
1.3.2 普通小球藻代謝組學(xué)檢測
取適量樣品,加入500 μL甲醇-水(4∶1)溶液,低溫下采用高通量組織破碎儀進(jìn)行破碎;加入 200 μL氯仿,冰浴超聲萃取30 min,4 ℃、12000 g條件下離心20 min,取上清液,真空抽干;加入80 μL的甲氧胺鹽酸鹽吡啶溶液,于 37 ℃肟化反應(yīng) 90 min;加入 80 μL 的 BSTFA(含 1%TMCS)和 20 μL的正己烷,70 ℃反應(yīng)60 min,進(jìn)行氣相色譜-質(zhì)譜聯(lián)用(安捷倫8890B-5977B)分析。樣品經(jīng)HP-5MS毛細(xì)管柱(30 m×0.25 mm×0.25 μm)分離后進(jìn)入質(zhì)譜,全掃描模式,質(zhì)量掃描范圍為m/z50-500。在GC-MS檢測分析的過程中,采用質(zhì)控樣品評價整個分析系統(tǒng)的穩(wěn)定性。
采用 MassHunter軟件進(jìn)行預(yù)處理,同時利用Fiehn數(shù)據(jù)庫對代謝物進(jìn)行搜庫鑒定,導(dǎo)出包含代謝物名稱、保留時間、質(zhì)核比和峰面積等的數(shù)據(jù)矩陣。采用偏最小二乘法判別分析PLS-DA分析來區(qū)分各組間代謝輪廓的總體差異,變量權(quán)重VIP值大于1,同時單因素方差分析顯著性分析P<0.05(FDR校正)的代謝物被認(rèn)為是發(fā)生顯著變化的差異代謝物。采用 KEGG通路數(shù)據(jù)庫(http://www.genome.jp/kegg)、MetaboAnalyst(https://www.metaboanalyst.ca/faces/home.xhtml)對差異代謝物進(jìn)行代謝通路分析,獲得差異代謝物的生物功能、顯著富集的代謝通路等信息。
1.4 數(shù)據(jù)分析
本文使用 GraphPad Prisim進(jìn)行單因素分析(ANOVA,Analysis of Variance)。代謝物分析使用SIMCA-P 14.0軟件進(jìn)行偏最小二乘判別分析(PLSDA,Partial Least Squares Discriminant Analysis)。使用MeV軟件(4.8.1版)繪制代謝物相對含量的熱圖。本文中圖主要使用Graphpad Prisim繪制。
2 結(jié)果與討論
2.1 小球藻葉綠素a含量變化分析
葉綠素是光合作用的物質(zhì)基礎(chǔ),光合作用是影響小球藻細(xì)胞生長的重要因素,葉綠素的含量驟降會導(dǎo)致小球藻光合作用效率降低,因此,在碳基納米銅復(fù)合材料暴露下普通小球藻細(xì)胞內(nèi)葉綠素a的降低將直接影響小球藻的光合作用和生長狀態(tài)(Luimstra et al.,2019;朱津輝等,2019)。對實驗周期內(nèi)不同濃度碳基納米銅體系第2、4、6、8天的葉綠素a含量進(jìn)行了檢測,結(jié)果如圖2所示。
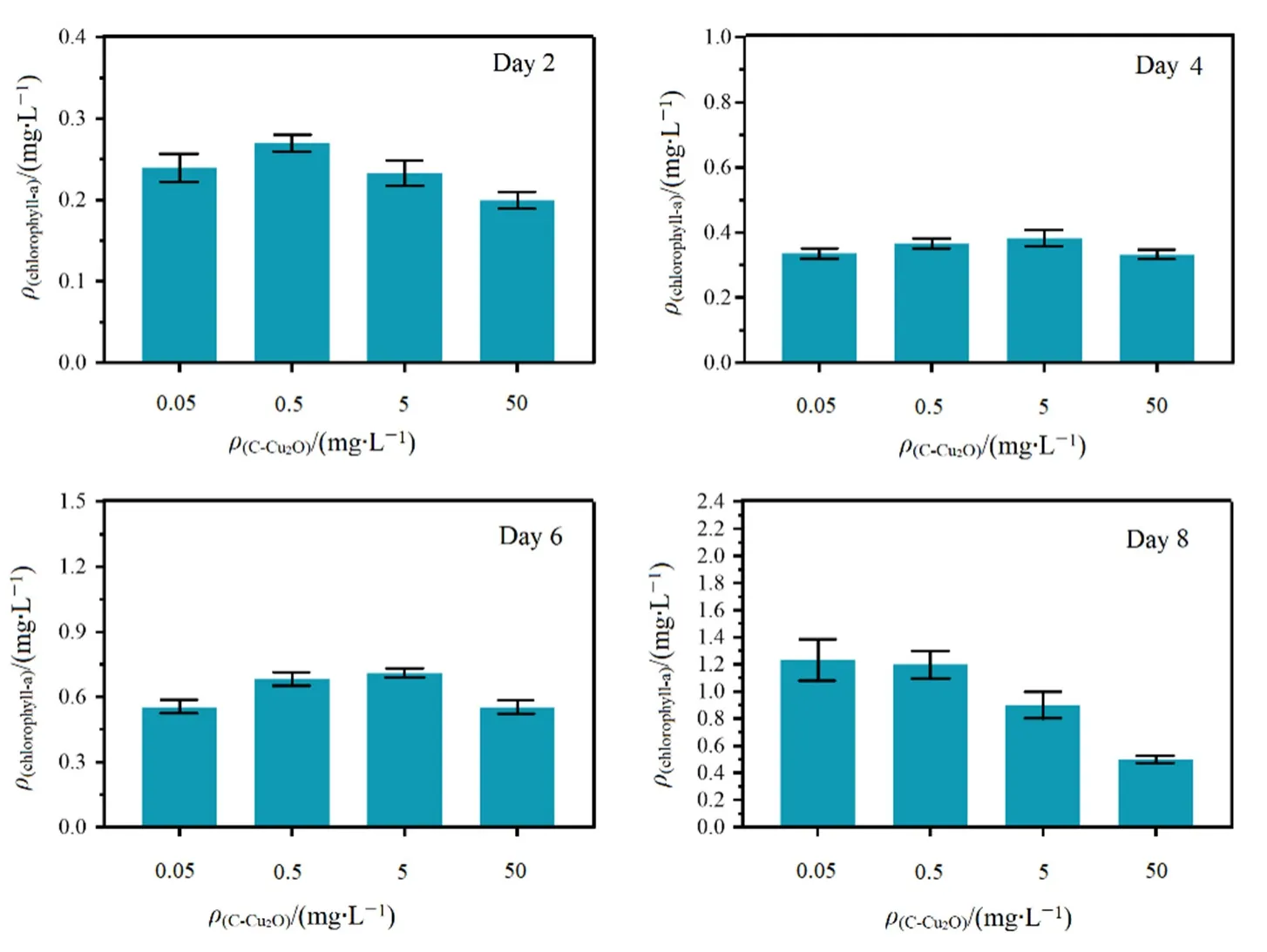
圖2 不同濃度碳基納米銅濃度條件下普通小球藻的葉綠素a含量變化Fig. 2 The chlorophyll-a content of C. vulgaris under different concentrations of C-Cu2O
由圖2可知,0.05—5 mg·L-1碳基納米銅暴露條件時,普通小球藻葉綠素a含量逐漸增加,這表明低濃度碳基納米銅對普通小球藻葉綠素a含量變化的影響并不明顯。50 mg·L-1碳基納米銅暴露條件下的普通小球藻葉綠素 a含量隨時間的推移明顯降低,這表明高濃度碳基納米銅對普通小球藻葉綠素a含量有明顯的抑制作用。這與以往研究中低濃度碳基納米銅(5—10 mg·L-1)對普通小球藻葉綠素a含量并無明顯影響而高于20 mg·L-1的濃度時,碳基納米銅對普通小球藻葉綠素a含量產(chǎn)生抑制作用的結(jié)果一致(Fazelian et al.,2019)。
2.2 碳基納米銅迫條件下的普通小球藻代謝組學(xué)特性變化分析
2.2.1 不同濃度碳基納米銅條件下普通小球藻的差異代謝物分析
不同濃度碳基納米銅材料會對普通小球藻的差異代謝物產(chǎn)生一定的影響,碳基納米銅的濃度越高,對代謝物種類的影響也越大,實際自然水體中檢測到的碳基納米銅的濃度范圍為0.5—50 mg·L-1(Alho et al.,2020;Miao et al.,2020)。在實驗周期的第 8天,觀察發(fā)現(xiàn) 5 mg·L-1和 50 mg·L-1實驗組中,葉綠素a濃度變化差異最為顯著,而葉綠素a的產(chǎn)生是普通小球藻細(xì)胞內(nèi)代謝調(diào)控的結(jié)果,基于此,對 5 mg·L-1和50 mg·L-1實驗組進(jìn)行代謝組學(xué)檢測,以驗證碳基納米銅復(fù)合材料對普通小球藻胞內(nèi)代謝所產(chǎn)生的影響。對各濃度碳基納米銅體系中的普通小球藻進(jìn)行洗滌,進(jìn)行代謝組學(xué)測試分析。對5 mg·L-1和 50 mg·L-1碳基納米銅體系及空白對照組體系中的普通小球藻代謝輪廓進(jìn)行 PCA(Principal component analysis)分析,結(jié)果如圖3A所示,5 mg·L-1碳基納米銅組與對照組代謝輪廓有部分交叉,而50 mg·L-1碳基納米銅組則距離二者較遠(yuǎn)。此結(jié)果表明,50 mg·L-1碳基納米銅體系中的普通小球藻出現(xiàn)嚴(yán)重的代謝紊亂,代謝輪廓發(fā)生明顯偏移,而5 mg·L-1碳基納米銅則未能明顯擾動普通小球藻的代謝。這表明50 mg·L-1碳基納米銅對普通小球藻的代謝特性產(chǎn)生了較為明顯的影響。相較之下,5 mg·L-1碳基納米銅則對普通小球藻胞內(nèi)代謝物變化的影響不明顯。造成這種現(xiàn)象的原因是由于普通小球藻本身對外界脅迫具有較強的耐受性,相較之下,同屬綠藻門的斜生柵藻則對碳基納米銅復(fù)合材料更為敏感,當(dāng)受到水體環(huán)境中同等濃度碳基納米銅復(fù)合材料的脅迫時,其代謝輪廓更容易發(fā)生變化,會比普通小球藻更易發(fā)生較為明顯的質(zhì)壁分離現(xiàn)象,同時,斜生柵藻也比普通小球藻更易內(nèi)吞水體環(huán)境中的碳基納米銅復(fù)合材料,從而造成胞內(nèi)細(xì)胞器的損傷(Landa et al.,2017;Wang et al.,2020)。
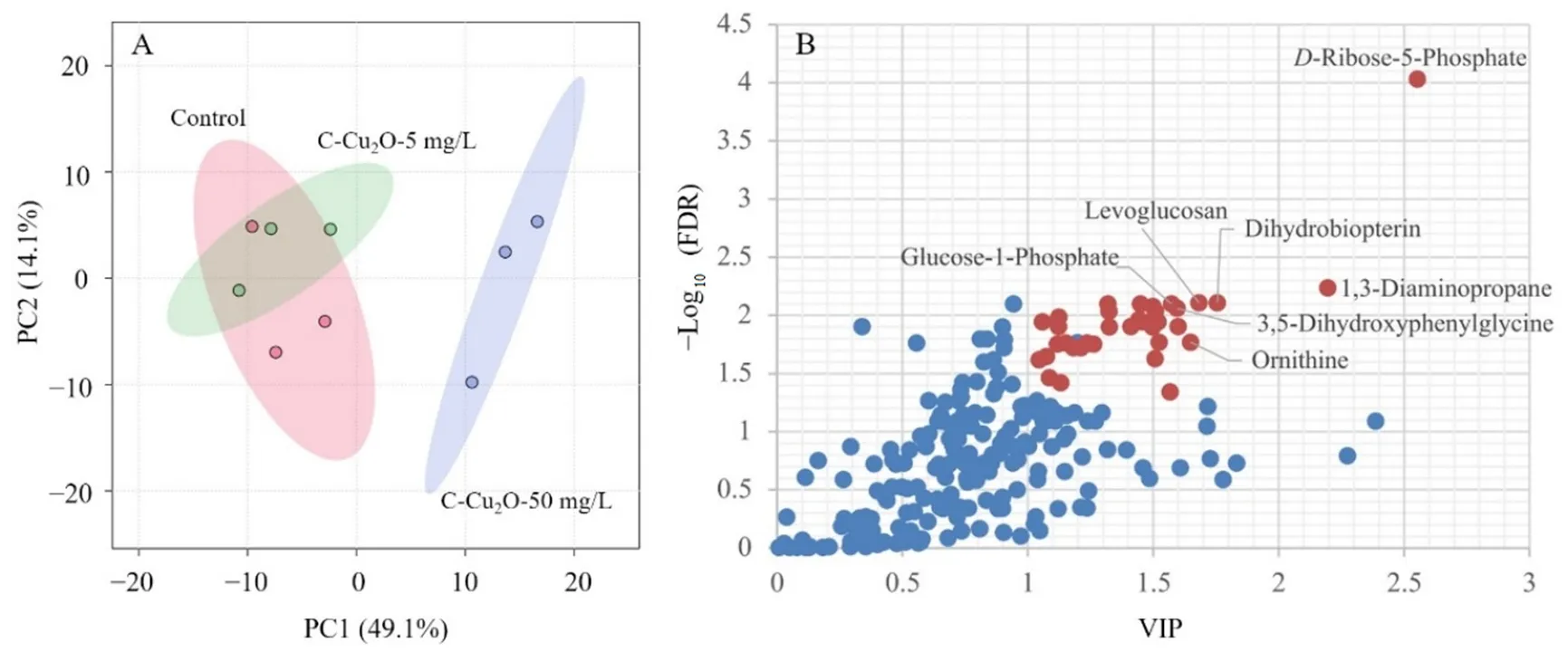
圖3 不同濃度碳基納米銅材料體系中藻細(xì)胞代謝物主成分分析Fig. 3 PCA of C. vulgaris metabolites in C-Cu2O nanoparticles system with different concentrations
為了進(jìn)一步有效地找出與高濃度碳基納米銅相關(guān)的內(nèi)源代謝物,進(jìn)一步對普通小球藻代謝輪廓數(shù)據(jù)進(jìn)行PLS-DA分析,獲得能直觀反映3組間代謝差異貢獻(xiàn)率的VIP圖(圖3B)。通過對相關(guān)數(shù)據(jù)進(jìn)行獨立樣本ANOVA檢驗,篩選VIP值大于1且P值小于0.05(FDR校正)的差異代謝物,通過HMDB、KEGG和MetPA等數(shù)據(jù)平臺進(jìn)行檢索,最終確定了如表2所示的36種差異代謝物。

表2 空白對照組與5 mg·L-1、50 mg·L-1碳基納米銅體系中普通小球藻的差異代謝物Table 2 Different metabolites of C. vulgaris in control group and 5 mg·L-1, 50 mg·L-1 C-Cu2O nanoparticles
表2中,碳基納米銅與空白對照組間代謝物相對含量升高,用“↑”標(biāo)記;代謝物相對含量降低,用“↓”標(biāo)記。與對照組比,5 mg·L-1和 50 mg·L-1碳基納米銅體系中,左旋葡聚糖(Levoglucosan)、葡萄糖-磷酸(Glucose-Phosphate)顯著上調(diào);其中,5 mg·L-1碳基納米銅體系中,哌啶酸(Pipecolic Acid)、氧代戊二酸(Oxoglutaric acid)顯著上調(diào)。與空白對照組相比,5 mg·L-1和 50 mg·L-1碳基納米銅體系中,1, 3二氨基丙烷(1, 3-Diaminopropane)、腺苷-5-但磷酸鹽(Adenosine-5-Monophosphate)、丁烷-2, 3-二醇(Butane-2, 3-Diol)、二羥丙酮磷酸( Dihydroxyacetone phosphate)、羥胺(Hydroxylamine)、草酸(Oxamic Acid)、核酮糖-5-磷酸鹽(Ribulose-5-Phosphate)、維生素C(Ascorbic Acid)、反丁烯二酸(Fumaric acid)、二十一烷酸(Heneicosanoic Acid)、木酮糖(D-Xylulose)、磷酸海藻糖(Trehalose-6-Phosphate)等代謝物顯著下調(diào)。上述差異代謝物變化表明,較高濃度碳基納米銅(50 mg·L-1)可對普通小球藻的糖類和小分子酸類代謝物產(chǎn)生明顯影響,導(dǎo)致這兩大類代謝物的下調(diào)。將所得到的差異代謝物進(jìn)行聚類分析,得到如圖4所示的熱圖。

圖4 空白對照組、5 mg·L-1、50 mg·L-1碳基納米銅體系普通小球藻代謝物熱圖Fig. 4 Heatmap of the C. vulgaris metabolites in control, 5 mg·L-1, and 50 mg·L-1 C-Cu2O nanoparticles groups
圖4中,紅色色塊表示對應(yīng)的代謝物相對豐度的增加,藍(lán)色色塊代表對應(yīng)代謝物的減少,顏色越深,代表增加或減少的程度越重。當(dāng)碳基納米銅的濃度從 5 mg·L-1上升至 50 mg·L-1時,大部分差異代謝物的相對含量降低,這些代謝物多為糖類、氨基酸類和小分子酸類。這表明,高濃度的碳基納米銅對普通小球藻產(chǎn)生了顯著的抑制作用,同等脅迫條件下,藍(lán)藻門的銅綠微囊藻對碳基納米銅的耐受能力遠(yuǎn)弱于普通小球藻,高濃度的碳基納米銅復(fù)合材料會導(dǎo)致銅綠微囊藻細(xì)胞內(nèi)部的淀粉粒數(shù)量明顯增加,大量的銅綠微囊藻細(xì)胞出現(xiàn)細(xì)胞膜破裂,與此同時,高濃度(50 mg·L-1)碳基納米銅的存在會使銅綠微囊藻的氨基酸循環(huán)遭到嚴(yán)重破壞(Faraz et al.,2019;Mavrikou et al.,2019)。
2.2.2 不同濃度碳基納米銅體系中藻細(xì)胞差異富集通路分析
在討論了不同碳基納米銅濃度對普通小球藻細(xì)胞代謝物種類數(shù)量變化、差異代謝物相對豐度的基礎(chǔ)之上,對差異代謝物進(jìn)行 MetPA(Metabolomics Pathway Analysis)分析。結(jié)果如圖5所示,橫坐標(biāo)為impact,縱坐標(biāo)為KEGG通路富集程度,圖中實心圓的大小代表該通路impact值。圖中圓點越大,表示通路impact值高,impact大于0.1為重要擾動通路;圖中顏色表示顯著性,顏色越深,表示P值越小。發(fā)現(xiàn)了與這些生物標(biāo)記物相關(guān)的9種代謝物顯著富集通路,分別為精氨酸合成通路(Arginine biosynthesis)、戊醣酸通路(Pentose phosphate pathway)、葡萄糖合成通路(Gluconeogenesis)、半乳糖代謝通路(Galactose metabolism)、抗壞血酸和醛酸代謝通路(Ascorbate and aldarate metabolism)、嘧啶代謝通路(Pyrimidine metabolism)、氨基糖和核苷酸糖代謝(Amino sugar and nucleotide sugar metabolism)。
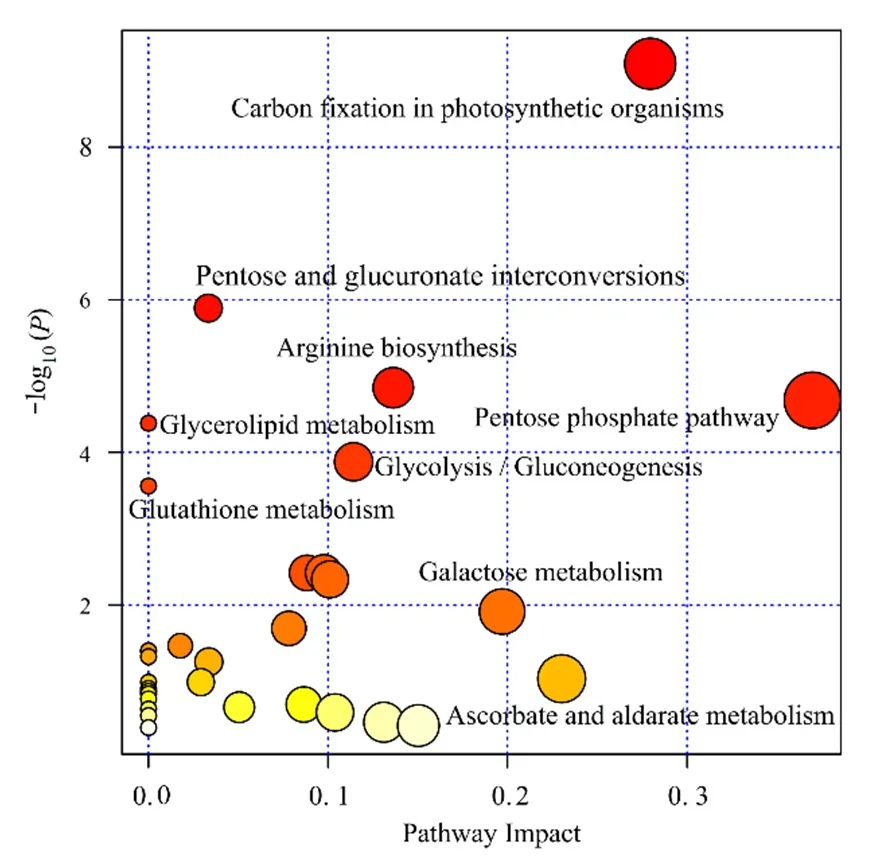
圖5 不同濃度碳基納米銅材料體系中藻細(xì)胞MetPA通路圖Fig. 5 MetPA pathway of C. vulgaris in different concentrations of C-Cu2O nanomaterials
如圖5所示,代謝通路分析表明,差異代謝物富集最多的是光合作用碳固定代謝通路(Carbon fixation in photosynthetic organisms),其次是氨基酸糖代謝通路(Amino sugar and nucleotide sugar metabolism)。高濃度碳基納米銅影響普通小球藻的機體代謝,抑制光合作用的碳固定過程,造成藻細(xì)胞體內(nèi)糖、氨基酸等物質(zhì)的穩(wěn)態(tài)變化(Chen et al.,2019;Gong et al.,2018)。研究表明,藻細(xì)胞內(nèi)氨基酸的合成與葉綠素a的形成密切相關(guān)(Lammel et al.,2019;Yarmohammadi et al.,2019),這與高濃度碳基納米銅導(dǎo)致普通小球藻葉綠素a含量明顯下降的現(xiàn)象吻合。當(dāng)藻細(xì)胞處于外界脅迫后,細(xì)胞會啟動自我防御機制,這種防御的常見表現(xiàn)之一是細(xì)胞內(nèi)淀粉粒的增加,而淀粉粒的增加與糖代謝通路的顯著富集有密不可分的關(guān)系(Kumar et al.,2015)。
圖6為不同濃度碳基納米銅復(fù)合材料對普通小球藻產(chǎn)生脅迫作用后,藻細(xì)胞內(nèi)部幾大類主要代謝通路和通路途徑中代謝物的變化。紅色標(biāo)記的為上調(diào)的代謝物,藍(lán)色標(biāo)記的為下調(diào)的代謝物。高濃度碳基納米銅的脅迫作用使藻細(xì)胞中糖代謝、氨基酸代謝、核酸代謝這三類代謝的特性發(fā)生了明顯的變化。高濃度的碳基納米銅會導(dǎo)致小球藻主要通路活性的降低(Lauritano et al.,2019)。圖6中,卡爾文循環(huán)(Calvin cycle)中的大部分代謝物降低,光合碳循環(huán)代謝被顯著削弱;同時,藻細(xì)胞內(nèi)的糖代謝也被明顯抑制,作為三羧酸循環(huán)(TCA cycle)上游產(chǎn)物的三磷酸甘油,其在卡爾文循環(huán)中含量的降低也導(dǎo)致了三羧酸循環(huán)中一部分代謝物的降低。這一變化進(jìn)而間接導(dǎo)致抗壞血酸和鳥氨酸等氨基酸的代謝降低,從而減弱整個氨基酸代謝通路(Hu et al.,2020)。

圖6 不同濃度碳基納米銅材料體系中藻細(xì)胞代謝通路變化Fig. 6 Metabolic pathway of C. vulgaris in different concentrations of C-Cu2O nanomaterials
3 結(jié)論
本文以普通小球藻為受試對象,設(shè)置了不同濃度的碳基納米銅暴露條件,在實驗周期內(nèi)進(jìn)行了藻細(xì)胞濃度變化、葉綠素a含量、代謝組學(xué)變化的測定,得出以下結(jié)論:
當(dāng)碳基納米銅質(zhì)量濃度范圍為 0.05—5 mg·L-1時,其對小球藻生長及葉綠素a含量并沒有明顯的抑制作用;當(dāng)碳基納米銅濃度達(dá)到50 mg·L-1時,其對普通小球藻產(chǎn)生了較為嚴(yán)重的脅迫作用,多數(shù)氨基酸的相對豐度降低,這表明高濃度的碳基納米銅復(fù)合材料使得藻細(xì)胞內(nèi)氨基酸代謝活力降低,這也導(dǎo)致藻細(xì)胞內(nèi)差異代謝物和代謝路徑的變化。高濃度的碳基納米銅復(fù)合材料導(dǎo)致藻細(xì)胞內(nèi)甘氨酸代謝被破壞,其相對豐度顯著降低,這直接導(dǎo)致了藻細(xì)胞光合作用的碳固定能力減弱;這些內(nèi)在的脅迫作用直接導(dǎo)致高濃度碳基納米銅復(fù)合材料脅迫條件下藻細(xì)胞內(nèi)葉綠素a含量的下降。
猜你喜歡
建材發(fā)展導(dǎo)向(2022年2期)2022-03-08 01:44:04
建材發(fā)展導(dǎo)向(2021年14期)2021-08-23 00:56:16
中國材料進(jìn)展(2019年10期)2019-12-07 05:32:14
纖維復(fù)合材料(2018年3期)2018-04-25 07:22:58
電子測試(2017年11期)2017-12-15 08:57:13
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:34
中國塑料(2015年6期)2015-11-13 03:02:54
中國塑料(2015年11期)2015-10-14 01:14:14
中國塑料(2015年8期)2015-10-14 01:10:41
應(yīng)用化工(2014年10期)2014-08-16 13:11:29
