干旱脅迫對馬鈴薯生理特性及解剖結構的影響
2021-03-26 08:10:02鄭植尹
中國農學通報 2021年8期
關鍵詞:差異
鄭植尹,王 芳,2,3,4,5
(1青海大學,西寧810016;2青海省農林科學院,西寧810016;3青海大學省部共建三江源生態(tài)與高原農牧業(yè)國家重點實驗室,西寧810016;4青藏高原生物技術教育部重點實驗室,西寧810016;5青海省馬鈴薯育種重點實驗室,西寧810016)
0 引言
馬鈴薯(Solanum tuberosumL.)為茄科(Solanaceae)茄屬(Solanum)的多年生雙子葉草本植物[1]。馬鈴薯是繼小麥、水稻、玉米之后第四大糧食作物,目前也是最大的非谷類食品[2]。馬鈴薯對保障中國糧食安全、促進農業(yè)增效、農民增收、振興農業(yè)經濟方面具有重要意義[3]。目前,干旱是制約馬鈴薯生產的主要問題之一,研究干旱脅迫下馬鈴薯的抗旱反應很有必要,以便為馬鈴薯的抗旱栽培和育種提供理論依據。水是作物生長發(fā)育過程中不可缺少的重要因素,是農業(yè)生產中重要的非生物脅迫,干旱會導致農業(yè)產量下降,持續(xù)的干旱脅迫會使農業(yè)生產顆粒無收[4]。干旱脅迫對作物產量、表型性狀、生理指標、顯微結構均有嚴重影響[5-9]。中國馬鈴薯大多種植在干旱、半干旱地區(qū),干旱是限制馬鈴薯產量的重要逆境因素之一,干旱脅迫嚴重影響著馬鈴薯的正常生長,馬鈴薯受到干旱脅迫時,會抑制其出苗或不出苗,影響馬鈴薯的根部生長,縮短馬鈴薯的無性生長期,進而會影響莖的數量;莖葉的生長受到抑制、根系伸展受到阻礙,結薯時間會明顯延遲,塊莖形成期縮短,從而影響產量,結薯期發(fā)生干旱脅迫會導致單株塊莖數降低[10]。馬鈴薯對干旱脅迫的響應是一個非常復雜的過程,包括滲透調節(jié)作用、抗氧化作用和植物激素調節(jié)作用等生理生化反應等[11-13],進而影響其植物學性狀;葉片顯微結構的變化是植物應對干旱脅迫的重要特征,植物能夠根據水分的虧缺程度及時調整葉片的顯微結構以維持其生存與生長。本文通過研究不同土壤含水量下2個馬鈴薯品種的植物學性狀、生理指標和顯微結構等,初步探討馬鈴薯在干旱脅迫下的抗旱反應和抗旱的生理機制。了解馬鈴薯的干旱適應機理與抗旱反應,了解干旱脅迫下的響應機制,以期為馬鈴薯的抗旱栽培和育種提供理論依據。
1 材料與方法
1.1 試驗材料
試驗材料2個:‘青薯9號’和‘閩薯1號’。2個品種均由青海省農林科學院生物技術研究所提供。
1.2 試驗設計
試驗材料均種植于直徑35 cm、高27 cm的花盆中。試驗采用Hsiao T C[14]提出的關于中生植物水分梯度劃分設計,2個品種各設置5個水平的持續(xù)性水分脅迫處理:(1)T0:土壤相對含水量(即盆栽土壤含水量占最大持水量的百分數)為70%~75%為對照、(2)T1:土壤相對含水量為55%~60%、(3)T2:土壤相對含水量為40%~45%、(4)T3:土壤相對含水量為25%~30%、(5)T4:出苗后不澆水。記錄每一盆出苗時間(取樣時取相同出苗時間的處理),每個處理3次重復。
各處理在播種前將土壤相對含水量保持在70%~75%。出苗后,采用TDR350土壤水分測量儀對土壤含水量進行測定,并按照T1~T4的土壤相對含水量控制,每天上午9:00通過測量并且維持相應的土壤含水量。在出苗后的第 20、35、50、65、80天進行取樣分析。試驗在2020年5—10月期間,青海省馬鈴薯育種重點實驗室進行。
1.3 指標測定
每次取樣時,各品種的各處理取3盆(3次重復)進行農藝性狀測量,包括:株高、莖粗、葉面積、地上部分鮮重和干重、根的鮮重和干重、含水量、匍匐莖數量和長度、薯塊數量和重量等。收獲時根據薯塊產量,計算抗旱系數和抗旱指數。計算公式如式(1)~(2)所示。

葉面積使用Image J軟件測量、組織含水量采用鮮重減干重方法獲得、顯微指標采用徒手切片在OLYMPUS BX51型號生物顯微鏡下觀察并使用Image J軟件測量獲得。
脯氨酸(Pro)測試盒(A107-1-1)、過氧化物酶(POD)測試盒(A084-3-1)、超氧化物歧化酶(SOD)測試盒(A001-1)、丙二醛(MDA)測試盒(A003-1)均購自南京建成。
1.4 數據處理
使用Microsoft Excel 2016進行數據統(tǒng)計分析。
2 結果與分析
2.1 不同土壤含水量對‘青薯9號’和‘閩薯1號’植物學性狀的影響
2.1.1 第一次取樣(出苗20天)形態(tài)學指標的測定 從表1可看出,‘青薯9號’和‘閩薯1號’的莖粗、葉面積、植株鮮重、干重和組織含水量均隨土壤含水量降低而下降;‘青薯9號’在T2處理下株高最大;‘青薯9號’在干旱脅迫下(T1~T4)的分枝數與正常對照(T0)相比極顯著增加。‘閩薯1號’在T1處理下株高值最大;‘閩薯1號’的分枝數在5個處理下均無差異。2品種在株高、莖粗、葉面積、植株干重等性狀上差異顯著,‘青薯9號’的株高、葉面積顯著低于‘閩薯1號’,但植株干重、植株含水量等性狀顯著高于‘閩薯1號’。
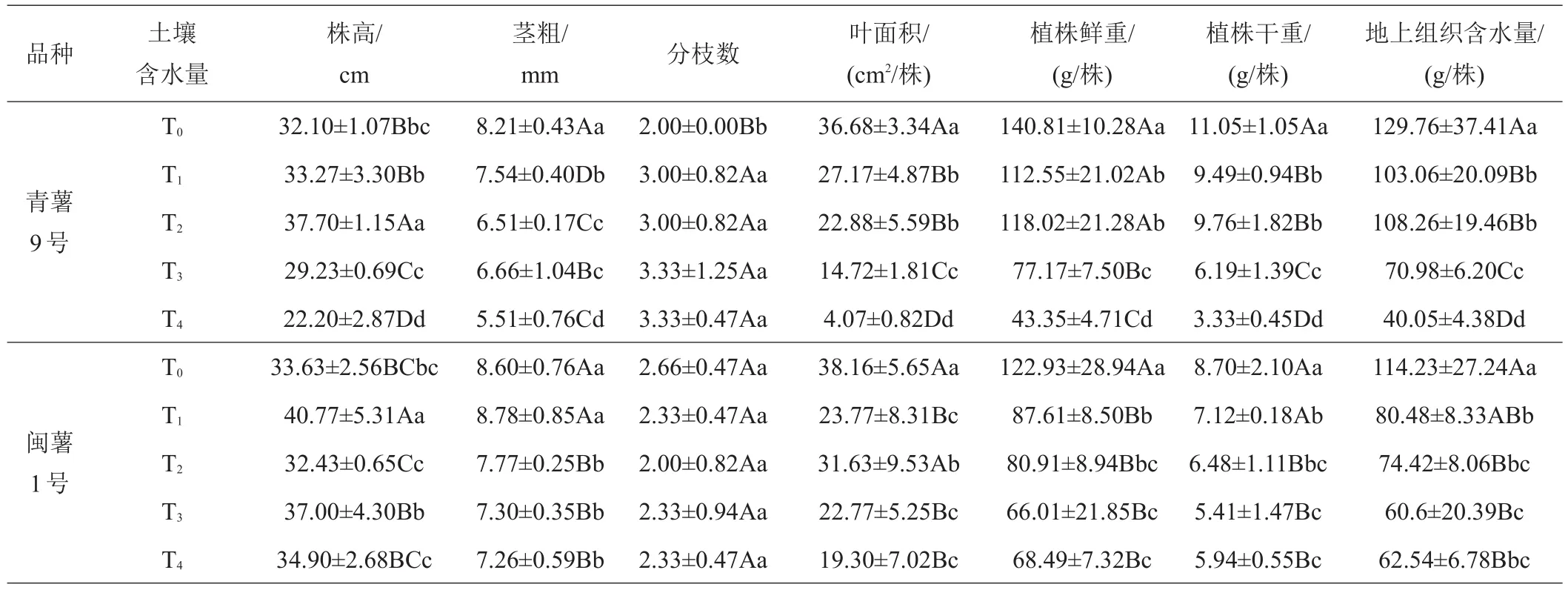
表1 20天時不同土壤含水量下馬鈴薯的植物學性狀
2.1.2 第二次取樣(出苗35天)形態(tài)學指標的差異 35天后第二次取樣,從表2和表3中可以看出,‘青薯9號’和‘閩薯1號’的株高、分枝數、植株鮮重和干重、組織含水量各處理均與對照T0相比,明顯降低,差異極顯著;其他指標如莖粗、葉面積與T0呈差異顯著;2個品種的植株干重在T1和T2處理下也呈極顯著差異,T2處理下降低明顯;隨著干旱脅迫強度增加,‘閩薯1號’的干重下降幅度比‘青薯9號’大,說明干旱脅迫對‘閩薯1號’的影響更大。‘閩薯1號’在各處理下均有薯塊形成,‘青薯9號’只在T0處理下有薯塊形成。2品種在株高、葉面積、分地上部分鮮重和干重、植株含水量、匍匐莖長度、根長、根鮮重和干重等性狀上差異顯著,‘青薯9號’的葉面積顯著低于‘閩薯1號’,但株高、地上部分鮮重和干重、植株含水量、匍匐莖長度、根長、根鮮重和干重等性狀顯著高于‘閩薯1號’,兩品種在匍匐莖數量上沒顯著差異。

表2 35天時不同土壤含水量下馬鈴薯地上部分的植物學性狀
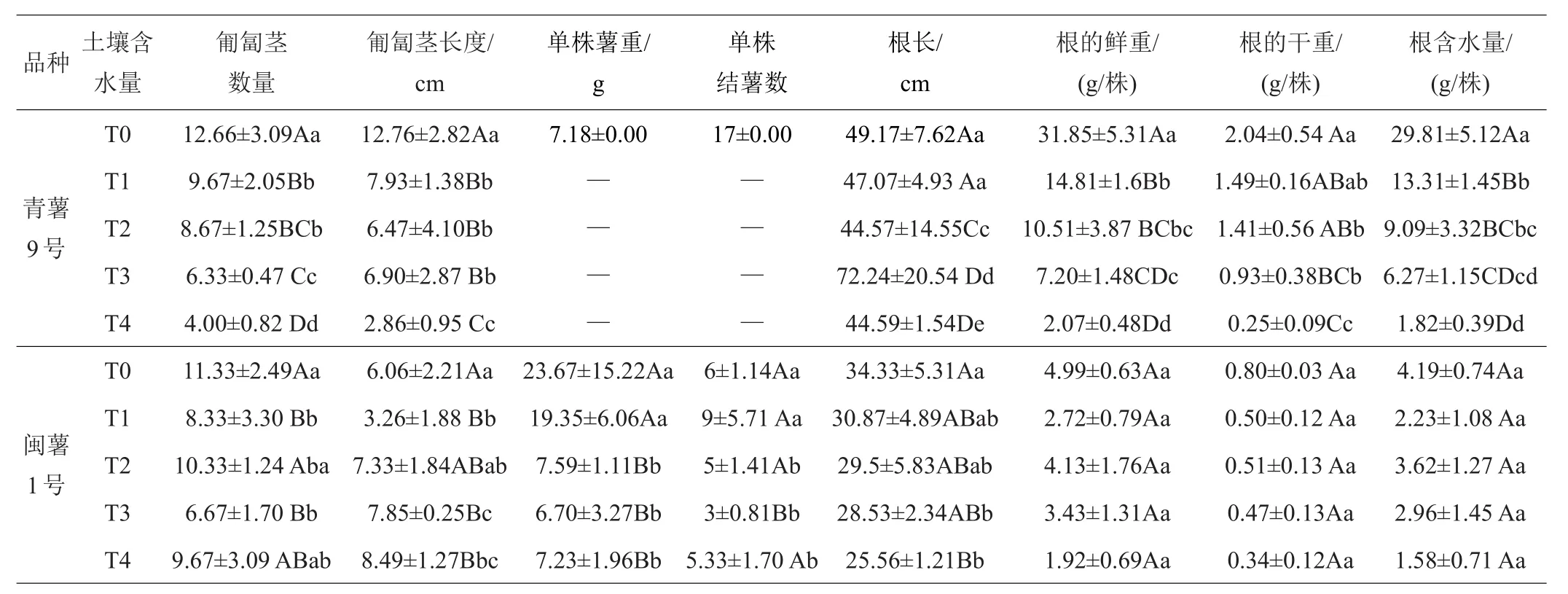
表3 35天時不同土壤含水量下馬鈴薯地下部分的植物學性狀
2.1.3 第三次取樣(出苗50天)形態(tài)學指標的差異 50天時,‘青薯9號’的地上部植株干重比35天時增加了73.69%。‘閩薯1號’的地上部植株干重比35天時增加了61.28%。‘閩薯1號’結薯的數量與35天時相比沒有明顯增加,但各處理下單株薯重均明顯增加,說明‘閩薯1號’已進入薯塊膨大期。2品種在株高、莖粗、分枝數、葉面積、地上部分鮮重和干重、植株含水量、匍匐莖長度、根長、根鮮重和干重等性狀上差異顯著,‘青薯9號’的葉面積還是顯著低于‘閩薯1號’,但株高、莖粗、分枝數、地上部分鮮重和干重、植株含水量、匍匐莖長度、根長、根鮮重和干重等性狀顯著高于‘閩薯1號’。
2.1.4 第4次取樣(出苗65天)形態(tài)學指標的差異 2個品種在65天時地上部分干重與50天時的干重相比,增加緩慢。65天時,‘青薯9號’的匍匐莖的數量在T0~T2處理下高于‘閩薯1號’,匍匐莖長度在各處理下均高于‘閩薯1號’,并開始有少量結薯。‘閩薯1號’薯塊在T0處理下比50天時顯著增加,T1~T3處理增加緩慢,T4處理沒有增加,而且與對照T0相比,單株薯重隨著土壤含水量的下降而下降,且達到極顯著差異,可見,這一時期是‘閩薯1號’薯塊膨大期,水份脅迫對馬鈴薯的生長產生了較大影響。2品種相比,‘青薯9號’的葉面積、單株薯重和數量顯著低于‘閩薯1號’,其他性狀上差異仍然顯著且都是‘青薯9號’高于‘閩薯1號’,特別是根系的鮮重和干重極顯著高于‘閩薯1號’。

表4 50天時不同土壤含水量下馬鈴薯地上部分的植物學性狀
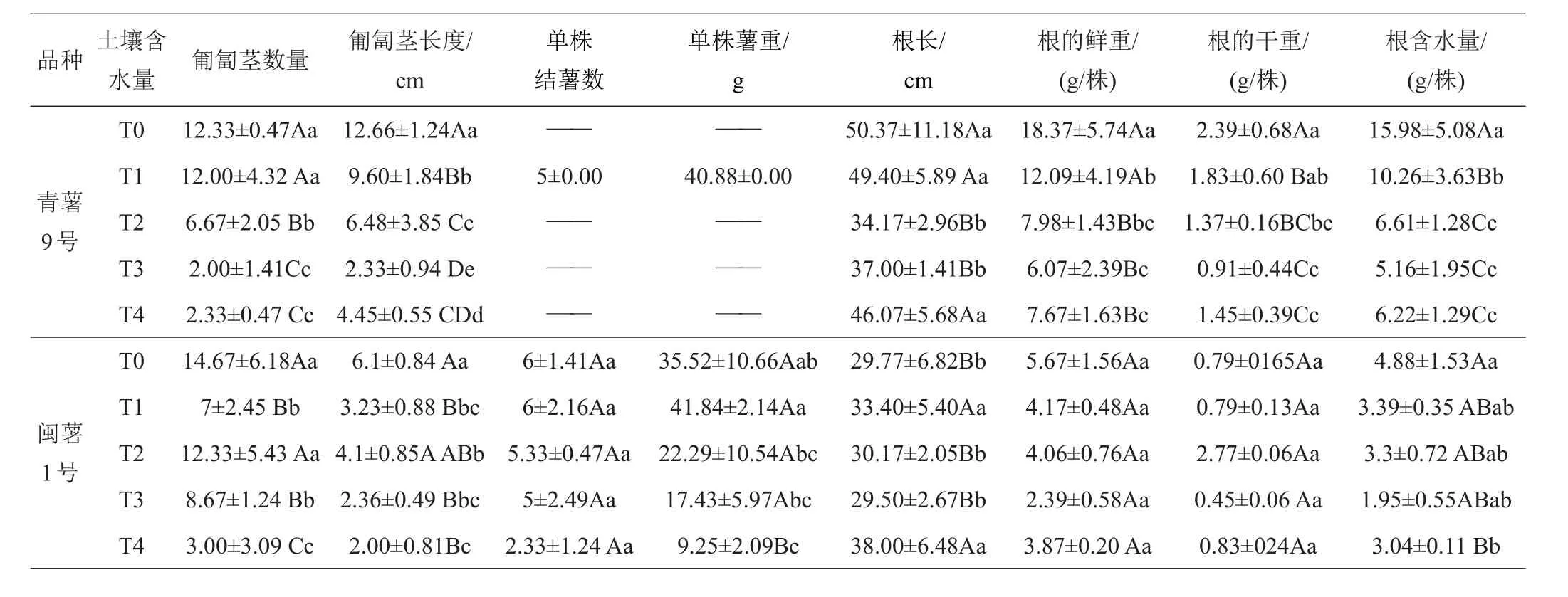
表5 50天時不同土壤含水量下馬鈴薯地下部分的植物學性狀

表6 65天時不同土壤含水量下馬鈴薯地上部分的植物學性狀
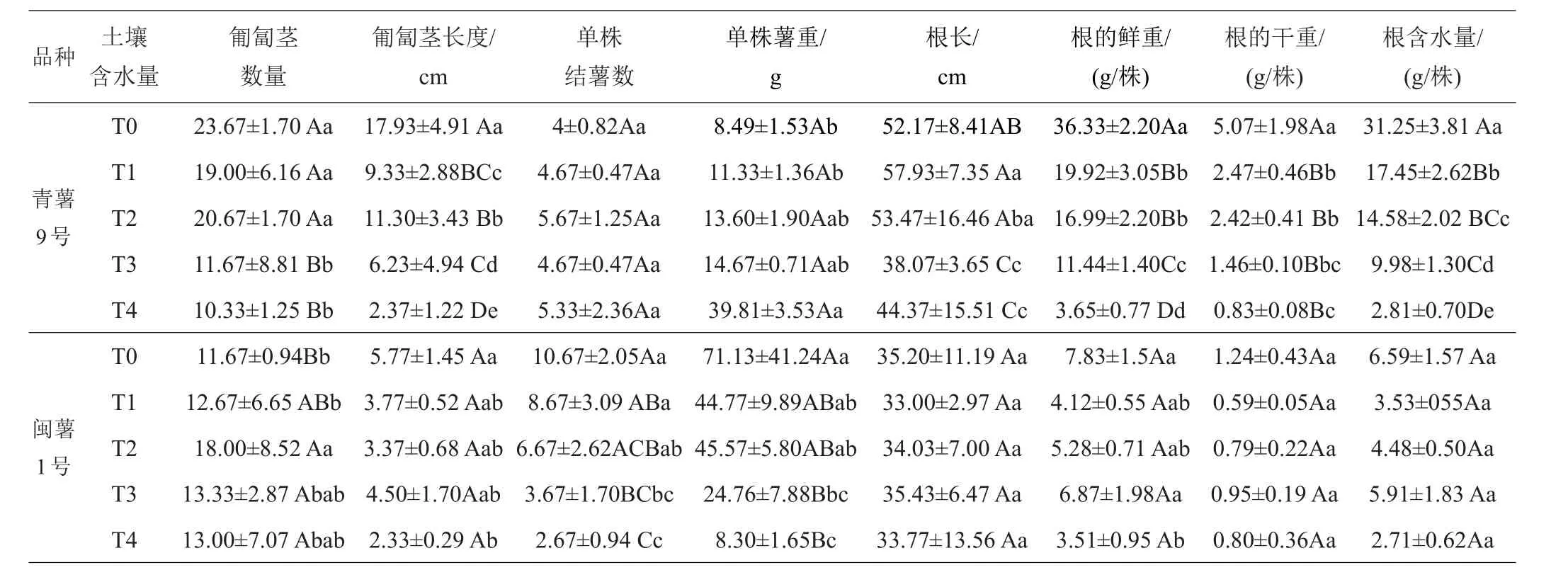
表7 65天時不同土壤含水量下馬鈴薯地下部分的植物學性狀
2.1.5 第4次取樣(出苗80天)形態(tài)學指標的差異 由于T4處理為出苗后不再澆水,2個品種在80天時T4處理因干旱致死,所以沒有調查數據。80天時2品種的地上部分鮮重、地上部分干重沒有增加,‘青薯9號’的各處理下地上部干重還略有降低,植株含水量各處理均略有降低;2品種根的重量增加趨勢不明顯,只有薯塊的數量和重量增加顯著,2品種的結薯數在4個處理下差異不顯著,為6~8個;‘青薯9號’的薯塊重量增加明顯,尤其在T0處理下,每株薯塊平均重量為93.70 g,2個品種在干旱脅迫的T1~T3處理下,每薯塊平均重量均極顯著低于對照T0。2品種相比,‘青薯9號’的葉面積還是顯著低于‘閩薯1號’,2品種在株高、植株鮮重和干重、根長、根鮮重和干重、含水量性狀上差異仍然顯著且都是‘青薯9號’高于‘閩薯1號’。
2.1.6 收獲期2品種的產量調查‘閩薯1號’是早熟品種,‘青薯9號’屬于晚熟品種,因此這2個品種分別在出苗后90天和135天全部收獲。在T0、T1、T2處理下,‘青薯9號’總薯重均高于‘閩薯1號’,2品種的抗旱系數和抗旱指數在T1、T2處理下,‘青薯9號’均高于‘閩薯1號’。
2.2 不同土壤含水量對馬鈴薯生理指標的影響
由圖1~4可以看出,干旱脅迫對葉片中脯氨酸含量有極顯著影響(P<0.01),在T0~T4處理下,脯氨酸含量有隨著土壤含水量的降低而增加的趨勢,且隨干旱脅迫時間的延長,各處理均有降低趨勢,平均含量在20天時達到最大,50天時最小。丙二醛在20天時含量隨著土壤含水量的降低而降低,且達到了顯著差異,在35天和50天時,丙二醛的含量卻隨著土壤含水量的降低而升高,在50天T4時達到最大‘,青薯9號’為1.619 mg port/mL,‘閩薯1號’為1.833 mg port/mL,并且在50天時均達到了極顯著差異。2個品種葉片中過氧化物酶的含量在20天時從T0~T4均為下降趨勢,在T2與T4時達到了極顯著差異;在35天時2品種的過氧化物酶含量最低且在5個處理間差異不大;在50天時2品種葉片中的過氧化物酶含量卻隨著土壤含水量的降低而升高,且兩個品種在50天T4時達到最大,‘青薯9號’和‘閩薯1號’分別為5.44 U/mg port和7.63 U/mg port,差異顯著。‘青薯9號’在20天與35天時葉片中超氧化物歧化酶的含量均有隨著土壤含水量的降低有降低的趨勢,且均與對照達到極顯著差異;但‘閩薯1號’在20天時T2處理最高,35天時含量隨著土壤含水量的降低而降低,50天時含量隨著土壤含水量的降低而升高。
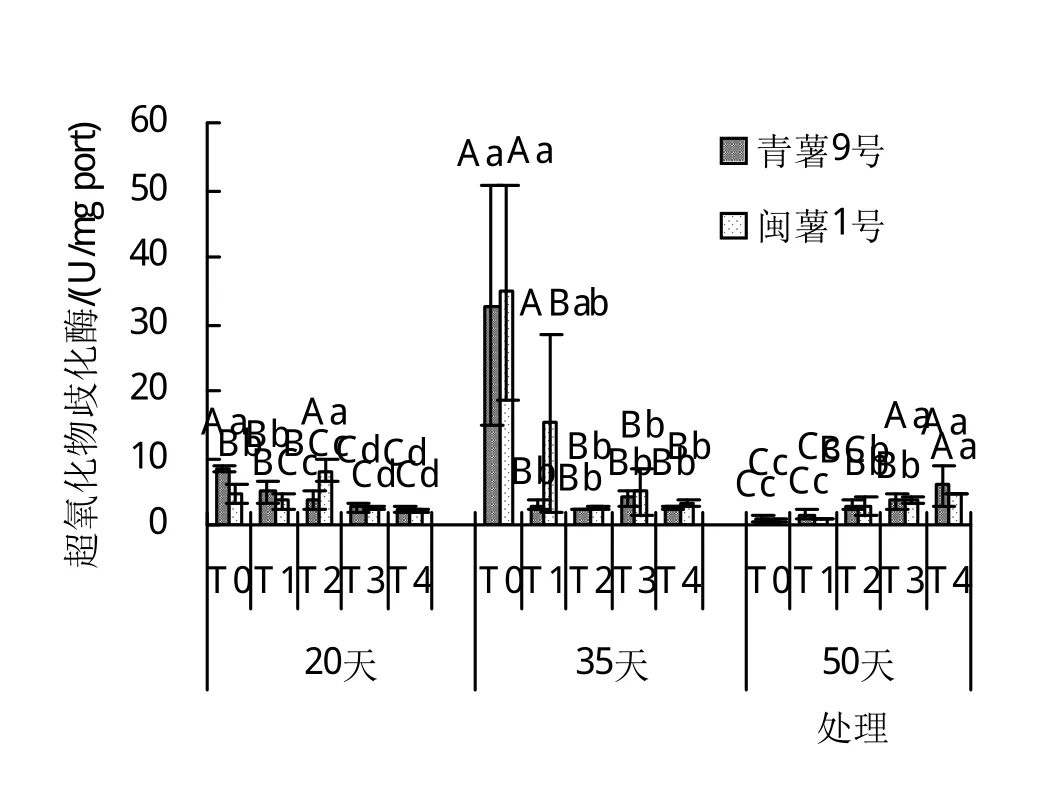
圖1 20、35、50天時不同土壤含水量超氧化物歧化酶(SOD)的含量圖
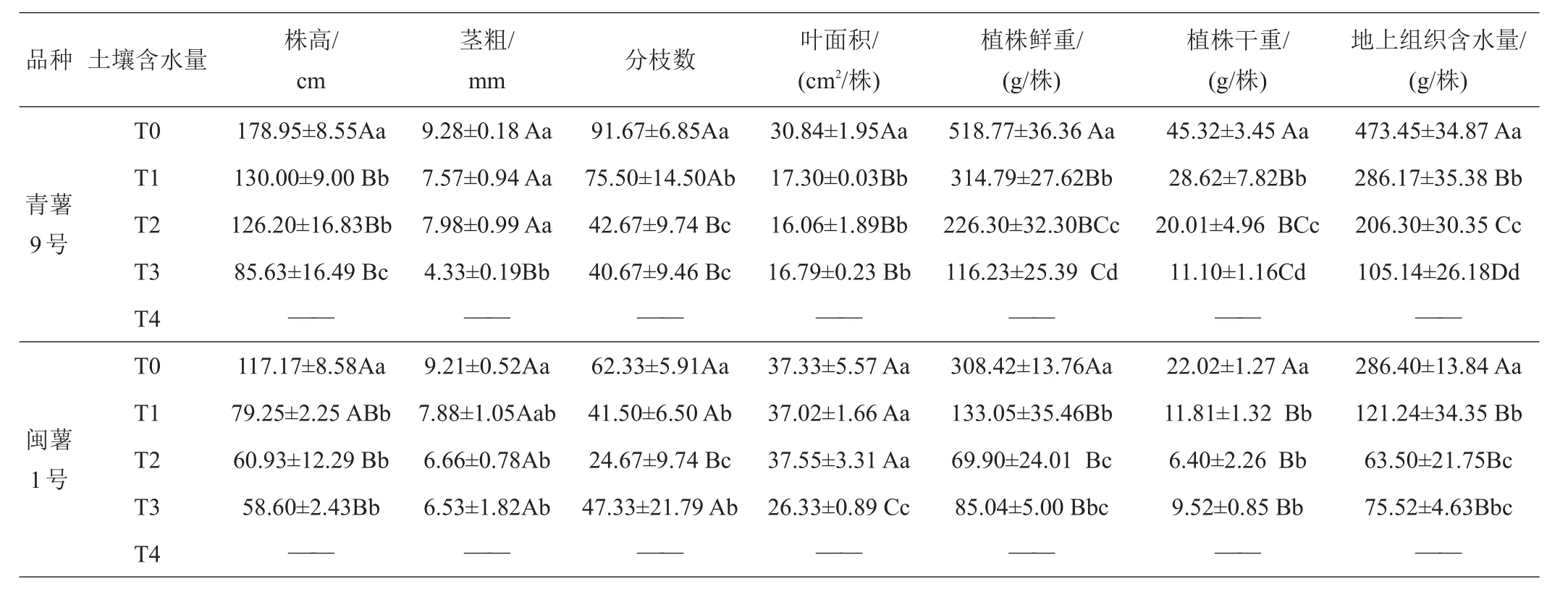
表8 80天時不同土壤含水量下馬鈴薯地上部分的植物學性狀
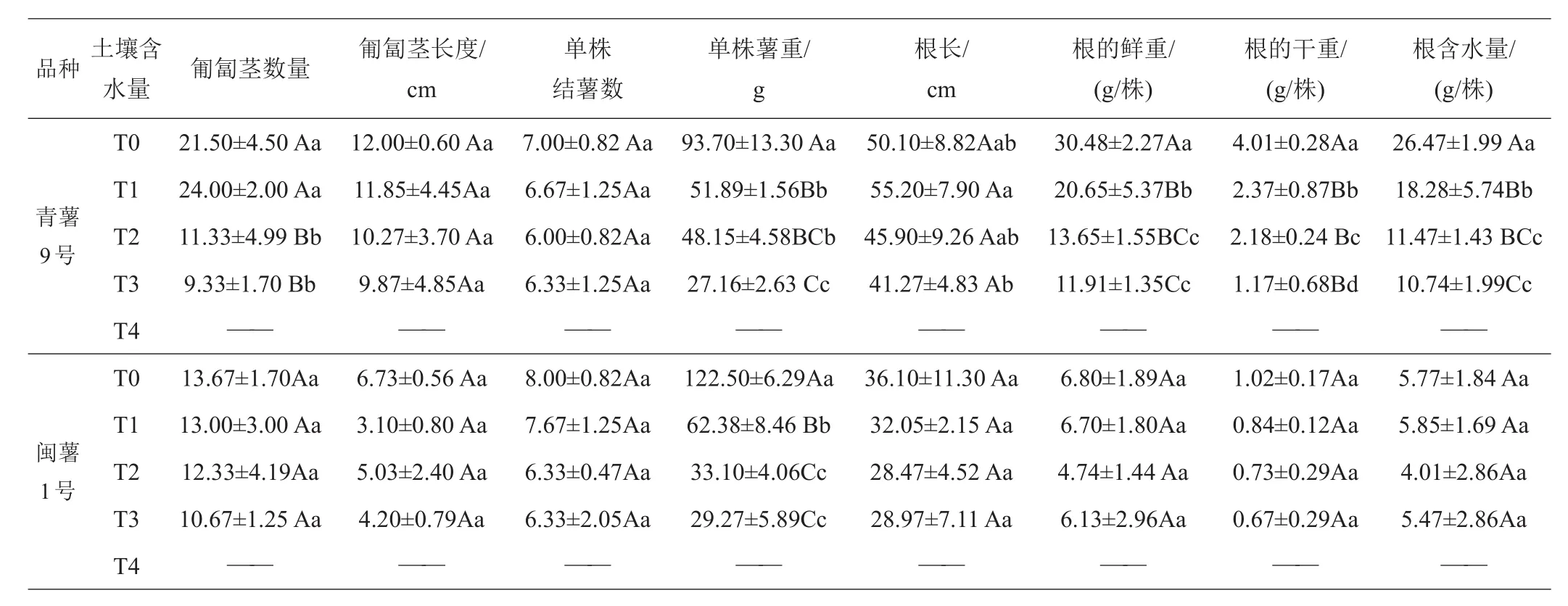
表9 80天時不同土壤含水量下馬鈴薯地下部分的植物學性狀
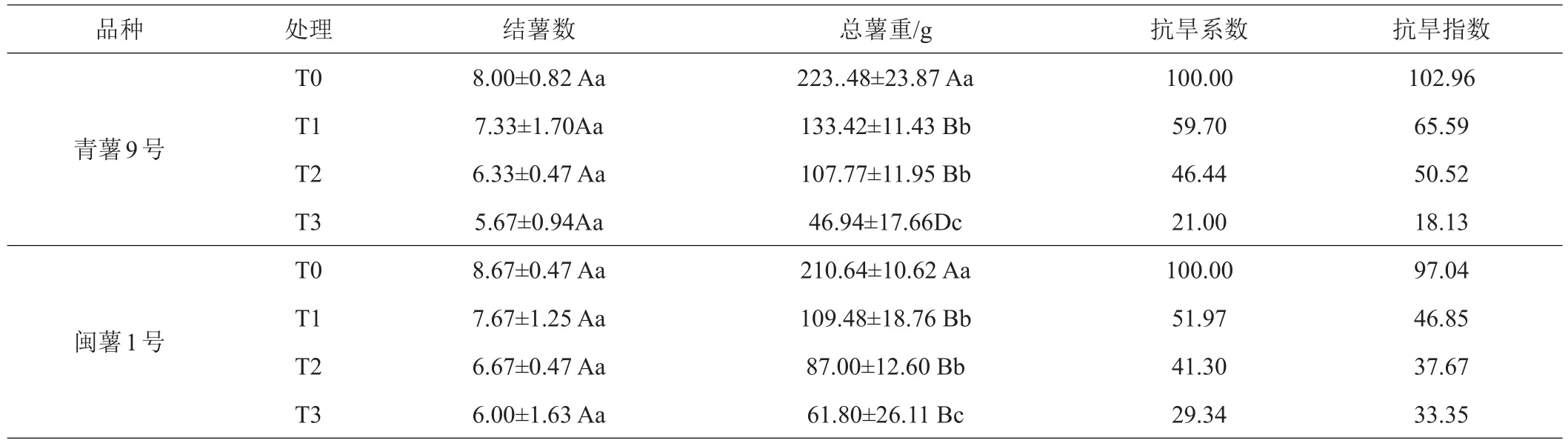
表10 收獲期馬鈴薯產量及抗旱性
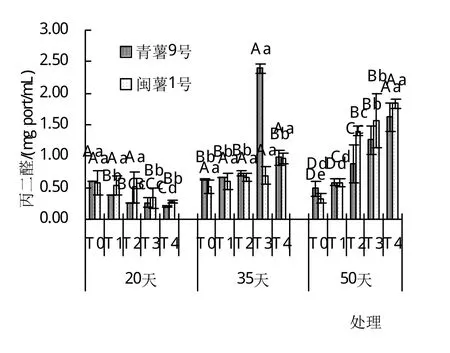
圖2 20、35、50天時不同土壤含水量丙二醛(MDA)的含量圖
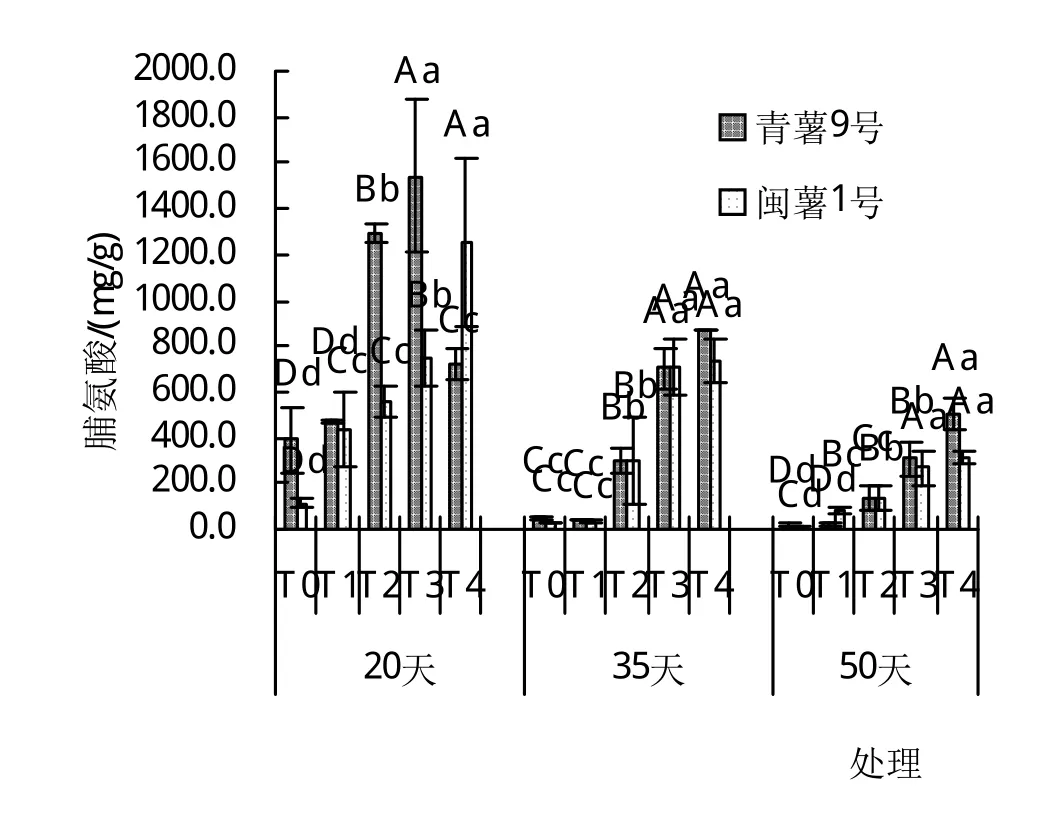
圖3 20、35、50天時不同土壤含水量馬鈴薯脯氨酸(Pro)的含量
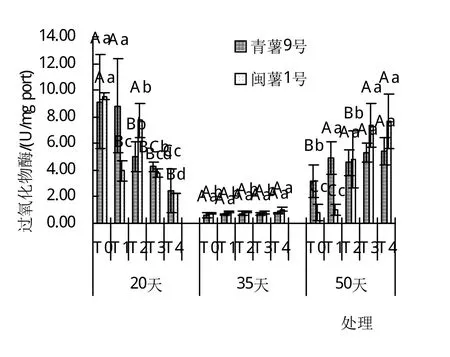
圖4 20、35、50天時不同土壤含水量過氧化物酶(POD)的含量
2品種相比,脯氨酸(Pro)含量在20天時,兩品種間差異顯著,‘青薯9號’顯著高于‘閩薯1號’;但在35天和50天時,‘青薯9號’的脯氨酸含量仍然高于‘閩薯1號’,但差異不顯著。2品種葉片中的丙二醛(MDA)含量在20天和50天時,閩薯1號均高于‘青薯9號’;兩品種超氧化物歧化酶(SOD)無顯著差異,過氧化物酶(POD)的含量在20天和50天時的T1處理下‘青薯9號’顯著高于‘閩薯1號’,其余處理差異不顯著。
2.3 不同土壤含水量對馬鈴薯顯微指標的影響
圖5為5次取樣葉片的解剖結構,表11為2個品種葉片的顯微結構調查結果。可看出,2品種的上下表皮厚度在20天時較薄,35天時明顯增加,隨后不再顯著增加。在不同的土壤含水量處理下‘,青薯9號’的上表皮厚度在20、35、65天時5個處理間無顯著差異,50天和80天時差異顯著但沒有規(guī)律。下表皮厚度在20天與35天時有隨土壤含水量減少而減小的趨勢,且20天時達到顯著差異,最高值為T0處理下的2.97μm,最低值為T3處理下的2.14μm;35天到65天時各處理的下表皮厚度無顯著差異,在80天時下表皮厚度差異達到顯著水平(P<0.05),但無明顯規(guī)律,最大值為T3處理的6.04μm,最小值為T2處理的3.85μm。‘閩薯1號’的上表皮厚度與下表皮厚度在20天到80天時間內均有著先增加后減小的趨勢,且在20、35、50天和65天差異達到顯著水平(P<0.05),但隨土壤水分脅迫加重變化規(guī)律不明顯。

圖5 5次葉片取樣(20~80天)解剖結構

續(xù)表11
2個品種海綿組織厚度在20天時隨土壤水分脅迫增加均有先減小后增加的趨勢,在35~65天期間均有先增加后減小的趨勢,且均達到顯著水平(P<0.05);2個品種的柵欄組織厚度變化規(guī)律不明顯,但2品種相比,20~35天期間,‘青薯9號’的柵欄組織厚度顯著高于‘閩薯1號’,在50天以后兩者差異不顯著。在20天時,2個品種的柵海比均有著隨著土壤含水量的減少而減小的趨勢,且均與對照達到極顯著差異,在35~80天的處理中,隨著土壤含水量的減少柵海比有先增加后減小的趨勢,且在50天和65天時達到顯著水平(P<0.05)。
把葉片的上表皮、下表皮、柵欄組織、海綿組織厚度相加,得到葉片的厚度。可以看出,在20~50天期間,‘青薯9號’的葉片厚度顯著高于‘閩薯1號’,65~80天期間,2品種差異不顯著,這期間葉片的厚度隨著土壤水分脅迫加重呈增加趨勢,處理間達極顯著差異。
3 結論
水分脅迫對2個馬鈴薯品種表型性狀(鮮重、莖粗、地上部分鮮重、地上部分干重、葉面積、根的鮮重、根的干重和根長、匍匐莖數量與長度)均有顯著影響,且隨著土壤含水量的降低而降低。‘青薯9號’的葉面積始終小于‘閩薯1號’,但‘青薯9號’的葉片厚度、分枝數、植株鮮重、干重、匍匐莖長度等性狀均大于‘閩薯1號’,特別是‘青薯9號’的根長、根重與‘閩薯1號’有顯著差異,這些性狀使‘青薯9號’有較好的抵抗干旱脅迫能力‘;青薯9號’的產量高于‘閩薯1號’,其抗旱系數和抗旱指數均高,也說明‘青薯9號’抗旱性好。
生理指標分析表明,水分脅迫下,馬鈴薯的脯氨酸含量、丙二醛和超氧化物歧化酶與對照相比有極顯著差異(P<0.01)。2個品種的脯氨酸含量都會隨著土壤含水量的降低而升高‘,青薯9號’的脯氨酸含量顯著高于‘閩薯1號’,增加了細胞和組織的保水能力,提高其抗旱性能。在出苗20天時2個品種丙二醛含量隨著土壤含水量的降低而降低,在35天和50天時2個品種的丙二醛含量隨著土壤含水量的下降而升高,說明持續(xù)的干旱脅迫對馬鈴薯苗已造成傷害;2品種葉片中的丙二醛(MDA)含量在20天和50天時‘,閩薯1號’均高于‘青薯9號’。超氧化物歧化酶則是在出苗20天和35天時隨著土壤含水量的下降而下降,在50天時隨著土壤含水量的下降而升高。水分脅迫對馬鈴薯和過氧化物酶的含量影響顯著(P<0.05),2品種過氧化物酶含量在出苗20天時隨著土壤含水量的下降而下降,但是在35天和50天時2個品種的過氧化物酶含量會隨著土壤含水量的下降而升高。
2個馬鈴薯品種隨著土壤含水量的降低葉片細胞排列變得混亂,但可看出:在20~50天期間‘,青薯9號’的葉片厚度顯著高于‘閩薯1號’,65~80天期間,2品種差異不顯著,這期間葉片的厚度隨著土壤水分脅迫加重呈增加趨勢,說明馬鈴薯在受到水分脅迫后,通過改變柵欄組織和海綿組織的厚度來適應水分脅迫的影響。
4 討論
目前干旱問題日趨嚴重,已經對馬鈴薯的生產造成了顯著影響,限制了馬鈴薯產量的進一步提高。本研究認為,隨著土壤水分脅迫的加重,致使‘青薯9號’和‘閩薯1號’的株高、莖粗、葉面積、植株鮮重、植株干重和組織含水量明顯降低。脯氨酸(PRO)是重要的滲透調節(jié)物質,正常生長時植物中的游離脯氨酸微乎其微,當植物受到干旱脅迫時,脯氨酸會大量增加,且耐旱性越強脯氨酸含量越高[16]。本研究表明,‘青薯9號’和‘閩薯1號’的脯氨酸隨著干旱梯度的增加而急劇上升,與白建芳[17]在玉米和張清海[18]在小麥中的結果一致。丁玉梅等[19]對馬鈴薯進行干旱處理后,其脯氨酸含量比正常灌溉時明顯升高,并且干旱時脯氨酸相對含量越高,品種產量的抗旱系數越大,抗旱性越強。劉學師等[20]的研究表明,在干旱脅迫下,脯氨酸是植物體內理想的滲透調節(jié)物質,使植物適應干旱逆境,防止細胞和組織脫水;另外,脯氨酸能增強生物聚合體的親水性,在可利用水減少的情況下,大量的脯氨酸積累能提高原生質膠體的穩(wěn)定性,是穩(wěn)定植物體內物質代謝的決定因素。
有研究表明[21],馬鈴薯的產量與分枝數呈正相關,本研究中,在干旱脅迫下,‘青薯9號’的分枝數增長數比‘閩薯1號’多,說明在干旱脅迫下,‘青薯9號’比‘閩薯1號’的產量潛力高,抗旱性好。
植物抗旱性與植物形態(tài)解剖結構有一定的相關性,葉片在植物的各器官中對于干旱脅迫較為敏感,其解剖結構通常用來反映植物的抗旱性。扁桃葉片在干旱脅迫下葉片結構會朝著減少葉片蒸騰、增強葉片細胞儲水能力的方向改變,以適應干旱脅迫逆境條件[22]。本文通過切片試驗結果顯示,在干旱脅迫下,‘青薯9號’的柵欄組織厚度有著隨著土壤含水量的降低先減少后增加的趨勢,可能是由于馬鈴薯柵欄組織在剛受到脅迫時,干旱會導致柵欄組織厚度有短時間的下降,之后馬鈴薯通過增加葉片柵欄組織的厚度的方式提高光合作用來緩解干旱導致的葉片光合速率下降。柵海比值變化規(guī)律不明顯,這與白刺花[23]和抗旱性強的花生品種[19]在干旱脅迫下柵欄組織和海綿組織的變化規(guī)律相同,但與艾納香[24]幼苗的研究結果相反,艾納香生長可承受的最大范圍為中度脅迫,艾納香幼苗長時間受到重度干旱脅迫后,柵欄組織減少,海綿組織增加,柵海比明顯降低[25]。馬鈴薯的柵海比值變化呈顯著差異,可能說明馬鈴薯與白刺花和花生一樣,具有較強的干旱耐受性,而艾納香對干旱的耐受性為中等有關。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
