基于角尺度的隨機體在森林穩定性維持中的作用*
2021-04-02 04:40:36惠剛盈趙中華張弓喬胡艷波
林業科學 2021年2期
關鍵詞:物種
惠剛盈 趙中華 張弓喬 胡艷波
(中國林業科學研究院林業研究所 國家林業和草原局林木培育重點實驗室 北京 100091)
森林群落穩定性維持機制是森林生態學研究的熱點和核心問題。穩定性概念源于熱力學理論,指對于外界干擾某個系統表現出的維持現狀與恢復原狀的潛力(Hurdetal.,1971),這里所說的群落穩定性是指群落在達到演替頂極后出現的能夠進行自我更新并維持其結構和功能長期保持在一個較高水平、波動較小的現象。頂極群落是生物與環境在長期相互作用演變過程中相互適應和協調統一的產物,具有維持其結構和功能相對不變的穩定性。
一個多世紀以來,已有眾多關于群落穩定性維持的理論先后被提出和驗證。MacArthur(1955)提出了著名的多樣性-穩定性假說,認為群落多樣性和食物網復雜性會促進生態系統穩定性,群落多樣性可為群落穩定性提供強有力的保障后盾,在一個群落中物種越多,其抵抗外界干擾的能力越強,恢復能力也越強。多樣性-穩定性關系的實質是物種冗余所致,由此產生了穩定性的冗余(redundancy)理論(Lawtonetal.,1994; 黨承林,1998),該理論來源于自動控制系統可靠性理論。在生態學中,Odum(1983)定義的冗余指超過1種以上的物種或成分具有執行相同特定功能的能力。Walker(1992)最初提出冗余種的概念是針對評價優先保護地區或優先保護物種的,認為冗余程度低的物種,一旦滅絕,對穩定性的影響十分明顯,同時建議在確定冗余種時使用“功能群”概念,相同功能群的種可以在功能上相互替代,也就是冗余效應,當某一功能群僅剩單個物種時,此物種若從系統中損失將會導致系統功能遭受無法挽回的破壞,冗余種被認為是維持一定穩定性的“必要的”保險(Walker, 1995; Naeem,1998)。Janzen(1970)和Connell (1971)認為宿主專一性天敵在森林中具有隨距宿主距離增加而降低的不均勻分布特點,導致靠近同種母體周圍的更新體更易受天敵危害而具有較高的死亡率(距離制約性死亡)。這就是著名的Janzen-Connell假說:首先,母樹周圍的同種幼苗增補受到抑制,為其他物種定植提供空間,從而促進森林群落中物種共存; 其次,寄主高密度地點能夠吸引天敵并有利于天敵的繁殖、傳播,導致更新體具有較高的死亡率(密度制約性死亡)。在這一背景下,出現了另一個重要的生態學假說,即物種異群保護假說(herd immunity hypothesis),該假說認為異種鄰體能夠阻礙寄主專一的有害生物傳播,目標個體周圍異種鄰體增加會提高其存活率。Ford(1975)、Weiner等(1984)證明了同種負密度制約和異群保護假說如何影響種群的自然發展,認為許多群落自然發展的過程是,期初林地被一些先鋒樹種侵占,根據同種負密度制約假說,先鋒樹種的優勢木周圍會排除同種個體,進而被其他樹種代替,而這些先鋒樹種周圍的其他樹種常為小樹。Pommerening等(2017)和Wang等(2018)指出大樹個體周圍常具有更高的混交度,并稱之為混交度-大小假說(Pommereningetal.,2020),其本質是從相鄰木關系角度詮釋天然混交林特征,進一步印證了前人的假說。此外,物種間競爭可通過生態位分化來降低競爭緊張度而使更多物種共存。群落中的物種之所以能夠長期穩定共存,是因為不同物種分別占據不同生態位,生態位互補性使不同樹種共享地上和地下資源(Huietal.,2014; Belloetal.,2019),避免或延緩物種間的競爭排除作用,生態位相同或重疊程度較大的物種會因競爭有限的資源而發生競爭排除,不能穩定共存(牛克昌等,2009)。
上述假說的某些方面在試驗中得到了證實,雖然許多生態過程仍然未知,但已暗示在穩定性研究中需要關注同種負密度制約、異群保護、生態位互補性以及物種多樣性和冗余效應等方面。天然林是森林生態系統的主體,其結構基本上是物競天擇而形成的進化且健康的結構,具有較高的生物多樣性、較復雜的群落結構、較豐富的生境特征和較好的生態系統穩定性,在保障農牧業生產、維持生物多樣性、保護生態環境、減緩自然災害、調節全球碳平衡和生物地球化學循環等方面發揮著極其重要和不可替代的作用(唐守正等,2000)。本研究試圖精細解譯天然林空間結構并挖掘其普適性規律來詮釋森林穩定性維持機制,以期為改善我國人工林穩定性提供科學參考。
1 數據
本研究采用我國不同緯度帶的11塊天然林長期定位試驗數據(表1)。樣地詳細情況參見文獻Zhang等(2018)。
2 方法
本研究利用林分空間結構參數——角尺度進行林木格局解析(惠剛盈等,2003)。為描述相鄰木圍繞參照樹的均勻性,惠剛盈等(1999)、Hui等(2002)提出了角尺度(Wi)概念。角尺度有5種取值,分別對應參照樹相鄰木的分布形式(圖1)。根據參照樹及其最近4株相鄰木構成的結構單元中相鄰木的分布形式定義為結構體,任意一個結構單元均由5 株林木即1株參照樹與其4株最近相鄰木組成。角尺度Wi=0或0.25的林木稱為鄰體均勻分布的參照樹,簡稱均勻木,相應的結構單元稱為均勻體; 角尺度Wi=0.75或1.00的林木稱為鄰體聚集或團狀分布的參照樹,簡稱聚集木,相應的結構單元稱為聚集體; 角尺度Wi=0.50的林木稱為鄰體隨機分布的參照樹,簡稱隨機木,相應的結構單元稱為隨機體(Zhangetal.,2018)。據此,任何一個森林群落最多由3種結構體組成,即均勻體、聚集體和隨機體(惠剛盈,2020)。

表1 樣地及其林分概況Tab.1 General situation of sample plots and stands
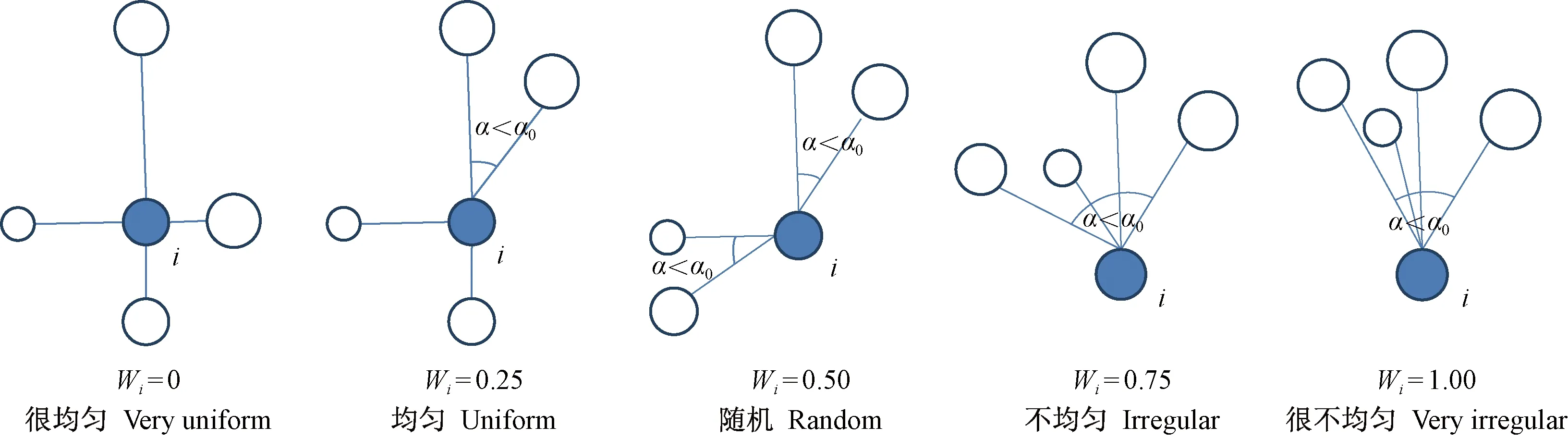
圖1 角尺度的取值及其意義Fig. 1 The value and significance of uniform angle index
運用R軟件編程,統計3種結構體的株數頻率及其斷面積占比。為消除邊緣效應,每塊樣地均設置5 m緩沖區,處于緩沖區的相鄰最近樹僅作為潛在的鄰體,不參與統計。
3 結果與分析
3.1 均勻木、隨機木和聚集木的株數占比
統計不同角尺度取值的林木數量,獲得林分角尺度分布,見圖2。由圖2可知,11塊天然林樣地林木角尺度分布呈現出一致規律:1) 林木角尺度基本為鐘型分布(近似正態分布),峰值均出現在Wi=0.50,即隨機木占多數; 11塊樣地中參照樹鄰體隨機分布的林木均占半數以上,比例最高的甚至達62%左右(C4、C6和C9),最低的也可達55%以上(C7和C8); 2)Wi=0和Wi=1.00,即非常均勻和非常聚集的林木都很少,其中非常均勻分布情況最不常見,在所有試驗樣地中最高的也只有1%,有5塊樣地中的比例均為0; 非常聚集分布次之,占全部林木的10%以內; 3)Wi=0.25(均勻)和Wi=0.75(不均勻)的林木一般在10%~30%之間。

圖2 天然林樣地的角尺度分布Fig. 2 Uniform angle index distribution of natural forest sample plots
3.2 均勻木、隨機木和聚集木的斷面積占比
林木株數比例表達結構體分布的數量特征,而斷面積占比在一定程度上可體現結構體分布的質量特征。分別統計均勻木、隨機木和聚集木斷面積占林分總斷面積的比例,得到不同結構體的斷面積分布見圖3。由圖3可知,林分中隨機木斷面積遠高于均勻木和聚集木,在所有樣地中均占林分總斷面積的50%以上,最大可達62%左右(C6和C9); 而均勻木和聚集木斷面積分別占林分總斷面積的10%~30%以內,2類林木斷面積總量基本小于林分總斷面積的半數。
3.3 結構體形成機制分析
基于角尺度揭示的天然林中隨機木、均勻木和聚集木的分布規律,既與天然林地域分布和森林類型無關,也與天然林樹種組成和格局類型無關。天然林林分中主要群體為隨機分布的林木個體(隨機木),這一現象出現必有其內在的形成機制。
對于聚集體來說,相鄰木擁擠在一起,使得中心木三面受光,可獲得更多的營養空間、更大的樹冠,甚至更高的生產力。相鄰大樹與小樹之間的不平等將導致非對稱競爭,尤其在相鄰木相對擁擠時,競爭加劇不僅導致小徑木生長放緩,而且相對弱勢的相鄰木可能在自然選擇過程中死亡,此時聚集體瓦解,原結構體內剩余林木將與其他林木重新組合構成新的結構體,林木越聚集,越容易發生瓦解和重組(圖4上)。而均勻體則相反,分布于均勻木四周的相鄰木分別占據3~4個方位,從而使中心木至少受到來自3個方向的擠壓或遮擋,造成中心木承受更大的競爭壓力,如果均勻木處于相對劣勢,則更容易在自然選擇過程中被淘汰,此時均勻結構體瓦解,原結構體內剩余林木將與其他林木重新結合成為新的結構體,林木越擁擠,越容易發生瓦解和接踵而來的重組(圖4下)。
不同于聚集體和均勻體,隨機體中心木(隨機木)具有兩面受光的可能性,相較于均勻木,參照樹競爭壓力小,而隨機木的鄰體之間擠壓程度較聚集體更小,因此隨機木及其鄰體相對來說承受的生存壓力較小,這種結構體更不容易由于競爭緣故而導致弱勢或不健康的林木頻出,比其他2種結構體中的林木有更大的可能性穩定生長,從而在自然演替過程中獲得更多的存活幾率。
4 討論
基于4株相鄰木空間關系的林分空間結構參數——角尺度,將森林群落中的林木分為隨機木、均勻木和聚集木,依次對應5株樹組成的隨機體、均勻體和聚集體(Zhangetal.,2018; 惠剛盈,2020),這一基于角尺度的森林結構解譯拓寬了森林培育新視野。目前,已有許多基于角尺度進行林木分布格局的研究(Aguirreetal.,2003; 安慧君,2003; 惠剛盈等,2003; 夏富才,2007; 趙中華等,2009),其中,最有代表性的是惠剛盈等(2003)利用Winkelmass軟件模擬產生了1 000個林木格局隨機分布的林分,發現格局隨機的林分角尺度分布為正態,其角尺度均值為0.496±0.007。隨后,許多有關森林群落林木分布格局的研究均證實天然林林分平均角尺度接近0.50(胡艷波等,2003; 趙中華等,2009; 李紀亮,2008; 鄭麗鳳等,2009; 趙洋毅等,2012; 彭輝,2017)。近年來,隨著研究不斷深入,基于角尺度的林分空間結構分析不僅僅局限于用其均值進行格局判斷,更重要的是格局形成機制的探索,愈加重視角尺度分布的研究。基于角尺度分布的天然林格局研究發現,處于隨機分布(Wi=0.50)的林木(隨機木)數量(株數或斷面積)最多,占比在50%以上(Zhangetal.,2018; 陳科屹,2018; 萬盼,2018),本研究印證了該結果。惠剛盈(2020)進一步從森林群落重要特征如樹種組成(多度分布)、林分結構(直徑分布、格局分布、混交度分布、密集度分布)和林木競爭狀態(競爭分布)入手,對比分析了天然林整體與天然林群落中隨機木的狀態特征,結果發現二者具有高度相似性。可見,隨機木是天然林的核心,在群落中起主體作用。在人工林空間結構與天然林對比研究方面,最有代表性的是曹小玉等(2020)對中亞熱帶典型林分空間結構的分析,其在立地條件基本一致的人工針葉純林[10杉木(Cunninghamialanceolata)、10馬尾松(Pinusmassoniana)、10柳杉(Cryptomeriafortunei)]、人工闊葉純林[10木荷(Schimasuperba)]、人工闊葉混交林[7木荷+3閩楠(Phoebebournei)]、人工針闊混交林[7杉木+3木荷; 6馬尾松+4木荷; 7柳杉+3馬褂木(Liriodendronchinense)]和天然次生林[7青岡櫟(Cyclobalanopsisglauca)+2苦櫧(Castanopsissclerophylla)+1冬青(Ilexchinensis)]中共設置27塊30 m×20 m樣地,每種類型各重復3次,對林分平均角尺度的研究結果人工針葉純林為0.35±0.025、人工闊葉純林為0.28±0.03、人工闊葉混交林為0.40±0.02、人工針闊混交林為0.39±0.02、天然次生林為0.49±0.02,進一步證實人工林格局分布的均勻性和天然林格局分布的隨機性。人工林分布格局的均勻性也表現在角尺度分布的左偏性上(袁士云,2010; 惠剛盈,2012; 岳永杰,2015)。人工林造林之初,如果嚴格按照相等株行距進行規則造林,根據角尺度定義可知,其分布格局必然如圖5(惠剛盈,2020; 曹小玉等,2020; 岳永杰,2008; 馮宜明等,2018)中幼齡林階段的分布格局,絕大多數林木角尺度為0,林分平均角尺度接近0; 隨著林齡增加和林木生長,人工林郁閉,林木發生自疏,一些林木死亡,人工林林木角尺度出現0.25、0.50甚至是0.75的情況,林分角尺度分布逐漸趨向于倒“J”形。此后,隨著人工林林齡進一步增加和林分自疏或多次經營干擾,林分中林木角尺度為0.25、0.50、0.75或1.00的林木越來越多,人工林角尺度分布也呈現出左偏狀態,角尺度均值進一步增大(圖5中齡林和成熟林階段),雖在短期內還達不到像天然次生林一樣的角尺度分布(圖6)(Zhangetal.,2018; 陳科屹等,2018; 萬盼,2018; 胡艷波等,2003; 趙中華等,2009; 李紀亮,2008; 鄭麗鳳等,2009; 趙洋毅等,2012; 彭輝,2017),但隨著進展演替,其隨機成分一定能達到像天然林一樣的50%以上,只是時間問題。也就是說,人工林隨著林分生長發育階段演進,林分穩定性在增加的同時,角尺度分布將發生均勻木數量減少而隨機木數量增多這樣一個顯著的變化過程(圖5)。

圖5 人工林角尺度隨時間的變化過程Fig. 5 Change process of uniform angle index of plantation with time
現有的森林穩定性維持機制主要關注組成森林群落的物種生態特性,對人工林培育的啟示只能是營造樹種生態位不同的多優樹種混交林。本研究提出的基于空間結構的隨機體-穩定性假說,對人工林培育的啟示是隨機化造林,考慮到生產可行性,提倡控位造林(惠剛盈,2020)。人工林林木分布格局的規則性使其本身失去了自然屬性,而天然林林木分布格局的隨機性則促使其更加穩定。現有人工林不穩定的根源除了樹種組成單一外,主要是空間結構尤其是林木分布格局的均勻性所致,因此,提高現有人工林穩定性的關鍵是增加隨機體,采用隨機化經營(惠剛盈,2020)。
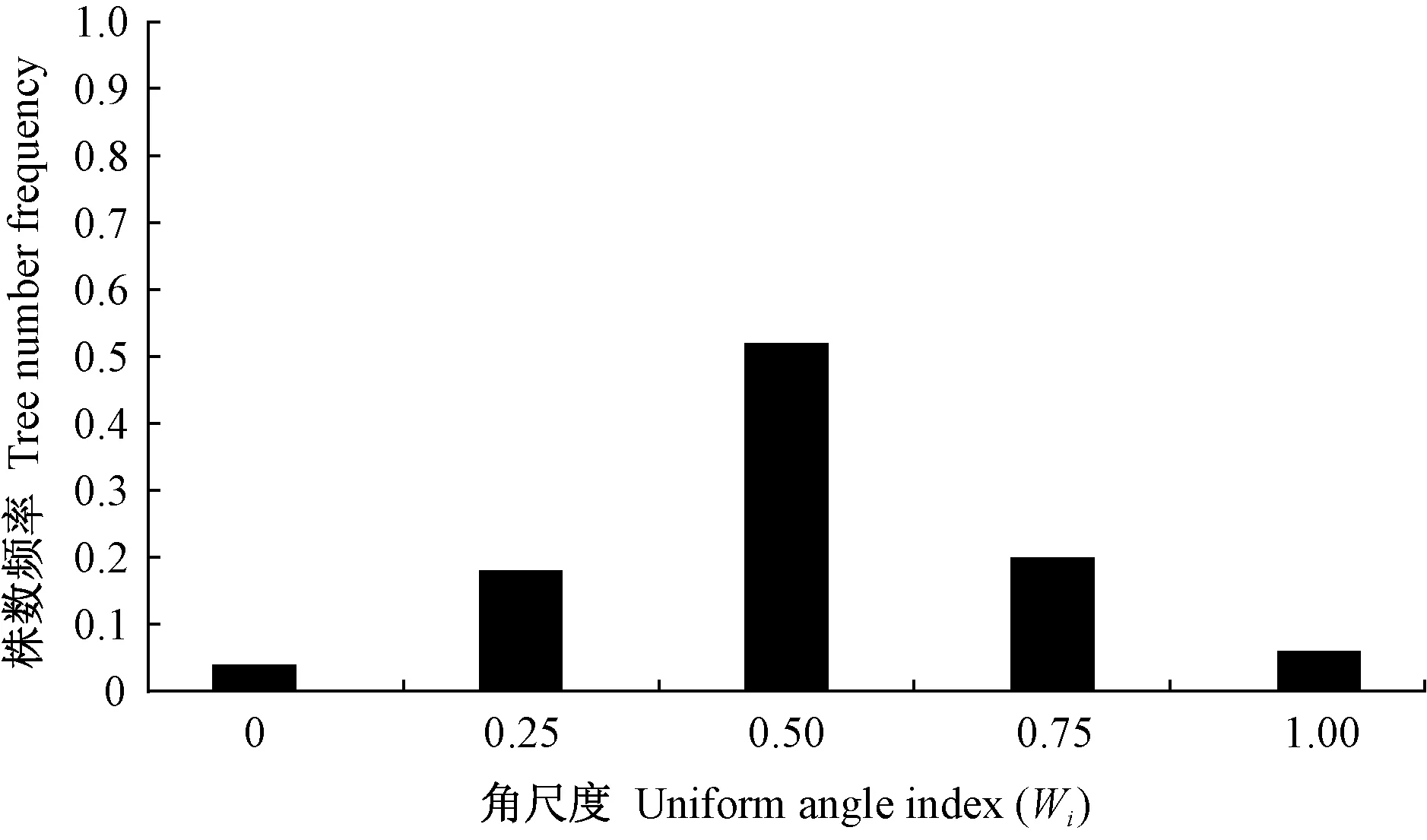
圖6 天然林的角尺度分布Fig. 6 Uniform angle index distribution of natural forest
5 結論
基于4株相鄰木空間關系的林分空間結構參數——角尺度,將森林群落中的林木分為均勻木(Wi<0.5)、隨機木(Wi=0.5)和聚集木(Wi>0.5)。天然林中主要的林木群體為隨機分布的林木個體(隨機木),隨機體作為可以長期存在且不易瓦解的結構體,可為林木提供長效、穩定的微環境,隨機體在天然林中的主體構成是促使森林穩定性的關鍵。
本研究提出森林穩定性維持的“隨機體-穩定性”假說,即林分中均勻體由于受四方的遮擋和擠壓,使得位于中心的均勻木對環境資源(光)的利用受阻,加大中心木死亡的幾率,導致結構體容易解體; 聚集體由于聚集木的鄰體相互擁擠從而可增大鄰體中個體死亡的幾率,也容易導致其結構體解體。林分中隨機體的架構形式使得隨機木具有兩面受光的鄰體微環境,相較于均勻體,其中心木受相鄰木的競爭壓力更小,而相較于聚集體,隨機木的鄰體之間擠壓程度則更小,隨機木及其鄰體(隨機體)相對來說承受的生存壓力較小,這種結構體比其他2種結構體中的林木有更大的可能性穩定生長,從而在自然演替過程中能夠獲得更大的存活幾率。隨機體被認為是林木在森林群落條件下最為普適的鄰體配置形式,植物由于位置的固定性,只有通過適者生存的自然選擇法則實現隨機體的形成; 隨機體數量越多,森林群落越穩定。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
