家蠅Actin-5C基因啟動(dòng)子的生物信息學(xué)分析及載體構(gòu)建*
2021-04-13 07:10:06王藍(lán)晨王昱王穎楊仕賽朱貴明
貴州醫(yī)科大學(xué)學(xué)報(bào) 2021年3期
王藍(lán)晨,王昱,王穎,楊仕賽,朱貴明
(貴州醫(yī)科大學(xué) 生物與工程學(xué)院 實(shí)驗(yàn)中心,貴州 貴陽(yáng) 550025)
家蠅(MuscadomesticaL.)屬于完全變態(tài)昆蟲(chóng),是世界大部分地區(qū)最常見(jiàn)的一種昆蟲(chóng)[1]。家蠅具有生育能力強(qiáng)、生命周期短、易于高密度培養(yǎng)等特點(diǎn),且育種技術(shù)簡(jiǎn)單,飼料來(lái)源豐富,已被列為世界上新的蛋白質(zhì)資源昆蟲(chóng)[2-4]。因此將家蠅作為一種新型的轉(zhuǎn)基因生物反應(yīng)器進(jìn)行研究,具有廣闊的發(fā)展前景。昆蟲(chóng)轉(zhuǎn)基因技術(shù)是利用功能基因開(kāi)發(fā)新昆蟲(chóng)品種的關(guān)鍵,也是實(shí)現(xiàn)昆蟲(chóng)有用蛋白質(zhì)合成和生物反應(yīng)器開(kāi)發(fā)的關(guān)鍵技術(shù)[5-6]。外源基因能否成功表達(dá),啟動(dòng)子的選擇顯得尤其重要[7]。目前,對(duì)家蠅的轉(zhuǎn)基因研究相對(duì)不成熟,技術(shù)體系的建立尚不完善,仍缺乏能夠?qū)崿F(xiàn)外源基因高效表達(dá)的理想啟動(dòng)子。在果蠅和家蠶中,肌動(dòng)蛋白(Actin)是一種具有收縮特性的高度保守的組成型蛋白[8],肌動(dòng)蛋白除存在于肌肉細(xì)胞中,還是細(xì)胞骨架的重要組成部分、參與細(xì)胞遷移、染色體分離及細(xì)胞內(nèi)大分子轉(zhuǎn)運(yùn)等[9]。肌動(dòng)蛋白5C(Actin-5C)屬細(xì)胞質(zhì)肌動(dòng)蛋白,通常認(rèn)為將它祝為表達(dá)外源目的基因的啟動(dòng)子比較合適[10-11]。本研究使用多種預(yù)測(cè)工具預(yù)測(cè)Actin-5C啟動(dòng)子區(qū),并克隆預(yù)測(cè)調(diào)控區(qū)域,重組到含螢火蟲(chóng)熒光素酶的報(bào)告基因pGL3-Basic載體上,為進(jìn)行下一步的實(shí)驗(yàn)研究篩選出啟動(dòng)子活性最強(qiáng)的家蠅Actin-5C啟動(dòng)子的調(diào)控區(qū)域。
1 材料與方法
1.1 材料
1.1.1家蠅及細(xì)胞來(lái)源 供試蟲(chóng)源家蠅由貴州醫(yī)科大學(xué)生物與工程學(xué)院昆蟲(chóng)房飼養(yǎng),溫度25~28 ℃,濕度70%,光照周期12 L ∶12 D。供試細(xì)胞sf9(草地貪夜蛾)為貴州醫(yī)科大學(xué)生物與工程學(xué)院實(shí)驗(yàn)室保存。家蠅幼蟲(chóng)經(jīng)麥麩飼養(yǎng),家蠅成蟲(chóng)以白糖和奶粉以1 ∶1比例混合進(jìn)行飼養(yǎng),飲用水為無(wú)菌蒸餾水。
1.1.2主要試劑 熒火蟲(chóng)熒光素酶報(bào)告基因載體pGL3-Basic和pGL3-Promoter購(gòu)于美國(guó)Promega公司,Q5?超保真2× 預(yù)混液、限制性內(nèi)切酶及T4 DNA連接酶購(gòu)自美國(guó)NEB公司,DNA Marker、純化回收試劑盒及2×TaqPCR MasterMix Ⅱ購(gòu)自天根生化科技(北京)有限公司,氯化鈉、酵母粉及胰蛋白胨購(gòu)自英國(guó)Oxoid公司,瓊脂糖購(gòu)自中國(guó)biosharp公司,DNA抽提試劑盒購(gòu)自德國(guó)QIAGEN。胎牛血清(fetal bovine serum,FBS)購(gòu)自德國(guó)PAA公司,Grace′s昆蟲(chóng)培養(yǎng)基和細(xì)胞轉(zhuǎn)染試劑購(gòu)自美國(guó)Thermo ScientificTM公司。Primer 5.0設(shè)計(jì)引物(表1)、引物合成及基因測(cè)序均由生工生物工程(上海)股份有限公司完成。

表1 PCR實(shí)驗(yàn)中使用的引物Tab.1 Primers used in PCR experiments
1.1.3主要儀器 Applied Biosystems PCR儀購(gòu)自美國(guó)Thermo ScientificTM公司,DYY-BC型電泳儀購(gòu)自北京市六一儀器廠,Milli-Q超純水儀購(gòu)自德國(guó)Millipore Pharmacia,超凈工作臺(tái)購(gòu)自蘇州安泰空氣技術(shù)公司。
1.2 方法
1.2.1啟動(dòng)子分析 在美國(guó)國(guó)家生物信息中心(national center for biotechnology information,NCBI)數(shù)據(jù)庫(kù)中檢索家蠅肌動(dòng)蛋白Actin-5C基因gDNA序列,使用生物信息學(xué)預(yù)測(cè)工具分析得到啟動(dòng)子區(qū),其中啟動(dòng)子轉(zhuǎn)錄起始位點(diǎn)預(yù)測(cè)使用的是Promoter 2.0(http://www.cbs.dtu.dk/services/Promoter/),啟動(dòng)子預(yù)測(cè)分析使用伯克利果蠅基因組計(jì)劃(berkeley drosophila genome project,BDGP)在線分析軟件,以及Softberry 啟動(dòng)子預(yù)測(cè)分析工具包(包括TSSG、TSSP和FPROM),轉(zhuǎn)錄元件的預(yù)測(cè)使用的是Cister(https://zlab.bu.edu/~mfrith/cister.shtml)。
1.2.2熒光素酶載體構(gòu)建 Primer 5.0軟件進(jìn)行引物設(shè)計(jì),DNASTAR軟件行酶譜分析,在上游和下游引物5′端分別引入XhoⅠ和KpnⅠ限制性內(nèi)切酶酶切位點(diǎn)和保護(hù)堿基(表1)。提取家蠅成蟲(chóng)DNA并以此為模板,采用超保真聚合酶擴(kuò)增相應(yīng)的目的片段。PCR條件為98 ℃變性10 s,64 ℃退火30 s,72 ℃ 延伸50 s,共35個(gè)循環(huán),最后4 ℃保存。擴(kuò)增產(chǎn)物經(jīng)1%的瓊脂糖凝膠電泳,回收PCR產(chǎn)物。XhoⅠ和KpnⅠ內(nèi)切酶雙酶切PCR產(chǎn)物及載體pGL3-Basic純化回收后進(jìn)行連接反應(yīng),得到的質(zhì)粒分別命名為pGL3-1156、pGL3-1745及pGL3-2059三種質(zhì)粒,重組質(zhì)粒經(jīng)XhoⅠ和KpnⅠ雙酶切及測(cè)序進(jìn)行鑒。
1.2.3瞬時(shí)轉(zhuǎn)染細(xì)胞及熒光素酶基因的表達(dá) 取sf9細(xì)胞以1×105個(gè)/孔密度接種于6孔板中,細(xì)胞90%融合時(shí),加無(wú)血清無(wú)雙抗Grace′s培養(yǎng)基100 μL/孔,再加空載體pGL3-Basic和pGL3-Promoter各4 μg作為對(duì)照組,pGL3-1156、pGL3-1745及pGL3-2059各4 μg為實(shí)驗(yàn)組,轉(zhuǎn)染時(shí)加內(nèi)參載體phRL-TK400 ng和Lipofectamine2000 10 μL,補(bǔ)充無(wú)血清無(wú)雙抗的培養(yǎng)基250 μL,置28 ℃ 孵箱培養(yǎng)。6 h后棄培養(yǎng)液,換完全培養(yǎng)基。24 h后提取細(xì)胞總RNA,反轉(zhuǎn)錄聚合酶鏈反應(yīng)(reverse transcription-polymerase chain reaction,RT-PCR)檢測(cè)螢火蟲(chóng)熒光素酶基因的表達(dá)情況,上游引物Fluc-F為5′ TGCCTCATAGAACTGCCTGCGTG 3′,下游引物Fluc-R為5′ TATCATCCCCCTCGGGTGTAATCAG 3′。PCR擴(kuò)增時(shí)64 ℃退火,延伸40 s,循環(huán)30次,產(chǎn)物長(zhǎng)度為420 bp;并以βactin作為內(nèi)參照,上游引物βactin-F為5′ GATGACCCAGATCATGTTTGAGACC 3′,下游引物βactin-R為5′ TGTCACGCACGATTTCCCTCTC 3′,PCR擴(kuò)增時(shí)62 ℃退火,延伸10 s,循環(huán)30次,產(chǎn)物大小約為250 bp。結(jié)束后取反應(yīng)液5 μL進(jìn)行濃度為1%的瓊脂糖凝膠電泳,凝膠成像系統(tǒng)檢測(cè)PCR產(chǎn)物。
2 結(jié)果
2.1 家蠅Actin-5C基因定位及啟動(dòng)子分析
在線軟件NCBI對(duì)Actin-5C(NW_004766441)基因結(jié)構(gòu)進(jìn)行分析,結(jié)果顯示Actin-5C基因在GenBank數(shù)據(jù)庫(kù)中位于Scaffold 20349上,基因全長(zhǎng)為5 359 bp。使用BDGP在線分析Actin-5C基因啟動(dòng)子,以Actin-5C基因轉(zhuǎn)錄起始密碼子(ATG)的第一個(gè)堿基A作為+1,Actin-5C基因5′端上游-2 609~-94 bp區(qū)域內(nèi)有啟動(dòng)子活性,且啟動(dòng)子分值達(dá)0.8以上,其中-2 198~-2 148 bp時(shí)的分值最高,達(dá)到0.98(表2)。使用軟件TSSG、TSSP、Promoter 2.0和FPROM預(yù)測(cè)Actin-5C啟動(dòng)子基因序列,將預(yù)測(cè)結(jié)果匯總后制圖(圖1)。使用Cister預(yù)測(cè)轉(zhuǎn)錄元件,在-220 bp處檢測(cè)到CAAT盒信號(hào);在-1 444 bp處檢測(cè)到TATA盒信號(hào)(表3)。經(jīng)比較取最有可能的1 156 bp(-1 236~-80 bp)、1 745 bp(-1 825~-80 bp)和2 059 bp(-2 139~-80 bp)截?cái)鄦?dòng)子片段構(gòu)建pGL3-1 156、pGL3-1 745及pGL3-2 059三種質(zhì)粒。

表2 BDGP啟動(dòng)子預(yù)測(cè)工具預(yù)測(cè)Actin-5C基因非編碼區(qū)序列中的啟動(dòng)子Tab.2 BDGP promoter prediction tool for Actin-5C promoter in non-coding region sequence
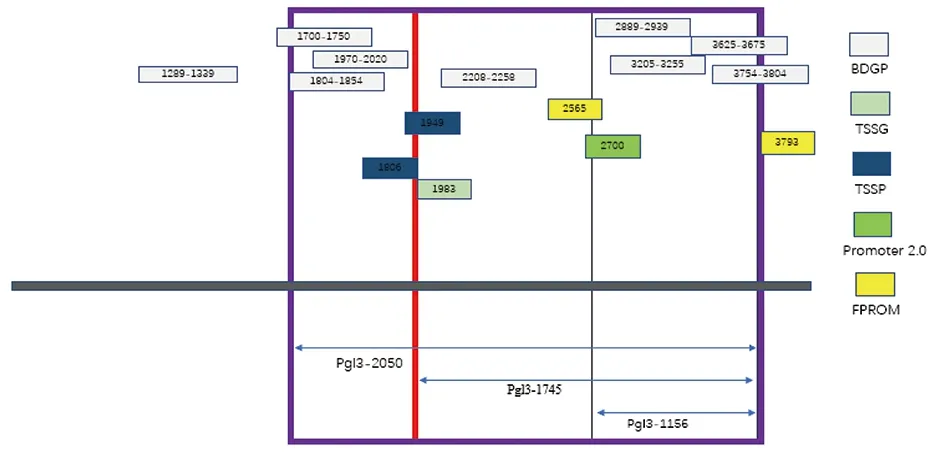
圖1 多種啟動(dòng)子預(yù)測(cè)軟件預(yù)測(cè)Actin-5C基因啟動(dòng)子Fig.1 A variety of promoter prediction software to predict the Actin-5C gene promoter
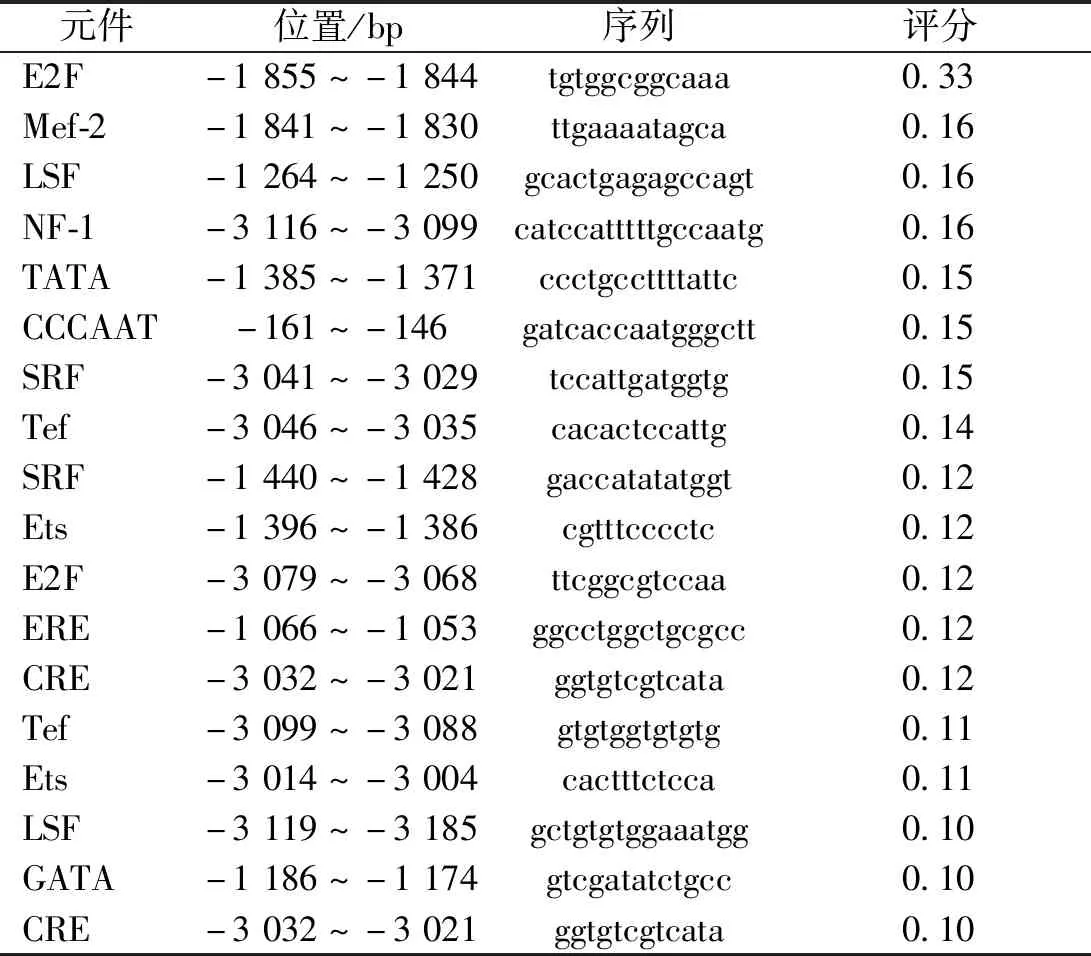
表3 Cister預(yù)測(cè)轉(zhuǎn)錄元件列表Tab.3 Cister predictive transcriptome list
2.2 家蠅Actin-5C轉(zhuǎn)基因載體的構(gòu)建
提取家蠅基因組DNA,并根據(jù)已經(jīng)報(bào)道的Actin-5C基因預(yù)測(cè)啟動(dòng)子區(qū),以家蠅基因組DNA為模板進(jìn)行PCR反應(yīng),PCR產(chǎn)物與載體pGL3-Basic經(jīng)同樣的酶切回收,回收片段大小分別為1 156、1 745及2 059 bp,片段大小正確;將回收后的目的片段連接到報(bào)告基因pGL3-Basic載體上,轉(zhuǎn)化篩選單克隆并提取陽(yáng)性重組質(zhì)粒,得到的重組質(zhì)粒pGL3-1 156,pGL3-1 745,pGL3-2 059,片段大小正確;經(jīng)Blast比對(duì)分析結(jié)果顯示堿基序列正確,表明重組質(zhì)粒構(gòu)建成功。見(jiàn)圖2和圖3。
圖3 重組質(zhì)粒的測(cè)序結(jié)果(節(jié)選)Fig.3 Sequencing results of recombinant plasmids(Excerpt)
2.3 熒光素酶編碼基因的轉(zhuǎn)錄檢測(cè)
將3個(gè)重組質(zhì)粒轉(zhuǎn)染sf9細(xì)胞后RT-PCR逆轉(zhuǎn)cDNA,以cDNA為模版檢測(cè)熒光素酶編碼基因,轉(zhuǎn)錄結(jié)果表示3個(gè)重組質(zhì)粒載體轉(zhuǎn)錄成功(圖4)。
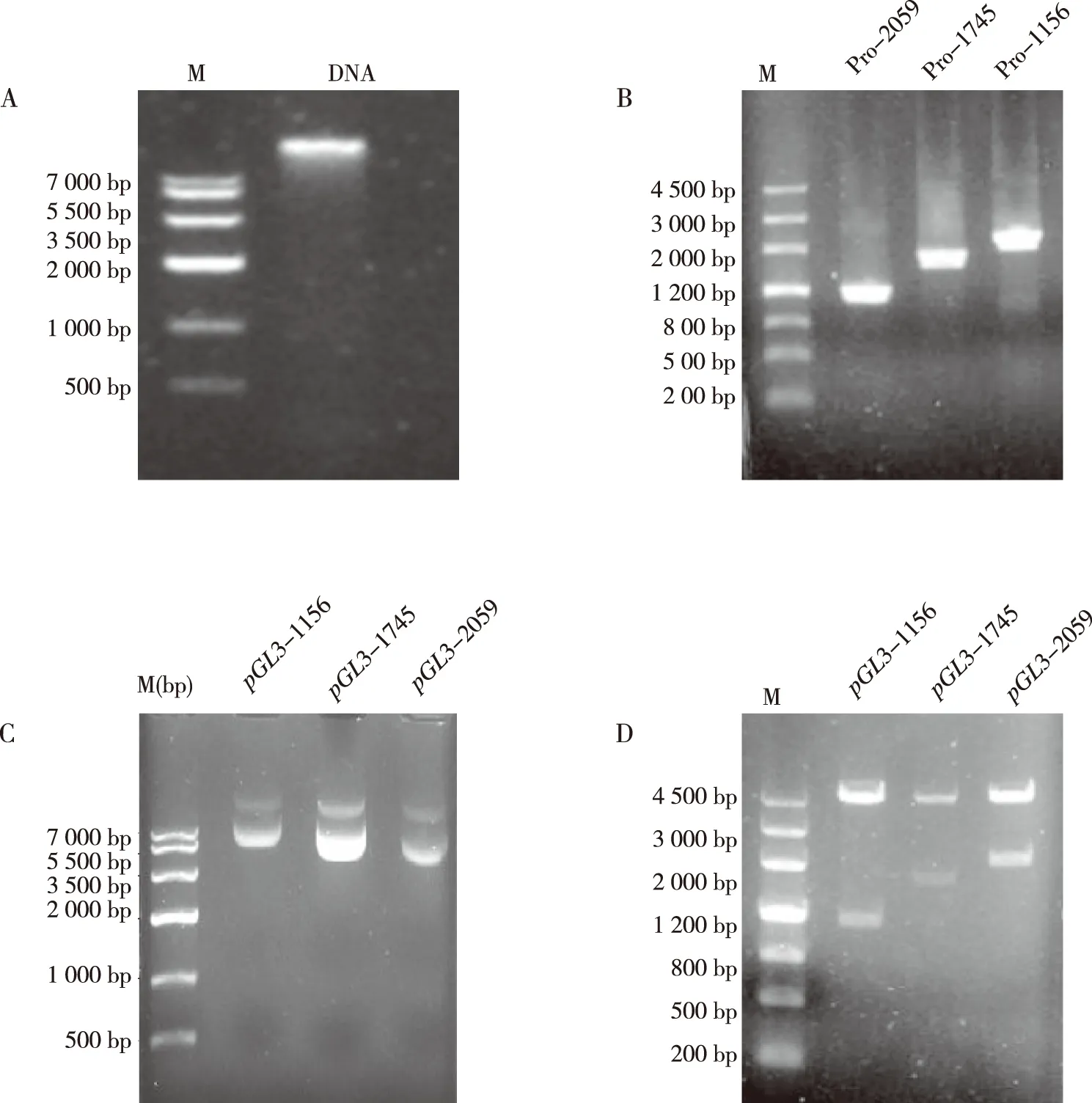
注:A為家蠅基因組DNA,B為酶切PCR產(chǎn)物,C為重組質(zhì)粒,D為雙酶切鑒定;M為DNA分子量標(biāo)準(zhǔn)Marker。圖2 啟動(dòng)子重組質(zhì)粒構(gòu)建Fig.2 Construction of recombinant plasmid using promoter
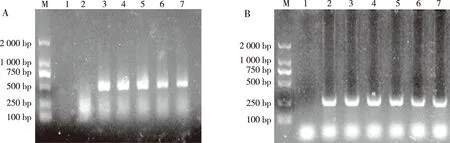
注:A為RT-PCR檢測(cè)結(jié)果,B為內(nèi)參;M為DNA分子量標(biāo)準(zhǔn)Marker,1為陰性對(duì)照,2為未轉(zhuǎn)sf9,3為pGL3-Basic,4為重組質(zhì)粒pGL3-1156,5為重組質(zhì)粒pGL3-1745,6為重組質(zhì)粒pGL3-2059,7為陽(yáng)性質(zhì)粒pGL3-Promoter。圖4 RT-PCR檢測(cè)重組質(zhì)粒Fig.4 RT-PCR detection of recombinant plasmids
3 討論
具有轉(zhuǎn)錄起始特異性的啟動(dòng)子是位于結(jié)構(gòu)基因5′末端上游的非編碼DNA序列,可以活化RNA聚合酶并使RNA聚合酶精確結(jié)合DNA模板,是基因轉(zhuǎn)錄調(diào)節(jié)的核心區(qū)域[12-13];真核生物啟動(dòng)子在轉(zhuǎn)錄調(diào)節(jié)中有著非常重要的作用,是RNA聚合酶特異性結(jié)合位點(diǎn)的轉(zhuǎn)錄調(diào)控元件[14-15]。利用生物信息學(xué)預(yù)測(cè)啟動(dòng)子,對(duì)于基因轉(zhuǎn)錄調(diào)控機(jī)制的研究這是一種重要的方法,它可以有效地從基因組中找到已知和未知的轉(zhuǎn)錄調(diào)控元件,為基因結(jié)構(gòu)和功能的研究提供可靠的依據(jù)[16-17]。在真核基因表達(dá)調(diào)控系統(tǒng)中,基因轉(zhuǎn)錄的起始和調(diào)控主要通過(guò)順式作用元件、反式作用因子和RNA聚合酶的協(xié)同作用實(shí)現(xiàn)[18]。順式作用元件在起始位點(diǎn)和基因轉(zhuǎn)錄的轉(zhuǎn)錄效率中起決定性作用[19],并且基因的轉(zhuǎn)錄起始通常通過(guò)啟動(dòng)子實(shí)現(xiàn)[20]。啟動(dòng)子序列由核心元件TATA盒、正向核心啟動(dòng)子元件BRE、反向啟動(dòng)子元件DPE、啟動(dòng)子、CAAT盒和GC盒組成,這些都是通過(guò)生物信息學(xué)研究啟動(dòng)子的重要信息[21-23]。本研究在NCBI數(shù)據(jù)庫(kù)對(duì)Actin-5C基因結(jié)構(gòu)定位分析,對(duì)Actin-5C基因5′端正向3 898 bp序列進(jìn)行分析,發(fā)現(xiàn)該基因的啟動(dòng)子區(qū)和核心啟動(dòng)子區(qū);利用Promoter 2.0軟件預(yù)測(cè)該基因的轉(zhuǎn)錄起始位點(diǎn);通過(guò)Cister軟件預(yù)測(cè),對(duì)非編碼區(qū)序列進(jìn)行分析,發(fā)現(xiàn)Actin-5C基因有TATA盒信號(hào),也有CAAT盒信號(hào),經(jīng)預(yù)測(cè)評(píng)分初步確定Actin-5C啟動(dòng)子的核心區(qū)域。
研究啟動(dòng)子需要生物信息學(xué)預(yù)測(cè)工具或軟件,它可以大大簡(jiǎn)化理解基因的過(guò)程,通過(guò)預(yù)測(cè)和基因比對(duì),人們可以更充分地了解基因及其編碼的蛋白質(zhì)[24]。生物信息學(xué)預(yù)測(cè)工具或軟件等作為指引科研實(shí)驗(yàn)方向的輔助工具,其結(jié)果必須經(jīng)過(guò)實(shí)驗(yàn)研究進(jìn)一步證明[25-26]。目前常借助一些報(bào)告基因如雙熒光素酶報(bào)告基因(luciferase)、綠色熒光蛋白報(bào)告基因(enhanced green fluorescent protein,EGFP)等瞬時(shí)表達(dá),根據(jù)報(bào)告基因的表達(dá)情況,確定啟動(dòng)子中的一些相應(yīng)元件的部位、功能及特性[18,27]。
對(duì)于家蠅分子生物學(xué)的研究與蠶、果蠅等昆蟲(chóng)相比,其理論研究不夠深入,比如基因組特點(diǎn)、功能等,基因的表達(dá)調(diào)控是其發(fā)揮生物功能、調(diào)節(jié)機(jī)體生命過(guò)程的基礎(chǔ),而啟動(dòng)子是表達(dá)調(diào)控的關(guān)鍵,構(gòu)建新的家蠅啟動(dòng)子片段報(bào)告基因載體是成功進(jìn)行轉(zhuǎn)基因家蠅研究的必要條件。本研究將Actin-5C基因生物信息學(xué)分析得到的結(jié)果,根據(jù)基因啟動(dòng)子的分析,克隆并成功構(gòu)建不同啟動(dòng)子片段報(bào)告基因載體,為分析家蠅轉(zhuǎn)基因載體的表達(dá)特征、家蠅轉(zhuǎn)基因細(xì)胞系的建立以及轉(zhuǎn)基因家蠅的養(yǎng)成奠定了基礎(chǔ),為研究Actin-5C基因啟動(dòng)活性及轉(zhuǎn)錄調(diào)控元件,探討基因表達(dá)調(diào)控機(jī)制提供了實(shí)驗(yàn)依據(jù)。轉(zhuǎn)基因家蠅是一個(gè)大工程,啟動(dòng)子效率的研究尤為重要,實(shí)驗(yàn)至此構(gòu)建的3個(gè)重組質(zhì)粒載體,初步檢測(cè)均具有活性,為今后實(shí)驗(yàn)篩選出最強(qiáng)啟動(dòng)子提供了條件,具有重要意義。
