植物精油對真菌微生物抑制作用的研究進展
2021-04-01 03:27:14翟秀超馮文旭吳殿輝王璐陸健
食品與發酵工業 2021年6期
關鍵詞:植物
翟秀超,馮文旭,吳殿輝,王璐*,陸健*
1(無錫中糧工程科技有限公司,江蘇 無錫,214035)2(江南大學, 工業生物技術教育部重點實驗室,江蘇 無錫,214122) 3(江南大學, 糧食發酵工藝與技術國家工程實驗室,江蘇 無錫,214122)4(江南大學 生物工程學院,江蘇 無錫,214122)
谷物、果蔬等農作物目前正遭受著曲霉屬(Aspergillus)、青霉屬(Penicillium)和鐮刀菌屬(Fusarium)等多種真菌的污染。真菌侵染農產品后,在適宜的溫度、濕度和足夠的營養條件下開始萌發繁殖,并不斷分解農產品中的糖類、蛋白質等成分,造成營養物質損失,進而影響農產品的質量和產量,造成巨大的資源浪費和經濟損失[1]。并且真菌對農產品的侵染往往伴隨著其次級代謝產物——真菌毒素(mycotoxin)的產生,這嚴重威脅著人類的身體健康和生命安全。據糧農組織估計,全世界約有25%的作物受真菌污染并受真菌毒素影響,估計損失達數10億美元[2]。真菌污染對全球糧食生產及儲備造成了嚴重影響,引起了全球性關注。
目前常用的抑菌措施是噴灑化學殺菌劑,但是化學殺菌劑的濫用使得病原微生物的抗性逐漸增強,進而人們需要噴灑更多的化學殺菌劑,造成了惡性循環。而且很多化學殺菌劑不可生物降解,它們可以在土壤、植物和水中積累,并通過食物鏈影響人類。由于其較長的降解期和對人類健康的潛在影響(致癌性、致畸性、高殘留毒性和激素失衡等),允許采后施用的化學殺菌劑的類型和濃度受到限制。近年來,公眾對殺菌劑殘留污染的關注度明顯增加。考慮到這些因素,需要開發既有效又經濟可行的安全易降解替代品[3]。
精油(essential oils, EOs)是天然抗菌劑,多種植物均可產生,以保護自身免受感染,在食品工業和農業中,在對抗病原微生物方面具有很大的潛力[4]。一些精油,如丁香油,百里香油,茶樹油和肉桂油,具有廣譜抗菌性。同時精油具有生物降解性、高揮發性和低殘留等特點,不會像化學殺菌劑一樣對生態環境造成負面影響,因此精油可作為綠色殺菌劑的重要來源之一。精油通常是含有大約20~80種不同化合物的復雜混合物,萜烯、萜類化合物和酚類是其抑制微生物的主要成分。精油成分的不同導致不同精油具有不同的抗微生物活性和抑菌機理,而特定精油的抗微生物活性主要取決于其濃度和化學組成[5]。
為了使植物精油在農業和食品工業上更好的應用,本文概述了植物精油對真菌微生物的抑制作用和降低真菌毒素產生的作用,并對抗菌和抑制毒素的作用機制進行了匯總,最后介紹了精油目前在農業方面的應用研究及精油乳液的研究前景。
1 真菌及真菌毒素污染
真菌毒素中毒(mycotoxicosis)[6]概念的提出是在60多年前,然而當時并沒有人認為發霉的食物可以導致疾病,經過半個多世紀的深入研究,人們已經認識到真菌毒素是由多種真菌產生的具有不同毒性的次級代謝產物。常見的真菌毒素主要包括黃曲霉毒素(aflatoxin, AFs)、脫氧雪腐鐮刀菌烯醇(deoxynivalenol, DON)、雪腐鐮刀菌烯醇(nivalenol, NIV)、伏馬菌素(fumonisins, FBs)、赭曲霉毒素A(ochratoxins A, OTA)、T-2毒素(T-2 toxin)和玉米赤霉烯酮(zearalenone, ZEA)等。如表1所示,真菌毒素不僅具有極強的急性毒性,致畸性、致癌性和致突變性的“三致”作用,還具有肝臟毒性、細胞毒性和免疫毒性等,給人類的身體健康造成極大的威脅[1]。

表1 幾種常見的真菌毒素的產生菌種及毒性Table 1 Several common mycotoxin production strains and toxicity
目前在農業上控制真菌毒素的方法主要有物理方法、化學方法和生物方法3種。但是這3種方法各有各的缺陷[17],通過機械分離密度小或粒徑小的谷物、熱水處理和放射處理的方法來去除被毒素污染的谷物難以工業化應用;添加甲醛、雙氧水等化學試劑來處理糧食或者加入吡咯苯類、咪唑類等殺菌劑[1]則會造成環境污染問題;通過添加乳酸菌、白地霉等有益菌來抑制致病菌生長的方法也容易造成接種微生物的二次污染。因此,研究精油等綠色防霉劑來控制真菌和真菌毒素的污染在食品安全研究方面具有重要意義。
2 植物精油抗菌及降低毒素的研究
2.1 植物精油
精油是含有揮發性芳香化合物的濃縮油性液體,包括萜烯類,萜類化合物(氧化萜烯)和酚類等,揮發性芳香化合物大多衍生自植物材料,例如花,芽,葉和樹皮[5]。通常情況下,植物中的油含量<1%,一般需要通過提取、蒸餾和溶劑萃取獲得高度濃縮的精油,水蒸汽蒸餾是精油商業分離最常用的方法。植物精油目前由于其低殘留、易揮發、易降解及綠色無污染等特性逐漸應用于抑菌防霉劑領域,關于精油主要成分及其對真菌抑制作用的研究也越來越廣泛。
2.2 植物精油對真菌生長的抑制作用
具有抗微生物活性的常見精油及其主要成分見表2,精油的主要成分中抗微生物活性最高的是酚類物質,其次是萜類化合物。與酚類和萜類化合物相比,精油中的萜烯、酮類和酯類(如β-月桂烯,α-噻吩酮和乙酸香葉酯)的抑菌能力和生物活性較弱[18]。

表2 植物精油主要成分及其抗真菌活性和抑制毒素積累的情況Table 2 Main components of plant essential oils and their antifungal activities and inhibition of toxin accumulation

續表2
由表2可知,大部分精油都具有廣譜抗菌活性。以丁香油為例,它對鐮刀菌屬、曲霉屬等真菌都具有較好的抗菌活性。JUGLAL等[19]評估了9種植物精油對寄生曲霉和鐮刀菌(F.moniliforme)生長的影響,發現丁香油的抑菌作用大于肉桂精油、牛至精油和肉豆蔻精油,并且丁香油可以顯著降低AFs的形成(降低78%),并影響伏馬菌素的產生。SHARMA等[20]發現丁香油抑制尖孢鐮刀菌(F.oxysporum)生長的最小抑菌濃度為31.25 μg/mL,通過乳化的方法將丁香油包裹入納米乳液并用于番茄幼苗階段,有效控制了番茄中61.1%的由尖孢鐮刀菌引起的枯萎病。
2.3 植物精油抑制真菌產真菌毒素的作用
植物精油不僅對真菌生長有較好的抑制效果,而且在減少各種真菌毒素(AFs、DON、OTA和ZEA等)積累方面效果顯著。MARIN等[16]通過將精油直接添加到已知水分活度(water activity,Aw)的玉米粒中并接種禾谷鐮刀菌的方式進行抑制產毒實驗,發現丁香油在20 ℃,0.995Aw下可以完全抑制禾谷鐮刀菌產生DON,而且丁香油、香茅油和玫瑰草精油可以在30 ℃,0.950Aw下完全抑制禾谷鐮刀菌產生ZEA。
然而精油的水溶性相對較差,直接添加的方式往往存在著混合不均勻、利用率低等缺點,導致產毒實驗的重復性差。研究人員提出了多種添加方法(如蒸氣熏蒸、與吐溫等乳化劑混勻等),改善了產毒實驗的效果。(1)蒸汽熏蒸:袁媛[22]研究發現肉桂精油熏蒸玉米的處理,可以抑制黃曲霉的生長,顯著降低AFB1的積累,降幅達98.94%,而且肉桂油可以降解玉米粉中的AFB1,降解率達43.96%,這也為添加植物精油直接降解真菌毒素的應用提供了參考。(2)液體直接添加:BASILICO等[14]通過在補充酵母提取物液體培養基(yeast extract with supplement medium,YES)中接種赭曲霉孢子并加入各種植物精油進行產毒實驗,發現750 μg/mL的牛至油在14 d內完全抑制赭曲霉產生OTA,同濃度的薄荷油僅能在7 d 內完全抑制赭曲霉產生OTA,而1 000 μg/mL的牛至油和薄荷油在21 d內完全抑制赭曲霉產生OTA。(3)添加乳化劑:FERREIRA等[24]將生姜精油溶于0.001%的吐溫-80并接入PDA液體培養基中,發現500 μg/mL的生姜油對禾谷鐮刀菌產生DON的抑制率達到了47.3%,而在2 000 μg/mL時完全抑制了真菌生長和DON的產生。
2.4 植物精油抑制真菌的作用機制
從植物中提取的精油通常含有一些抗菌或抗氧化化合物,它們被認為是天然殺菌劑的良好選擇,精油的抗菌活性已在許多不同抗菌模型中得到證實[5],其抗菌機制主要取決于精油中存在的化學成分的濃度和類型。不同的化學成分可以通過不同的機制起作用,也可以靶向不同種類的微生物,并且不同的作用機制也可以相互影響,從而達到抑菌的目的。
2.4.1 增強細胞通透性
精油的親脂性和低分子質量組分容易穿過真菌細胞質膜,導致細胞膜滲透性和滲透平衡的破壞,從而導致細胞死亡,主要包括脂肪酸譜的改變和離子的泄漏。許多研究人員將精油的作用機制歸因于它們穿透細胞膜進入細胞內部的能力,這使得細胞結構發生變化,周圍的精油更容易滲透進入細胞[4, 29-30]。
(1)脂肪酸譜改變:萜類的抗菌機制發生在細胞膜水平,其抗微生物活性歸因于它們具有抑制呼吸和其他能量依賴性過程的能力[31],然而,當作為單一成分時,萜類的總體抗微生物活性較低[29]。研究發現,萜類化合物的作用機制通常先是親脂性成分改變細胞膜脂肪酸組成,導致了膜滲透性的改變和細胞內物質的滲漏[32],進而使萜類化合物跨膜進入細胞內發揮其影響代謝的作用。
(2)離子泄漏:ULTEE等[33]研究發現香芹酚由于其較好的脂溶性可在細胞質膜中積累(最高可達6×104μg/g),引起細胞膜擴張,并導致膜不穩定及離子泄漏。另外,香芹酚通過將其羥基質子交換為另一種離子(K+等)而充當單價陽離子的跨膜載體,使質子釋放到細胞質內而降低質子動力(圖1),影響ATP的合成進而導致細胞死亡。

圖1 香芹酚降低質子動力示意圖[34]Fig.1 Schematic diagram of carotenol reducing proton dynamics
2.4.2 影響細胞結構及功能
據文獻報道,有些精油及其主要抑菌成分也會通過影響細胞結構及功能來達到抑菌的目的。羅曼[34]研究發現檸檬醛可以使菌絲體DNA、RNA、脂類和蛋白質等的生物合成受到抑制,進而影響相關基因的表達與調控,擾亂能量代謝與遺傳穩定性,最終導致細胞死亡。研究發現檸檬醛損傷黃曲霉細胞核DNA,導致局部斷裂,且其損傷程度與檸檬醛濃度呈正相關。檸檬醛通過損傷細胞壁和細胞膜上的功能脂質蛋白,導致膜對營養物質(如糖和蛋白質等)的吸收轉運率分別下降為23.3%和30.61%。此外,檸檬醛可以使黃曲霉體內的琥珀酸脫氫酶、蘋果酸脫氫酶活性分別呈不可逆趨勢下降為31.82%和24.2%,并隨著檸檬醛濃度的升高,2種酶的活性逐漸消失。
目前也有很多研究通過電子顯微鏡等手段直接觀察精油對真菌結構的損傷。SUN等[35]通過掃描電子顯微鏡和透射電子顯微鏡觀察肉桂醛對真菌菌絲體的損傷,發現肉桂醛可以破壞黃曲霉菌絲體并導致其內容物流失并破壞線粒體,如圖2與圖3所示。

a,d-對照真菌菌絲體在所有部位均呈正常和規則的管狀;b,e-用低濃度肉桂醛(52 μg/mL)處理的菌絲(形態上略有塌陷);c,f-用高濃度的肉桂醛(78 μg/mL)處理的菌絲(形態上明顯干癟有褶皺)圖2 肉桂醛對黃曲霉超微結構的影響(掃描電子顯微鏡)Fig.2 Effect of cinnamaldehyde on the ultrastructure of A.flavus (scanning electron microscope)
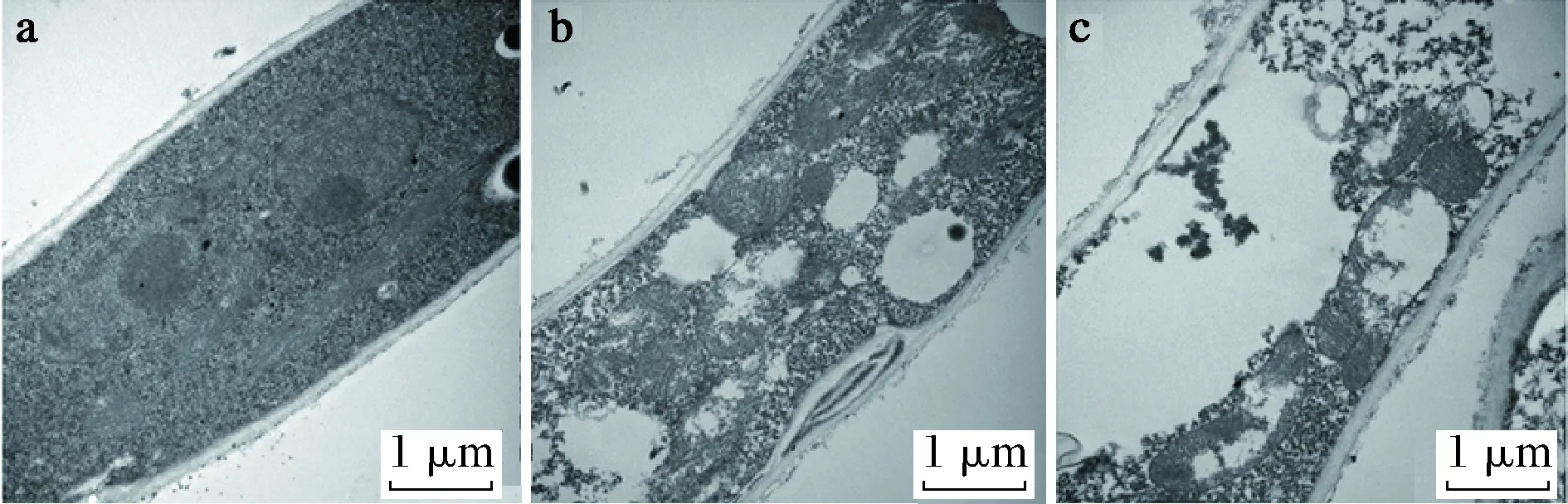
a-顯示均勻的細胞質和細胞內正常的細胞器結構(包括線粒體,細胞核和囊泡);b-添加52 μg/mL肉桂醛的菌絲;c-添加78 μg/mL肉桂醛的菌絲圖3 肉桂醛對黃曲霉超微結構的影響(透射電子顯微鏡)Fig.3 Effect of cinnamaldehyde on the ultrastructure of A.flavus (transmission electron microscope)
2.4.3 紊亂能量代謝
精油紊亂真菌能量代謝的作用主要通過損傷線粒體及抑制菌絲體DNA、RNA、脂類和蛋白質等的生物合成來實現。精油會對線粒體產生一定的破壞作用[34, 36],受損的線粒體DNA抑制電子傳遞蛋白的表達,進而導致線粒體內活性氧(reactive oxygen species, ROS)的累積。李金寒等[37]通過電化學傳感器實時監測和熒光分析觀察檸檬醛作用下胞內外ROS的變化,結果表明檸檬醛能夠刺激黃曲霉產生ROS,并在細胞內蓄積。ROS是氧代謝過程中產生的一系列中間產物,具有較強的氧化能力,能夠與生物體內的活性大分子物質(脂質、蛋白質等)發生反應,對其產生不可逆的氧化損傷,從而導致生物體損傷[37]。同時ROS可影響很多酶的活性從而產生轉錄水平上的抑制。
羅曼[34]的研究發現檸檬醛通過損傷膜功能脂質蛋白,降低營養物質轉運率,并可通過降低琥珀酸脫氫酶、蘋果酸脫氫酶的活性紊亂能量代謝。添加檸檬醛后以琥珀酸、α-酮戊二酸和丙酮酸為底物時所測定線粒體呼吸速率分別下降了24.1%,14.3%和36.1%,能量代謝發生紊亂并促進細胞死亡。
2.4.4 精油組分間的協同作用
精油是20~80種化合物組成的復雜混合物,精油各組分之間的相互作用可能會產生協同或拮抗的抗菌作用。例如萜類化合物的作用機制通常是與親脂性成分協同作用,親脂性成分破壞細胞膜后使得萜類化合物進入細胞發揮作用。
JIANG等[38]利用GC-MS分析了迷迭香精油的主要成分,評估了1,8-桉葉素(26.54%)和α-蒎烯(20.14%)及迷迭香精油對金黃色葡萄球菌、枯草芽孢桿菌、銅綠假單胞菌、大腸桿菌、白色念珠菌及黑曲霉等的最小抑菌濃度,發現對所有測試微生物而言,迷迭香精油的最小抑菌濃度為0.03%~1.0%,α-蒎烯的最小抑菌濃度為0.3%~4.0%,而1,8-桉葉素的最小抑菌濃度為0.2%~4.0%,證實了植物精油的整體抗菌活性比其單一抗菌成分更強。
此外,BAYDAR等[39]發現香薄荷精油的抗菌活性要高于野生牛至油,然而其主要抗菌物質香芹酚和百里酚的含量卻比牛至油的比例低,這是因為香薄荷含有更高濃度的ρ-傘花烴和γ-萜品烯。然而ρ-傘花烴和γ-萜品烯單獨測試時,它們對任何微生物都沒有抑制作用。這表明,精油中的微量成分通過協同作用對其抗菌活性產生重要作用。
不同精油協同作用的機理歸根于不同精油不同化學組成的協同作用,相似作用機制的精油進行復配可能并沒有好的協同作用,這也解釋了精油復配作為抑菌劑添加會產生協同或者拮抗作用的原因。
2.5 植物精油抑制真菌產生毒素的作用機制
關于植物精油抑制真菌產生真菌毒素的機理非常復雜多樣,通常認為在較高濃度下,植物精油主要通過抑制真菌生長而抑制真菌毒素的產生,而在較低濃度下,則通過下調毒素關鍵調控和合成結構基因的表達而抑制真菌毒素的生物合成。
FERREIRA等[24]以麥角甾醇的含量表征鐮刀菌生物量來研究生姜精油(ginger essential oil, GEO)對禾谷鐮刀菌生長及毒素產生的影響。結果表明,GEO在1 000 μg/mL的質量濃度下顯著降低麥角甾醇的含量,并且在高于2 000 μg/mL時達到99.9%的抑制率。但是,在500 μg/mL時,隨著麥角甾醇的含量增加,DON的含量反而下降47.3%。證實了在低濃度下,植物精油不通過抑制真菌生長而抑制毒素產生。LIANG等[40]的研究也發現肉桂醛(0.40 mmol/L)、丁香酚(0.80 mmol/L)和檸檬醛(0.56 mmol/L)在不抑制真菌生長的情況下顯著降低AFB1產量,抑制率分別為68.9%、95.4%和41.8%。
SUN等[35]的研究表明肉桂醛可以通過降低過氧化氫和還原型谷胱甘肽水平來改變細胞內氧化還原狀態進而抑制黃曲霉產生毒素。研究表明78 mg/L的肉桂醛使過氧化氫酶和谷胱甘肽過氧化物酶的活性分別上升了1.9倍和2.2倍,AFB1的含量降低了65.6%。
真菌毒素的產生受到相關基因的調控,SUN等[35]研究發現經肉桂醛短時作用(5 min)或長時作用(120 h)均能下調黃曲霉的黃曲霉毒素合成調控基因aflR的表達,并有明顯的濃度梯度效應。LIANG等[40]發現肉桂醛、檸檬醛和丁香酚可以通過下調黃曲霉YC-15的黃曲霉毒素生物合成基因aflR,aflT,aflD,aflM和aflP來降低AFB1的產生,0.80 mmol/L的丁香酚在7 d內下調了5種基因的表達量,分別減少了15、4.8、5.2、138和2 061倍,降低了89.2%~97.9%的AFB1的生物合成,肉桂醛和檸檬醛也不同程度的下調了5種基因的表達并降低毒素的產生。
3 植物精油抑制真菌微生物的應用及研究現狀
植物精油抑制真菌微生物的應用主要集中在糧食的貯存方面和果蔬保鮮方面,目前在玉米、小麥、大麥、花生、大豆、番茄、圣女果及草莓等方面均有深入的報道,并且已經有在農作物生長階段添加精油制劑抑制病原微生物生長的報道[20],這也是下一步研究的方向。另外,與體外模型系統相比,實際應用中往往需要更高濃度的精油才能達到相同的抗微生物活性[5]。而高濃度的精油會因其高揮發性,低溶解度、不穩定性和獨特的風味特征導致精油的利用率低而影響其抗菌活性。因此,近年來,人們對提高精油作為抗菌劑的抗菌活性的研究越來越多,包括制備微膠囊、納米乳液及脂質體等[41]。
3.1 植物精油的應用研究
3.1.1 糧食貯存
植物精油目前最具有實用性的應用是作為熏蒸劑進行添加。已經報道了在封閉容器貯藏時,孜然油和薄荷油可以保護>63%的小麥和鷹嘴豆樣品免受真菌侵害[42]。袁媛等[43]使用肉桂精油熏蒸玉米,20 d內完全抑制黃曲霉生長,且在玉米水分含量為13%時,對AFB1的抑制率達98.94%。同時,肉桂油可以降解玉米中的AFB1,降解率高達64.5%,這為植物精油的進一步應用提供了基礎。精油熏蒸糧食是在貯存階段防治真菌侵害比較好的方法,因為其價格低廉且低殘留(糧食曬干后可以最大程度上去除精油蒸汽)而廣受歡迎。
低劑量γ輻射在消毒和提高農產品的保質期方面已經得到世界衛生組織和聯合國糧食與藥物管理局的認可。將輻射與其他脫毒劑(如精油和化學殺菌劑)相結合是提高輻射殺菌效率有效創新的方法,組合方法減少了其所需的輻射劑量,并且可以在低輻射劑量下促進食品安全。KALAGATUR等[44-45]采用依蘭精油和印度香水姜精油結合γ輻射處理玉米,在1.89 mg/g香水姜精油和4.12 kGy輻射處理下觀察到禾谷鐮刀菌的生長,DON和ZEA的產生則被完全抑制;在2.5 mg/g依蘭精油和4 kGy γ輻射下完全抑制DON和ZEA的產生。
3.1.2 果蔬保鮮
植物精油在果蔬保鮮方面也有較出色的應用。徐仕翔[46]制備了平均粒徑11.9 nm的月桂精油乳液,將1 000 μg/mL的微乳液作用于櫻桃番茄,有效抑制了鏈格孢菌(Alternariaalternata)引起的灰霉病。與對照組相比,經月桂精油乳液處理后,抑菌率達33.4%。范小靜等[47]研究發現15%的檸檬油微乳劑對圣女果有明顯的保鮮作用,處理15 d后,壞果率僅為14.13%,比空白對照處理下降了62.89%。而且,檸檬油微乳劑可顯著延緩圣女果可溶性固形物、有機酸和VC含量的下降,貨架期可達到7 d。趙亞珠等[48]將百里香油制備成納米乳液涂布于紙箱用于儲藏草莓,4%的百里香精油抗菌紙箱可以明顯降低灰霉(Botrytisallill)、青霉、交鏈孢霉和根霉(Rhizopus)引起的草莓腐爛,并降低菌落總數,使貨架期延長1~2 d。
植物精油的引入可以較好地抑制真菌引起的果蔬腐爛,在延長果蔬貨架期方面有較好地潛力。不僅如此,植物精油也可以較好地控制果蔬生長階段的真菌病害。SHARMA等[20]制備了包括丁香油和檸檬草油的復合納米乳液,有效控制了番茄幼苗階段由尖孢鐮刀菌引起的番茄枯萎病,抑制率達70.6%。
3.2 精油乳液抑制真菌微生物及真菌毒素的前沿進展
WU等[49]以肉桂油為油相,以卵磷脂為乳化劑,添加中鏈甘油三酯(medium chain triglycerides,MCT)抑制奧斯特瓦爾德熟化(ostwald ripening),添加殼聚糖進行包裹來提升其穩定性,制得平均粒徑560 nm的納米乳液。抑菌實驗表明納米乳液對禾谷鐮刀菌孢子萌發的抑制率達100%,在產毒實驗中對DON和3ADON產生的抑制率分別達91.3%和70.0%。馮文旭等[50]將丁香油、肉桂油、牛至油及山蒼子油等分別制備成納米乳液,4種乳液對禾谷鐮刀菌菌絲生長及DON、15ADON的產生都具有較好的抑制作用,40 mg/g的牛至油乳液可完全抑制菌絲的生長,0.3 mg/g的肉桂油/MCT乳液對DON和15ADON的抑制效果可達98.8%和96.6%。
此外,精油制備成納米乳液后由于其優良的穩定性使得其在應用時有較好的緩釋作用。KALAGATUR等[51]利用殼聚糖包裹馬提尼精油制備納米乳液,納米乳液中的馬提尼精油在48 h內僅緩慢釋放了43.37%。研究表明殼聚糖包埋的方法可以將精油的揮發性醇和酚基團轉化為更穩定的絡合物形式,通過逐漸釋放抗真菌成分來增加其殺菌作用的有效作用時間。
3.3 精油應用的局限性
與體外模型相比,實際應用中往往需要更高濃度的精油才能達到相同的抑菌活性。比如,在半脫脂牛奶中約需要2倍的濃度,在豬肉香腸中約為10倍,在湯中則為50倍[4-5]。而高濃度精油的添加會因其高揮發性,低溶解度和不穩定性等導致其利用率降低甚至影響其抗菌活性。并且添加高濃度的精油可能會造成生物毒性或具有一定的刺激性,如丁香酚,薄荷醇和百里酚等會引起口腔組織刺激[5],因此精油在不同食品中允許使用的安全濃度需要進一步研究。
另外,精油常常有著令人不愉悅的刺激性氣味,過量添加可能會對相應食品的感官造成一定的影響。精油的添加對于草藥、香料及調味品等食品的氣味方面影響較小,對于一些需要進一步烹飪的肉類、魚類的影響也較小,在烹飪后即使有少量的殘留也不會對風味及外觀造成不良影響,且一些精油也會賦予這類食品一些較好的風味[5]。然而,精油應用于果蔬保鮮方面時,會與果蔬等本身的醇類、酯類等風味物質形成混合的芳香物質,影響了精油在食品行業的應用。植物精油與果蔬之間需要合理搭配來進行果蔬的品質(顏色、香氣等)控制[52],也應該充分評估植物精油的殘留導致的感官變化與精油的抑菌程度間的制約關系。
4 結論與展望
許多植物精油在體外模型中表現出了強烈的抗菌活性,并且部分精油的功效已經在實際應用中得到證實。天然植物精油是環境友好型的天然殺菌劑,是未來新型殺菌劑的重要來源之一,隨著我國各類植物資源的不斷開發研究,必將展現出更為廣闊的發展前景。人們對各類植物精油抑菌機理的研究不斷深入,目前主要體現在3個方面:(1)增強細胞通透性;(2)影響細胞結構及功能;(3)紊亂能量代謝,并且精油不同組分對真菌的抑制有協同作用。精油在抑制真菌產生真菌毒素方面也有著較好的效果,在較低濃度下就可以通過下調關鍵基因的表達降低毒素含量,展現出了較好的應用前景。
我國的植物資源非常豐富,大量的資源待開發,研究更為廣泛的精油以挑選合適的抑菌防霉劑是科學研究及實際應用的需要。同時,為了合理的利用精油資源,提高抗微生物活性及利用率,需要在分子水平上深入研究精油的抑菌機理,如小分子化合物對轉錄翻譯及代謝過程的影響,同時開發新的應用方式也是必不可少的。由于植物精油具有光、熱、pH穩定性差、揮發性強等特點,導致植物精油在實際應用中的利用率低,人們提出了利用各類精油的輸送系統來提升抗微生物活性。精油納米乳液的制備為其提供了很好的思路,這也是下一步研究的方向。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
