微小膜殼絳蟲感染對ICR小鼠小腸干細胞相關基因mRNA表達的影響*
2021-04-13 07:10:06楊漢蕾趙敬張科鄢宇航門萬琪皮煥婷牟榮
貴州醫科大學學報 2021年3期
關鍵詞:小鼠
楊漢蕾,趙敬,張科,鄢宇航,門萬琪,皮煥婷,牟榮*
(1.貴州醫科大學 基礎醫學院 人體寄生蟲學教研室,貴州 貴陽 550004;2.貴州醫科大學 現代病原生物學特色重點實驗室,貴州 貴陽 550004;3.貴州醫科大學 臨床醫學院,貴州 貴陽 550004)
微小膜殼絳蟲(Hymenolepisnana,H.nana)是人類常見感染的絳蟲,其成蟲可寄生于人或鼠類小腸[1],能夠在一個宿主中完成整個生命周期,因此能夠自體感染[2]。近年來,有研究發現腸道寄生蠕蟲在破壞小腸組織完整性的同時也產生積極的損傷修復作用[3-4]。由小腸的穩態機制提示小腸干細胞在維持腸道正常生理功能及組織結構的重要作用[5],但關于腸道寄生蠕蟲感染與小腸干細胞的關系研究并不多見,因此本研究在前人的研究基礎上篩選出幾個腸干細胞相關基因,G蛋白偶聯受體5基因(leucine-rich repeat-containing G-protein coupled receptor5,Lgr5)[6]、Bmi1多梳復合蛋白基因(Bmi1 proto-oncogene polycomb ring finger,Bmi1)[7]、Msi1武藏RNA結合蛋白基因(musashi RNA-binding protein 1,Msi1)等干細胞標記物[8]及淋巴細胞抗原6復合物基因(lymphocyte antigen 6 complex locus A,Ly6a)[9-10]。Lgr5蛋白在體內分布廣泛,如腦、眼、脊髓、卵巢及胃腸道等都有一定的表達[11];Lgr5在器官、胚胎和其他一些生理過程中,以及成體干細胞的自我更新和維持中起著非常重要的作用,其在小腸和結腸隱窩基底柱狀上皮細胞中特異性表達,基因編碼產物為陽性的細胞,可以分化產生腸上皮所有的細胞類群[12];經研究證實Lgr5為小腸和結直腸干細胞標記基因,表達量的增加能夠促進胃腸黏膜的自我更新能力[6]。Bmil是最早發現的+4位腸干細胞標記基因,是Polycomb基因家族的一部分,該基因在胎盤、大腦及心臟中廣泛表達,對干細胞的自我更新以及腫瘤的發生起著重要作用[13-14];通過譜系追蹤技術證明,Bmi1能標記少量慢速增殖的+4位腸干細胞,這些細胞可增殖、擴張及自我更新,形成所有已分化的小腸上皮細胞譜系[7];有研究認為Bmil干細胞誘導腸上皮損傷后修復[15]。Msil是一種進化十分保守的RNA結合蛋白,在維持干細胞自我更新、分化和腫瘤發生方面起著重要作用[16];在哺乳動物小腸中,Msi1在距隱窩基底部4~6個細胞處表達,與公認的小腸干細胞位置相符,Msil在哺乳動物體內參與了包括維持干細胞內穩態、細胞分化及腫瘤形成等一系列的調控[17],因此Msil也可以作為小腸干細胞的標志物。Ly6a編碼的干細胞抗原-1(stem cell antigen-1,Sca-1)在化學性結腸炎模型的研究中被證明起動態調節作用,相比于正常腸組織,Sca-1表達并不明顯,而在損傷腸組織中,腸上皮修復相關的腫脹區域Sca-1表達水平顯著增高[18];此外,當損傷部位僅在絨毛而不涉及隱窩時,Ly6a表達并不上調[3];因此,Ly6a一般被當做腸道隱窩損傷的特異性標志物。本實驗擬在建立H.nana實驗感染動物模型的基礎上,采用實時熒光定量聚合酶鏈式反應(quantitative real-time polymerase chain reaction,qRT-PCR)技術檢測感染H.nana的ICR小鼠腸組織干細胞相關基因Lgr5、Bmi1、Msi1及Ly6a的表達,為探索H.nana感染所致宿主腸壁損傷后的修復機制提供有意義的基礎資料。
1 材料與方法
1.1 材料
1.1.1蟲體來源和實驗動物 蟲體采自貴陽花溪山區野生小鼠,經形態學觀察和分子生物學鑒定,確定采集蟲體為H.nana,收集成蟲孕節及蟲卵。6周齡美國癌癥研究所(institute of cancer research,ICR)雌鼠60只,購自貴州醫科大學實驗動物中心,體質量26~28 g,體健,經糞檢和間接凝集實驗證實無寄生蟲感染;稱重后隔離飼養觀察3 d,無健康問題后可作實驗用;實驗期間小鼠嚴格隔離飼養,水瓶與鼠糧不能交叉污染,每周清洗更換3次飼養籠及飼料、墊料。
1.1.2主要試劑和儀器 Trizol(美國invitrogen),qRT-PCR引物(上海生工生物工程有限公司合成);Nikon科研級成像系統(日本Nikon,Nikon DS-RI2),ABI7300 熒光定量PCR儀(美國 ABI,7300),生物組織切片機(德國徠卡,RM2016)。
1.2 方法
1.2.1H.nana形態學鑒定及分子生物學鑒定 取出野生小鼠小腸內獲取的的蟲體置于培養皿,生理鹽水沖洗3次,取載玻片滴入生理鹽水,用棉簽將蟲體挑到玻片上,使蟲體舒展開,置于400倍顯微鏡下觀察成蟲及蟲卵的形態;根據GenBank中記錄的COX1全基因序列設計保守引物[19-20],提取蟲總DNA,以其為模板利用保守引物擴增PCOX1,引物序列:PCOX1 Forward Primer為5′-ACCGCGTCGTGTGTGTATTT-3′,PCOX1 Reverse Primer為5′-ACATGCAACTGGGCTCATACG-3′,擴增產物預測大小202 bp。使用1%的瓊脂糖凝膠電泳35 min,凝膠成像系統拍照。將PCR 產物送上海生工公司進行測序,然后將測得序列與GenBank中記錄的COX1序列進行blast比對分析。
1.2.2實驗動物感染、分組及標本采集 60只ICR小鼠隨機均分為實驗組和對照組,H.nana蟲卵淘洗定量后,實驗組ICR小鼠灌胃500個蟲卵,對照組小鼠灌胃等量生理鹽水。感染后第1天開始,每天分別隨機處死實驗組小鼠和對照組小鼠各3只,取距離回盲部6~8 cm處小腸組織,同時記錄實驗組小鼠小腸組織內蟲體檢獲情況并計算蟲體檢獲率[蟲體檢獲率=(當天檢獲蟲體的小鼠數/3)×100%],共10 d。
1.2.3小鼠小腸目的基因的qRT-PCR檢測 利用 Primer 5.0 和 DNA Club 軟件根據Lgr5、Bmi1、Msi1及Ly6a各基因的 cDNA 序列設計 qRT-PCR特異性引物,同時以GAPDH基因為內參基因(引物序列見表 3),交由上海生工生物公司合成。反應體系為SYBR Premix Ex Taq Ⅱ(2×)10 μL,上游引物 0.8 μL,下游引物 0.8 μL,ROX Reference DyeⅠ(50×)0.4 μL,c DNA 1.0 μL,ddH2O 7.0 μL。反應條件為95 ℃預變性30 s、95 ℃變性5 s及60 ℃退火34 s,共40個循環,最后72 ℃延伸30 s。采用2-ΔΔCT法計算各目的基因的相對表達量。

表1 qRT-PCR 引物及產物大小Tab.1 Primers used in qRT-PCR
1.3 統計學分析
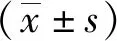
2 結果
2.1 H.nana形態學鑒定及分子生物學蟲種鑒定
顯微鏡下觀察野生小鼠小腸內獲得的蟲體,可見蟲體頭節有4個吸盤以及一個可自由伸縮的頂突;劃破孕節使蟲卵逸出,可見蟲卵卵殼較薄,胚膜兩端凸起發出絲狀物,內有一六鉤蚴(圖1)。提取該蟲總DNA,以其為模板利用保守引物擴增PCOX1,結果在200 bp處有一特異性條帶,與202 bp的預期結果大致相符(圖2);PCOX1序列與 GenBank 中登錄的COX1參考株核苷酸序列同源性為99%(圖3)。
圖3 COX1基因擴增產物同源性分析結果Fig.3 Homology analysis of COX1 amplified products
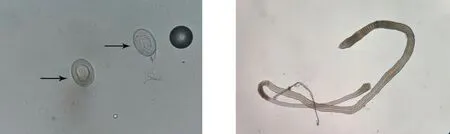
蟲卵(400×) 成蟲(40×)注:箭頭所示為蟲卵。圖1 野生小鼠體內檢獲H.nana蟲卵及成蟲的形態學特征(糞便生理鹽水涂片)Fig.1 Morphological characteristic of H.nana worms from wild mice(fecal physiological saline smear)
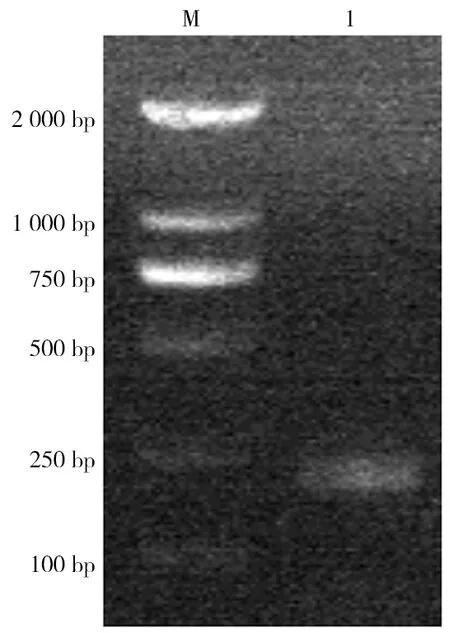
注:M為DNA分子質量標準,1為COX1核酸擴增產物。圖2 COX1基因擴增產物電泳結果Fig.2 Electrophoretogram of COX1 amplification products
2.2 H.nana蟲卵實驗感染小鼠
感染后第7~10天,實驗組小鼠距離回盲部6~12 cm處小腸內均檢獲白色細小蟲體,鏡下可分辨出有4吸盤的頭節和頸部(圖4),蟲體檢獲率100%;實驗組小鼠小腸內均未檢獲蟲體。
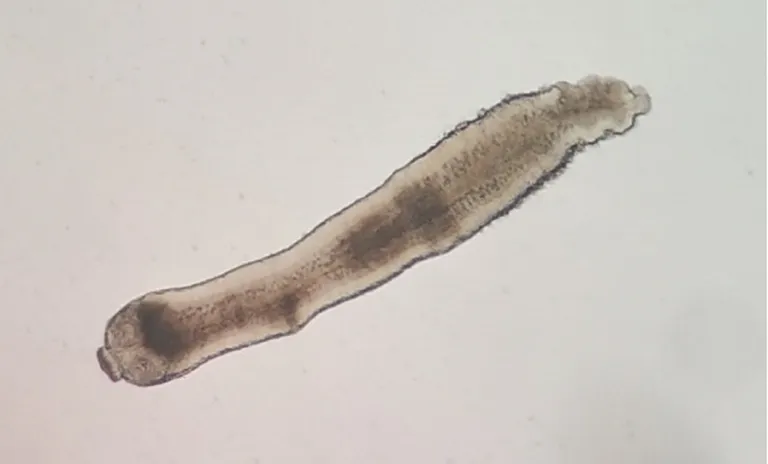
圖4 H.nana感染小鼠后第7天小鼠小腸內檢獲的蟲體(生理鹽水涂片,×40)Fig.4 The worm in the small intestine of mice infected with H.nana on the 7th day(Saline smear,×40)
2.3 qRT-PCR檢測ICR小鼠小腸組織干細胞相關基因mRNA的相對表達
感染H.nana后第1~10天,實驗組和對照組ICR小鼠干細胞相關基因mRNA的相對表達量結果顯示,與對照組相比,實驗組ICR小鼠Lgr5 mRNA在感染后第1天(即H.nana似囊尾蚴期)有一過性的高表達(P<0.05)、感染后第5~8天即H.nana成蟲期表達明顯下調(P<0.01);與對照組相比,實驗組ICR小鼠Bmi1 mRNA在感染后第1天(即H.nana似囊尾蚴期)有一過性的高表達(P<0.05)、感染后第6~8天即H.nana成蟲期表達下降(P<0.05);實驗組ICR小鼠Msi1 mRNA在感染后第一天表達有一過性上升但差異無統計學意義(P>0.05),第2~3天和第6~10天即H.nana似囊尾蚴期和成蟲期表達均下調(P<0.05);與對照組相比,實驗組ICR小鼠Ly6amRNA在感染后第2天(即H.nana似囊尾蚴期)表達呈一過性下降(P<0.05)、感染后第6~10天(即成蟲期)表達明顯上升(P<0.01)。見圖5。
3 討論
H.nana是一種古老的人獸共患寄生蟲,蟲體頂突上有一圈單層排列的小鉤,該結構幫助其牢牢附著在宿主腸壁組織上,從而給宿主造成嚴重的機械損傷,成蟲附著的部位,腸黏膜發生壞死,深達肌層,并有淋巴細胞和中性粒細胞浸潤[2]。研究發現腸道寄生蠕蟲可能具有誘導腸隱窩的新機制的損傷修復方式[21]。Nusse等[3]在使用多形類卷體線蟲(Heligmosomoidespolygyrus)感染小鼠發現幼蟲肉芽腫上腸隱窩增生明顯,但是腸干細胞基因如Lgr5表達下降,該過程與腸隱窩損傷標志物Sca1表達顯著增加有關。本實驗發現H.nana感染ICR小鼠后似囊尾蚴期編碼組織損傷因子Sca-1的基因Ly6a表達有一過性下降,腸干細胞相關基因Lgr5和Bmi1表達上升,而在成蟲期Ly6a表達持續上升,腸干細胞相關基因表達Lgr5、Bmi1及Msi1均顯著下降。H.nana的致病作用主要是由于成蟲頭節上的小鉤和體表微毛對宿主腸壁的機械損傷以及蟲體的毒性分泌物所致[2]。動物實驗結果顯示從感染第6天開始,H.nana的似囊尾蚴穿破腸絨毛回到腸腔,以頭節吸盤固著在腸壁上,逐漸發育為成蟲,對腸黏膜造成機械損傷,引起炎癥反應,所以推測感染后第6~8天Ly6a基因呈現明顯的高表達,可能與似囊尾蚴穿破腸絨毛回到腸腔引起腸黏膜的炎癥反應密切相關。
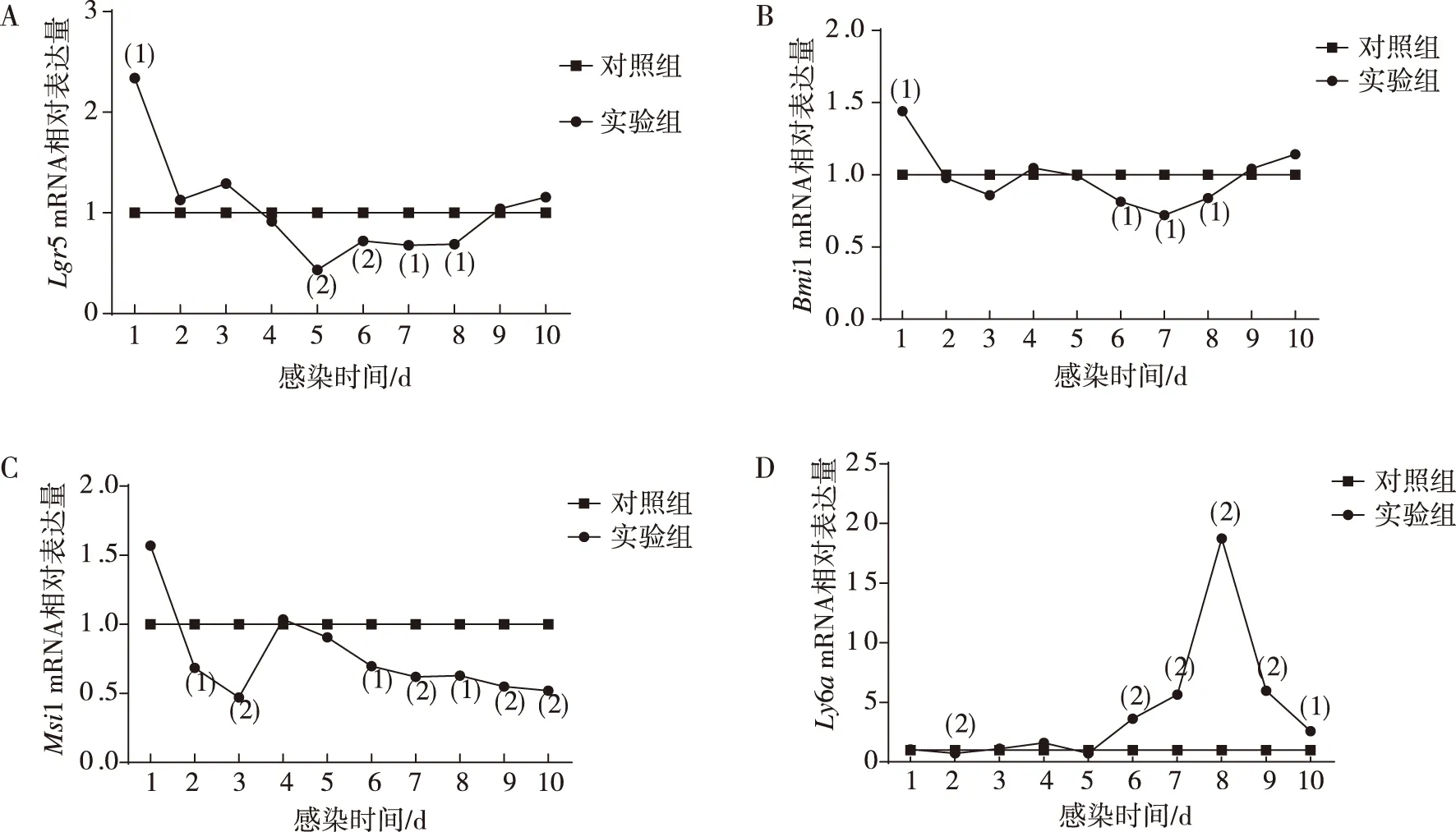
注:A、B、C及D分別為Lgr5、Bmi1、Msi1 及Ly6a mRNA相對表達量;與對照組比較,(1)P<0.05,(2)P<0.01。圖5 ICR小鼠小腸中干細胞相關基因mRNA的相對表達Fig.5 Relative mRNA expression of stem cell-related genes in small intestine of ICR mice
本實驗發現在感染成蟲期,無論是快速增殖干細胞基因Lgr5,還是緩慢增殖干細胞基因Bmi1和Msi1表達均顯著下降。Lgr5+干細胞是位于腸隱窩基底的快速增殖干細胞,其在腸道正常的生理代謝以及損傷修復過程中發揮著重要作用,當腸粘膜受到損傷時,Lgr5+干細胞迅速作出反應,快速增殖分化為各種腸上皮細胞類型,對腸上皮損傷進行損傷修復[5]。持續的損傷造成Lgr5+腸干細胞數目減少,這也許是快速增殖腸干細胞的一個特性。有研究發現在硫酸葡聚糖(disuccinimidyl suberate,DSS)誘導腸道炎癥的早期階段,Lgr5+腸干細胞的增殖會顯著加快,其分化的子代細胞增多,而在炎癥后期,小腸和結腸的Lgr5+腸干細胞的數目顯著減少[22]。當損傷較重Lgr5+腸干細胞不足以補充腸組織正常結構時,Bmi1+干細胞和Msi1+干細胞快速增殖,產生新的快速增殖干細胞,從而對腸上皮進行修復[23]。本實驗結果顯示Bmi1和Msi1在成蟲期表達顯著下降,推測在H.nana感染ICR小鼠損傷修復過程中也存在其他未知因子發揮作用,抑制了干細胞基因的表達,具體機制有待進一步研究。有研究發現在腸道受輻射誘導的炎癥階段Msi1和Msi2表達上升[24],與本研究正好相反,可能是由于損傷誘導方式的不同以及Msi1在不同組織表達模式不同所致。推測輻射誘導的損傷腸道涉及基因突變,為避免將突變基因遺傳下去,休眠的緩慢增殖干細胞會被激活,而H.nana感染主要對腸道造成機械性損傷,損傷修復模式及主要應答細胞也可能不同[25]。
綜上所述,本實驗發現H.nana感染可明顯影響小鼠小腸組織Lgr5、Bmi1、Msi1及Ly6amRNA水平,基因表達差異主要分2個階段,分別對應H.nana發育的似囊尾蚴階段和成蟲階段。下一步可使用條件性基因敲除或敲除技術,對干細胞相關基因在H.nana感染ICR小鼠的腸道損傷修復過程中所起的調節作用作進一步研究。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
