脫氧雪腐鐮刀菌烯醇致動物厭食和嘔吐機制的探討
2021-04-13 02:40:56白永松馬愷笛孟慶維單安山
動物營養學報 2021年3期
關鍵詞:小鼠
白永松 馬愷笛 孟慶維 單安山
(東北農業大學動物營養研究所,哈爾濱150030)
霉菌毒素是由霉菌產生的有毒次級代謝產物,廣泛存在于飼料原料和人類食品中,嚴重威脅著動物健康和人類食品安全。霉菌毒素污染是世界上多個國家都面臨的重大問題,全球每年有很多食品或飼料受到不同程度的霉菌毒素污染[1]。在中國,霉菌毒素污染的形勢也尤為嚴峻。Yang等[2]發布的最新報道顯示,中國的玉米和玉米制品中霉菌毒素的檢出率較高,并且檢出率排在前3位的霉菌毒素分別是脫氧雪腐鐮刀菌烯醇(deoxynivalenol,DON)、黃曲霉毒素B1(aflatoxin B1,AFB1)和玉米赤霉烯酮(zearalenone,ZEN)。DON是眾多霉菌毒素中污染范圍最廣、檢出頻率最高的一種霉菌毒素,因此,DON污染是一個重大的全球性食品安全問題。
DON是由禾谷鐮刀菌(F.graminearum)和黃色鐮刀菌(F.culmorum)等產生的一種具有毒害作用的真菌毒素,其廣泛存在于小麥、大麥、燕麥、玉米等谷物中,嚴重威脅著食品和飼料安全。DON對人和動物有很強的毒性,急性中毒能引起嘔吐,而長期接觸低濃度DON可引起厭食、體重下降、腹瀉、腸黏膜損壞以及免疫系統損傷[3-4]。DON攝入能夠使動物產生氧化應激反應,進而導致DNA損傷和細胞凋亡[5]。DON還能夠改變神經內分泌信號傳導、誘導促炎基因表達、破壞生長激素軸和改變腸道的完整性[4]。此外,動物的食欲中樞也是DON致機體損傷的重要靶點,它可造成動物采食量下降,產生厭食或嘔吐反應。厭食和嘔吐反應是非常復雜的過程,它由神經中樞、胃腸激素以及腸道微生物等元素共同進行調控。目前在學術界對于DON引起動物厭食和嘔吐反應的機制已有一些報道,本文從食欲調控中樞、食欲調節激素、炎癥細胞因子以及腸道微生物這4個方面總結了DON誘導動物產生厭食和嘔吐反應的可能途徑,闡述了腸道微生物與DON誘導的動物厭食和嘔吐反應之間的潛在聯系,為更深層次研究DON的毒性機制奠定基礎,也為尋找DON解毒劑提供新的思路。
1 DON誘導動物產生厭食和嘔吐
DON具有神經毒性,能夠改變神經內分泌信號傳導,對腦部產生毒性作用,導致動物產生厭食反應[6]。豬采食受到低濃度DON(1~2 mg/kg)污染的飼糧可導致采食量和日增重下降,引起部分厭食,而當攝入飼糧的DON濃度達到12 mg/kg時就會引起完全厭食反應[7-8]。Meta分析顯示,飼糧中含有約3.63 mg/kg的DON可導致生長豬采食量和日增重均下降26%,并且豬的采食量隨飼糧中DON濃度(0.02~2.50 mg/kg)的增加而線性降低[9-10]。最新研究顯示,28日齡斷奶仔豬連續28 d攝入含有1.28或2.89 mg/kg DON的飼糧,可造成其采食量和日增重顯著降低[11]。此外,除了以豬為模型研究DON的厭食作用外,部分學者還利用大鼠和小鼠建立模型研究DON的毒性作用。大鼠在攝入0.25~1.00 mg/kg DON污染飼糧持續9周后,可明顯觀察到大鼠采食量降低以及體重下降[12]。給小鼠連續7~14 d飼喂0.35~3.00 mg/kg DON污染飼糧,可造成小鼠采食量和體重顯著降低[13-14]。另外,小鼠腹腔注射和灌胃模型均證明了DON的厭食作用。按體重給小鼠腹腔注射1.0或2.5 mg/kg DON,在注射0.5~3.0 h后小鼠出現厭食反應,并且呈劑量依賴性,但在6 h后小鼠采食量逐漸恢復,至16 h時觀察到了劑量依賴性促食欲反應,因此短期DON暴露對動物食物攝入的影響是短暫的,并且短期DON暴露有一定的促食欲作用[15-16],灌胃模型小鼠試驗也證實了這一理論,按體重給小鼠灌胃6.25、12.50或25.00 mg/kg的DON,在灌胃后的0~3 h內小鼠采食量顯著降低,而在6 h后小鼠采食量逐漸增加,在18~24 h時各試驗組小鼠采食量均高于對照組,出現了促食欲的作用;但從整個試驗周期的統計數據可知,DON有明顯的厭食作用[17]。
嘔吐反射是動物在攝取有毒食物后機體產生的自我保護機制。DON攝入對人和動物有著強烈的催吐作用,嘔吐是DON致人和動物損傷的主要癥狀之一[18]。慢性低濃度攝入DON,可使機體產生厭食反應、體重增加緩慢甚至下降以及生長激素失調;而急性高濃度攝入DON可引起機體腸胃炎癥,產生嘔吐反應甚至是休克[19]。不同動物對DON的敏感性不同,物種的差異導致DON吸收、分布以及代謝有著很大差異,動物對DON敏感性從大到小依次為豬、小鼠、大鼠、家禽、反芻動物[20]。早在1977年,Forsyth等[21]建立了DON致豬嘔吐的模型,研究結果顯示對于9~10 kg的仔豬,按每千克體重口服0.1~0.2 mg DON或者按照每千克體重腹腔注射0.05 mg DON即可引起仔豬的嘔吐反應。而對于15~20 kg的仔豬,按體重靜脈注射80 μg/kg DON或口服300 μg/kg DON則可觀測到仔豬發生嘔吐[22]。除了口服或注射一定劑量DON可使動物產生嘔吐反應外,動物采食受到DON污染的飼料也會導致動物發生嘔吐。仔豬食用含有約20 mg/kg DON的飼糧即可引起仔豬嘔吐,當攝入飼糧中DON濃度為12 mg/kg時會引起仔豬拒食,而飼糧中含有僅1.3 mg/kg DON就會引起仔豬采食量下降和體重增加速度變緩[23]。
2 DON對下丘腦食欲調控中樞的影響
動物的采食量或食欲受到外周神經和下丘腦中樞神經系統的調控。來自腸道、胰腺、肝臟或脂肪組織的外周信號直接或間接聚集于下丘腦,形成臟器、組織、腦干和下丘腦之間的通信網絡,下丘腦和腦干則整合外周激素和中樞神經遞質信號并通過高級皮質中樞調控食欲,從而影響腸胃運動、激素分泌以及機體的能量平衡[24]。下丘腦弓狀核(arcuate nucleus,ARC)在食欲調控中發揮著重要作用,其與下丘腦腹內側核(ventromedial nucleus,VMN)、背中核(dorsomedial nucleus,DMN)、室旁核(paraventricular nucleus,PVN)、外側下丘腦(lateral hypothalamic area,LHA)、視交叉上核(suprachiasmatic nucleus,SCN)等共同調控動物的食欲。ARC是中樞食欲調節神經網絡的核心,其包含2類主要的神經元,一類是促食欲神經元神經肽Y(neuropeptide,NPY)/刺鼠相關蛋白(agouti-related peptide,AGRP),另一類是抑食欲神經元阿片黑素皮質素原(pro-opiomelanocortin,POMC)/可卡因-安菲他明調節轉錄物(cocaine-amphetamine-regulated transcript,CART)[25]。
DON誘導的動物厭食反應與其對大腦的毒性有關,按體重給小鼠灌服25 mg/kg DON,在5 min后便可在大腦內檢測到(0.8±0.1) μg/g DON,這表示DON可迅速轉移至大腦并對其產生毒性作用,這也暴露了毒素干擾正常食欲調節的可能性[26]。促食欲神經元和抑食欲神經元共同調控動物食欲。在肥胖小鼠模型中,小鼠連續7周攝入含有10 mg/kg DON的飼糧后,可明顯觀察到下丘腦中AGRPmRNA表達水平上升,并且POMCmRNA表達水平也呈上升趨勢[27]。小鼠按體重急性灌胃12.5 mg/kg DON 3 h后,下丘腦POMC及其下游基因黑皮質素受體4(melanocortin 4 receptor,MC4R)、腦源性神經營養因子(brain-derived neurotrophic factor,BDNF)、酪氨酸蛋白激酶受體B(tyrosine kinase receptor B,TrkB)的mRNA表達水平顯著升高,同時促腎上腺皮質激素釋放激素(corticotrophin-releasing hormone,CRH)mRNA表達水平呈上升趨勢,但對下丘腦NPY和AGRP的mRNA表達水平沒有顯著影響,這表示DON調節厭食途徑而不影響攝食途徑[17]。然而,物種差異可能導致DON誘導的厭食反應機制也不一致,給仔豬飼喂含2.89 mg/kg DON的飼糧持續28 d,仔豬下丘腦POMC的蛋白表達水平顯著增加,而NPY的蛋白表達水平顯著下降,顯示DON既能夠對厭食途徑產生影響,也能夠對攝食途徑產生影響[28]。然而,以豬為模型研究DON對厭食和攝食中樞調控的報道較少,DON誘導的厭食機制在不同物種之間的差異有待進一步研究。
5-羥色胺(serotonin,5-HT),又稱血清素,其作為神經遞質參與調控動物的食欲。研究顯示,動物的攝食量和5-HT濃度之間呈負相關,并且5-HT還與抑食欲和促食欲的多肽有著緊密的關聯,是POMC、α-黑素細胞刺激素(α-melanocyte-stimulating hormone,α-MSH)、NPY以及AGRP的重要上游調節因子[29]。按體重給小鼠灌胃2.5 mg/kg DON,在24 h后小鼠小腦和下丘腦中的5-HT濃度顯著增加[30]。此外,按體重急性給藥0.25 mg/kg DON 8 h后便可觀察到豬下丘腦中的5-HT濃度顯著升高[31]。以水貂為模型研究DON誘導嘔吐反應機制的研究顯示,根據體重給水貂腹腔注射0.10或0.25 mg/kg DON后,在15~30 min內可觀察到嘔吐反應,并且在60 min后水貂血漿5-HT濃度顯著升高,用5-HT3受體拮抗劑進行預處理后可完全抑制DON誘導的嘔吐反應[32],并且DON能夠激活鈣敏感受體(calcium-sensing receptor,CaSR)和瞬時受體電位離子通道1(transient receptor potential ankyrin 1,TRPA1),從而誘導腸嗜鉻細胞(enterochromaffin cell,EC細胞)釋放5-HT(圖1)[33]。以豬為模型的研究顯示,豬攝入DON后血漿中5-HT及其代謝物5-羥基吲哚乙酸的濃度顯著增加[34],而5-HT3受體拮抗劑可抑制DON誘導的豬嘔吐反應[22],因此DON誘導的嘔吐作用與5-HT途徑有關,其機制可能是,5-HT3受體受到DON的刺激后引起細胞外鈣離子(Ca2+)通過5-HT3Rs/L型Ca2+通道進入細胞內,從而增加細胞內Ca2+濃度。細胞內Ca2+濃度的升高激活了鈣調素依賴性蛋白激酶Ⅱa(calmodulin dependent protein kinases Ⅱa,CaMKⅡa)和細胞外信號調節蛋白激酶1/2(extracellular regulated kinase 1/2,ERK1/2),促進動物產生嘔吐反應[35]。另外,5-HT調控食欲的相關受體主要有5-HT1BR、5-HT2CR和MC4R,黑素皮質素與MC4R結合后參與5-HT2CR對食欲的調控[36],5-HT與5-HT2CR結合能夠激活POMC,促進其表達,并產生α-MSH,從而抑制動物的食欲[37]。5-HT1BR能夠抑制促食欲神經元NPY/AGRP的活性,從而抑制動物的食欲[29,38]。因此,5-HT及其受體均參與DON誘導的動物食欲下降和嘔吐反應,進而導致動物拒食和體重降低。
3 DON對胃腸激素的影響
胃腸道在調節食欲中起著關鍵作用。動物的食欲和飽腹感不僅與腸道活動息息相關,還與位于胃腸道內的腸內分泌細胞(enteroendocrine cell,EEC)所分泌的胃腸激素有著密切的聯系[39]。EEC分泌的胃腸激素有30多種,其中與食欲調控相關的胃腸激素主要有酪酪肽(peptide YY,PYY)、膽囊收縮素(cholecystokinin,CCK)、胰高血糖素樣肽-1(glucagon-like peptide-1,GLP-1)、抑胃肽(gastric inhibitory peptide,GIP)等,這些胃腸激素均參與了DON誘導的厭食反應[40]。
PYY是調節食欲和能量的胃腸飽感激素,同時CCK也作為一種飽感激素調節動物攝食行為。PYY和CCK是DON誘導厭食反應和抑制動物生長的重要介質。按照每千克體重給小鼠腹腔注射或者口服1或5 mg DON 15 min后,血漿PYY和CCK濃度顯著增加,小鼠的采食量降低,引起小鼠產生厭食反應,并且NPY2受體拮抗劑能夠減弱DON誘導的厭食反應,而CCK受體拮抗劑的作用不明顯,這表明在DON誘導的厭食反應中PYY是一個比CCK更為關鍵的介質[41]。此外,成年小鼠(3個月)和老齡小鼠(22個月)對DON的敏感性不同,在急性DON暴露后成年小鼠和老年小鼠血漿PYY和CCK濃度均顯著增加,但老齡小鼠血漿胃腸飽感激素濃度更高,因此年齡是DON誘導胃腸激素改變程度大小的一個相關因素[42]。根據體重給小鼠灌胃2.5 mg/kg DON后的0.5 h便可觀察到血漿CCK濃度的顯著增加[43],因而DON可能通過刺激EEC增加CCK的濃度而誘導動物食欲降低。研究顯示,DON誘導CCK產生的機制可能是通過激活CaSR和TRPA1介導的[44],DON暴露可顯著提高胃腸組織CCK和TRPA1的mRNA表達水平[17],并且CCK濃度的增加可通過迷走神經激活下丘腦中的POMC神經元,進而降低動物食欲[45]。除CCK外,PYY的產生機制也與CaSR和TRPA1有著一定的關聯(圖1)[46],并且PYY還能夠通過迷走神經參與調控下丘腦中抑食欲神經元POMC和促食欲神經元NPY的表達,促進POMC的上調和NPY的下調[47],這說明胃腸激素和下丘腦共同調控動物的食欲,但具體機制仍然未知,有待進一步研究。
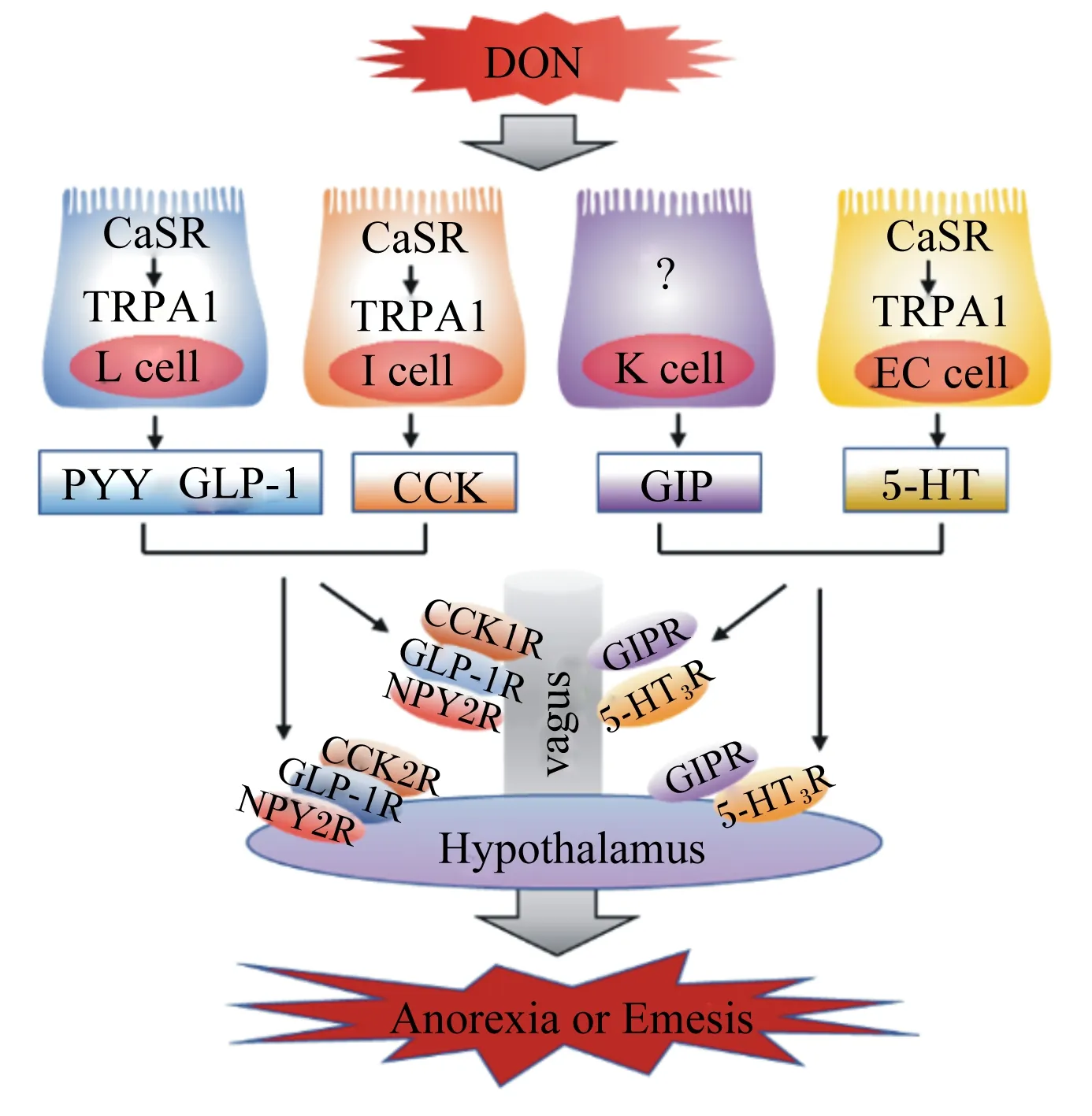
DON:脫氧雪腐鐮刀菌烯醇 deoxynivalenol;CaSR:鈣敏感受體 calcium-sensing receptor;TRPA1:瞬時受體電位離子通道1 transient receptor potential ankyrin 1;L cell:L細胞;I cell:I細胞;K cell:K細胞;EC cell:腸嗜鉻細胞 enterochromaffin cell;PYY:酪酪肽 peptide YY;GLP-1:胰高血糖素樣肽-1 glucagon-like peptide-1;CCK:膽囊收縮素 cholecystokinin;GIP:抑胃肽 gastric inhibitory peptide;5-HT:5-羥色胺 serotonin;CCK1R:膽囊收縮素1受體 cholecystokinin 1 receptor;CCK2R:膽囊收縮素2受體 cholecystokinin 2 receptor;NPY2R:神經肽Y2受體 neuropeptide Y2 receptor;GIPR:抑胃肽受體gastric inhibitory peptide receptor;5-HT3R:5-羥色胺3受體 serotonin 3 receptor;GLP-1R:胰高血糖素樣肽-1受體 glucagon-like peptide-1 receptor;vagus:迷走神經;Hypothalamus:下丘腦;Anorexia or Emesis:厭食或嘔吐。
由于嚙齒動物缺乏嘔吐反射,因此很多學者以豬和水貂為研究模型探討DON對動物產生的厭食和嘔吐反應。斷奶仔豬連續28 d攝入含1~3 mg/kg DON的飼糧,其血漿PYY濃度顯著增加[28]。仔豬血漿PYY和CCK濃度與DON的毒性作用有關,攝入不同濃度的DON污染飼糧,均可觀察到血漿PYY和CCK濃度增加,并且與DON攝入的時間和濃度呈線性關系,然而由于DON濃度較低,并未使仔豬產生嘔吐反應[48]。另外,按體重給水貂口服0.25 mg/kg DON后,僅15 min便可觀察到其開始產生嘔吐反應,并且其血漿PYY濃度顯著增加,在使用NPY2受體拮抗劑后可降低血漿PYY的濃度,并且產生嘔吐反應的概率下降50%[32]。因此,DON誘導的食欲下降和嘔吐反應與動物血漿PYY和CCK的濃度有著緊密的聯系,DON可促進PYY和CCK的分泌,進而導致動物食欲下降,采食量降低,甚至抑制動物生長。
GLP-1和GIP分別來源于腸內分泌L細胞和K細胞,二者是與能量平衡相關的飽感激素,在食欲調控中起著非常重要的作用。按每千克體重給小鼠灌胃1.0或2.5 mg DON,小鼠血漿GLP-1和GIP的濃度在30 min后顯著增加,但在6 h后各組之間血漿GLP-1和GIP的濃度差異不顯著,此結果表示DON誘導血漿GLP-1和GIP濃度升高是短暫可逆的,并且用GLP-1受體拮抗劑Exendin9-39和GIP受體拮抗劑Pro3GIP進行預處理可降低DON誘導的食欲下降[16]。研究顯示,DON通過激活CaSR以及瞬時受體電位離子通道-5(transient receptor potential ion channel-5,TRPM-5)和TRPA1,促進GLP-1的分泌,進而導致動物食欲降低(圖1)[16,44]。此外,斷奶仔豬攝入受到DON污染的飼糧后,其血漿GLP-1濃度顯著升高[28]。因此,GLP-1和GIP在DON誘導的食欲下降和厭食反應中起著重要作用,但DON誘導腸內分泌細胞分泌GLP-1和GPI的具體機制尚不清楚,未來的研究中應更深入探討DON誘導厭食反應的機制。
4 DON誘導的厭食反應與炎癥細胞因子有關
DON引起厭食反應的一個潛在原因可能是炎癥細胞因子的誘導。細胞因子作為一種多肽分子,能夠廣泛調節細胞功能,作用于免疫系統、造血系統以及神經、內分泌系統,對動物的生長發育具有重要作用。細胞因子基因的異常表達可引起許多發育、生理和免疫問題,例如白細胞介素-1β(IL-1β)、白細胞介素-6(IL-6)和腫瘤壞死因子-α(TNF-α)等細胞因子的異常表達可導致人類和動物產生疾病[49]。按體重給小鼠灌胃25 μg/kg DON持續10 d便可引起低程度的炎癥狀態,其血漿、脂肪組織、十二指腸和大腦中IL-1β表達均顯著增加[50]。另外,DON暴露可增加小鼠血漿IFN-α、白細胞介素-2(IL-2)、白細胞介素-4(IL-4)和IL-6的濃度,同時DON也不同程度地影響小鼠血漿IL-1β、白細胞介素-10(IL-10)和TNF-α的濃度[51]。已經有研究證實,促炎性細胞因子的分泌可能會降低動物的采食量,血漿IL-1β和IL-6的濃度與DON引起的厭食反應有關[42]。給小鼠灌服DON可誘導厭食反應,并且急性外源腹腔注射TNF-α和IL-1β同樣也可引起小鼠食欲下降和厭食,而使用腫瘤壞死因子-α受體拮抗劑(TNFR)和白細胞介素1受體拮抗劑(IL-1RA)對小鼠進行預處理后可緩解DON、TNF-α和IL-1β所引起的厭食反應,因此,TNF-α和IL-1β在DON誘導的厭食反應中發揮著重要作用[52]。小鼠在按照每千克體重口服12.5 mg DON 3 h后下丘腦的炎癥細胞因子和POMC的表達水平均顯著增加[17],最終誘導小鼠產生厭食反應,而炎癥細胞因子誘導厭食反應的中間媒介可能是核轉錄因子-κB(nuclear factor-kappa B,NF-κB),被證明可以調節炎癥相關基因,包括IL-1β、TNF-α和IL-6,并且NF-κB能夠激活POMC神經元,進而調節食欲調控中樞,產生厭食反應[53]。
5 腸道微生物在DON誘導厭食反應中的作用
腸道中棲居著大量的微生物群落,這些微生物與宿主之間有著緊密的聯系,建立了一個穩定共生的動態體系,維持宿主腸道微環境的穩態。腸道微生物可利用宿主腸道的營養物質進行發酵產生發酵產物,進而調控營養物質的吸收、代謝以及機體免疫系統。腸道微生物菌群失調可導致宿主產生相關應激行為,例如抑郁、焦慮以及食欲減退等[54],并且在一些精神類疾病和代謝性疾病中都可觀察到腸道微生物菌群的改變[55]。DON可破壞腸道組織完整性,降低腸道絨毛高度,抑制腸道細胞分化,破壞腸道微生物平衡[4,56]。研究顯示,斷奶仔豬攝入1或3 mg/kg DON飼糧持續28 d,其腸道微生物結構發生了巨大改變,DON顯著降低小腸中厚壁菌門和變形菌門的豐度,增加放線菌門和藍藻菌門的豐度[57]。另外,給4周齡小鼠按體重隔天灌胃5 mg/kg的DON,持續14 d,小鼠體重和日增重顯著降低,并且DON暴露改變了腸道微生物菌群的組成,在門水平上擬桿菌門、壁厚菌門及脫鐵桿菌門的物種豐度顯著增加,在屬水平上擬桿菌屬、Mucispirillum和副桿菌屬(Parabacteroides)的物種豐度顯著增加,在種水平上的物種豐度也發生了改變,這些腸道微生物的改變影響了機體正常的生物合成和降解,并進一步導致代謝途徑紊亂[58]。低濃度的DON也可引起腸道微生物的變化,小鼠按體重每日灌胃25 μg/kg的DON持續30 d,可引起腸道微生物菌群的顯著改變,在門水平上DON顯著增加了變形菌門和疣微菌門的物種豐度,在屬水平上DON顯著增加了Parabacteroides和腸桿菌屬的物種豐度,而乳桿菌屬、Odoribacter、未定毛螺旋菌屬(Lachnospiraceaincertaesedis)的物種豐度顯著下降[59]。因此,無論是高濃度還是低濃度的DON均可導致動物腸道微生物群落的改變,這種改變可能是導致動物厭食的一個主要原因。
近年來,新一代測序技術進一步揭示了腸道微生物和宿主的食欲之間的密切聯系。腸道微生物可對宿主的中樞神經系統產生影響,從而間接調控宿主的食欲[60]。腸道微生物利用宿主的營養物質進行發酵產生代謝產物,如短鏈脂肪酸(short chain fatty acids,SCFAs),能通過游離脂肪酸受體2(FFAR2)和游離脂肪酸受體3(FFAR3)刺激腸道內分泌細胞釋放PYY和GLP-1[61],而PYY和GLP-1均已被證實在DON誘導的厭食反應中起著重要作用[28]。腸道微生物還可通過影響免疫系統,間接調控宿主食欲。腸道微生物與機體形成的宿主-微生物代謝軸,調控動物機體營養素代謝和免疫應答,從而影響食欲中樞,調控動物的攝食和厭食反應[56,62]。另外,腸道微生物可能通過調控5-HT的水平進而調控機體食欲,色氨酸是5-HT的前體物質,研究顯示腸道微生物能夠從頭合成色氨酸,進而參與5-HT的合成,調控動物食欲[63]。腸道微生物還可利用色氨酸進行代謝產生吲哚,吲哚能夠刺激腸道內分泌L細胞,調節GLP-1的分泌,進而影響動物食欲[64]。腸道微生物調控動物食欲的信號主要通過其代謝產生的SCFAs、5-HT以及吲哚等代謝產物來進行傳遞,而DON對5-HT以及由吲哚調節分泌的GLP-1有著直接的影響,因此腸道微生物與DON誘導的厭食和嘔吐反應有著一定的潛在聯系,其深入機制還有待進一步挖掘。
6 小 結
DON作為較常見的一種霉菌毒素嚴重威脅著人類的食品安全和畜牧業發展,其主要的毒性特點為誘導動物產生厭食和嘔吐反應。目前,對于DON致厭食和嘔吐的機制已有大量研究報道,但大部分僅停留在對神經元基因表達和胃腸激素含量的影響層面,缺乏深入細致的研究。另外,DON致厭食和嘔吐的可能途徑有多個(圖2),如胃腸激素途徑、5-HT途徑以及免疫系統途徑,這些途徑均與腸道微生物有著不同程度的聯系,因此腸道微生物與DON誘導的厭食和嘔吐反應有著潛在的聯系,但具體的調控機制未見報道,仍需更多的研究來闡明二者之間的聯系,為探究DON的毒性機制奠定理論基礎,為尋找更有效的DON抑制劑或解毒劑提供新思路,從而降低霉菌毒素帶來的危害,促進畜牧業的發展。
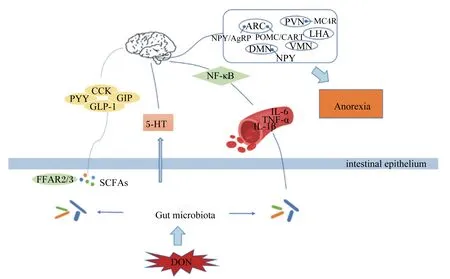
ARC:弓狀核 arcuate nucleus;PVN:室旁核 paraventricular nucleus;LHA:側下丘腦 lateral hypothalamic area;VMN:腹內側核 ventromedial nucleus;DMN:背中核 dorsomedial nucleus;MC4R:黑皮質素受體4 melanocortin 4 receptor;NPY:神經肽Y neuropeptide;AGRP:刺鼠相關蛋白 agouti-related peptide;POMC:阿片黑素皮質素原 pro-opiomelanocortin;CART:可卡因-安菲他明調節轉錄物 cocaine-amphetamine-regulated transcript;NF-κB:核轉錄因子-κB nuclear factor-kappa B;CCK:膽囊收縮素 cholecystokinin;PYY:酪酪肽 peptide YY;GIP:抑胃肽 gastric inhibitory peptide;GLP-1:胰高血糖素樣肽-1 glucagon-like peptide-1;5-HT:5-羥色胺 serotonin;IL-6:白細胞介素-6 interleukin-6;TNF-α:腫瘤壞死因子-α tumor necrosis factor-α;IL-1β:白細胞介素-1β interleukin-1β;Anorexia:厭食;FFAR2/3:游離脂肪酸受體2/3 free fatty acid receptor 2/3;SCFAs:短鏈脂肪酸 short chain fatty acids;intestinal epithelium:腸上皮;Gut microbiota:腸道微生物;DON:脫氧雪腐鐮刀菌烯醇 deoxynivalenol。
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國男科學雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
