果糖在食欲調控中的作用機制
2021-03-26 03:01:00甘凱文于浩泳
醫學綜述 2021年5期
甘凱文,于浩泳
(上海交通大學附屬第六人民醫院內分泌代謝科,上海 200233)
果糖是一種單糖,為葡萄糖的同分異構體,主要以游離形態存在于水果和蜂蜜中。1970年果糖的生產技術取得突破,自此果糖以果葡糖漿為主要形式被大量生產,并廣泛用于食品加工行業[1-2]。研究顯示,美國人均果糖攝入量從1970年末的37 g/d增加至1999—2004年的49 g/d[1]。近年來果糖消費量和攝入量劇增,每日人均攝入糖量遠超世界衛生組織指南的推薦量[3],全球范圍內肥胖的發生率與果糖攝入量呈正相關[2]。研究表明,攝入果糖可導致食欲亢進,引起2型糖尿病、胰島素抵抗、高脂血癥以及神經損傷等[4-6],與單純添加葡萄糖的等熱量飲食相比,果糖更易導致肥胖和內臟脂肪蓄積,降低飽腹感,從而導致更嚴重的代謝紊亂[7]。
中樞神經系統(central nervous system,CNS)和外周組織產生的食欲相關信號聯合作用調控食欲。在CNS中,下丘腦起主導作用,與弓狀核、腹內側核、背中核、室旁核、外側下丘腦等共同參與食欲信號的合成、釋放及調控不同區域的食欲信號[8];外周的食欲信號通路始于受攝食刺激分泌的胃腸道激素,然后通過不同神經通路傳入中樞,并通過中樞調節攝食行為[9]。現就果糖在食欲調控中的作用機制進行綜述。
1 果糖對中樞食欲信號的調控
1.1果糖與下丘腦食欲調節神經肽 下丘腦作為攝食中樞在能量代謝調控起主導作用,可合成食欲調節相關的神經肽,包括促食欲神經肽[神經肽Y、刺鼠基因相關蛋白(agouti-related protein,AgRP)、黑色素濃集激素、增食欲素]和抑制食欲的神經肽[阿片黑素促皮質素原(proopiomelanocortin,POMC)、可卡因-苯丙胺調節轉錄肽][5,8]。研究發現,接受高果糖飲食或腦室內注射果糖溶液的小鼠進食量和體重均有所增加,并觀察到POMC信使RNA(messenger RNA,mRNA)的表達水平下降[10-12]。關于果糖對神經肽Y和AgRP表達的影響存在爭議,可能與各研究中果糖劑量及觀察時長不同有關。長期僅給予果糖的間歇性攝食大鼠模型中觀察到伏隔核內增食欲素神經元活性下降和外側核增食欲素神經元活性增強,增食欲素神經元激活機制可能與果糖攝入引起外周循環三酰甘油水平顯著升高相關[13],表明長期果糖攝入可降低伏隔核中γ-羥基丁酸能神經元對外側下丘腦和腹側蒼白球的輸出,激活外側下丘腦內增食欲素神經元釋放增食欲素-A觸發攝食效應,導致果糖攝入持續增加,形成正反饋循環[12]。
1.2與果糖代謝途徑相關的AMP活化蛋白激酶(AMP-activated protein kinase,AMPK)/丙二酰輔酶A(malonyl-coenzyme A,MCoA)信號通路 果糖通過與腸黏膜上皮的葡萄糖轉運體結合進入血液循環,并運輸至肝臟進行代謝,葡萄糖轉運體5是果糖唯一的特異性轉運體,不受胰島素影響,主要存在于小腸、肌肉組織、海馬體、大腦皮質小膠質細胞以及小腦浦肯野細胞中[14-15]。研究指出,高果糖飲食喂養小鼠的小腸和中樞葡萄糖轉運體5 mRNA的表達上調,影響大腦能量代謝穩態[16-17]。雖然葡萄糖和果糖均通過糖酵解途徑代謝,但在肝臟代謝的起始步驟不同,果糖繞過糖酵解中的限速步驟自主代謝,由不依賴胰島素的特異性果糖激酶催化果糖磷酸化生成1-磷酸果糖。在中樞果糖繞過限速步驟會引起ATP的迅速消耗及AMP的代償性增多,造成下丘腦 AMP/ATP比值升高,進而激活AMPK使乙酰輔酶A羧化酶磷酸化失活,減少其催化產物MCoA的生成[6]。AMPK的激活受激素水平和營養狀態的調節,如胃饑餓素和低血糖,中樞AMPK激活刺激攝食,而瘦素、胰島素和葡萄糖則抑制攝食[18-19]。MCoA是下丘腦信號級聯中調節高等動物能量平衡的關鍵中間體,通過下丘腦食欲調節神經肽系統調控食欲信號和攝食行為[6]。MCoA的升高可激活弓狀核神經元,降低神經肽Y和AgRP的表達,并上調α-促黑素(POMC的蛋白水解產物)和可卡因-苯丙胺調節轉錄肽的表達,最終導致食欲及進食量下降[10]。相反,果糖可導致下丘腦MCoA水平下降[6,18],食欲亢進、攝食增加及肥胖[6](圖1)。果糖對血腦屏障通透性的作用尚未明確,曾有學者質疑膳食果糖直接進入CNS影響食欲的學說,但研究證實小鼠大腦的多個區域存在果糖代謝所需的酶類及果糖轉運蛋白,同時葡萄糖轉運體5也在血腦屏障中表達[20]。其他研究表明,外周靜脈注射果糖后下丘腦細胞外液中果糖水平增加,支持果糖直接經血腦屏障進入CNS進行代謝,而不需經肝臟轉化為葡萄糖后再進入CNS代謝的結論[16,21]。
1.3果糖與內源性大麻素系統(endocannabinoid system,ECS) 生物體自身可產生大麻素,稱為內源性大麻素(endocannabinoid,EC),是一類神經遞質分子,主要包括N-花生四烯酸氨基乙醇和2-花生四烯酸甘油[22]。N-花生四烯酸氨基乙醇主要是在N-酰基磷脂酰乙醇胺磷脂酶D和磷脂酶C的催化下合成,二酰基甘油脂肪酶α和β主要參與2-花生四烯酸甘油的合成,可分別被脂肪酸酰胺水解酶和單酰基甘油酯酶降解滅活,EC、與EC結合的受體以及其合成和降解酶類共同組成ECS[23]。大麻素具有促進食欲的作用。多項研究表明,ECS在食欲調節、脂肪合成、胰島素抵抗、能量代謝及攝食等方面發揮重要作用[23-24]。EC通過與CNS突觸前神經末梢表達的大麻素受體1(cannabinoid 1 receptor,CB1R)結合促進食欲[4,24-25]。目前已證實選擇性CB1R拮抗劑利莫那班可通過作用于CNS及外周組織的CB1R達到有效降低食欲和減重的目的,特別是降低對甜食、飲料及可口食物的攝食欲望,而CB1R激動劑則起相反作用[4,26]。研究表明,果糖喂養的小鼠下丘腦CB1R mRNA水平顯著升高,而葡萄糖喂養的小鼠則未觀察到明顯的CB1R mRNA水平上升[4,25]。另一項研究指出,果糖的攝入可影響EC的合成和降解酶系統,在果糖喂養的小鼠中可觀察到脂肪酸酰胺水解酶、N-酰基磷脂酰乙醇胺磷脂酶D、二酰基甘油脂肪酶β mRNA表達升高,而磷脂酶C、單酰基甘油酯酶、二酰基甘油脂肪酶α mRNA表達下降,提示果糖較其他糖類(蔗糖和葡萄糖)對ECS的影響更大[4],但該研究并未對相關基因及EC蛋白的表達水平進行分析,僅根據EC相關酶系統的基因轉錄水平變化難以直接評價果糖對ECS的實際影響。此外,ECS除直接通過中樞效應調節食欲,還可通過與腦內調節食欲的相關通路(如獎賞回路、5-羥色胺系統、增食欲素-A信號通路、內源性阿片肽)相互聯系,共同發揮調節食欲和攝食行為的功能[24,27]。
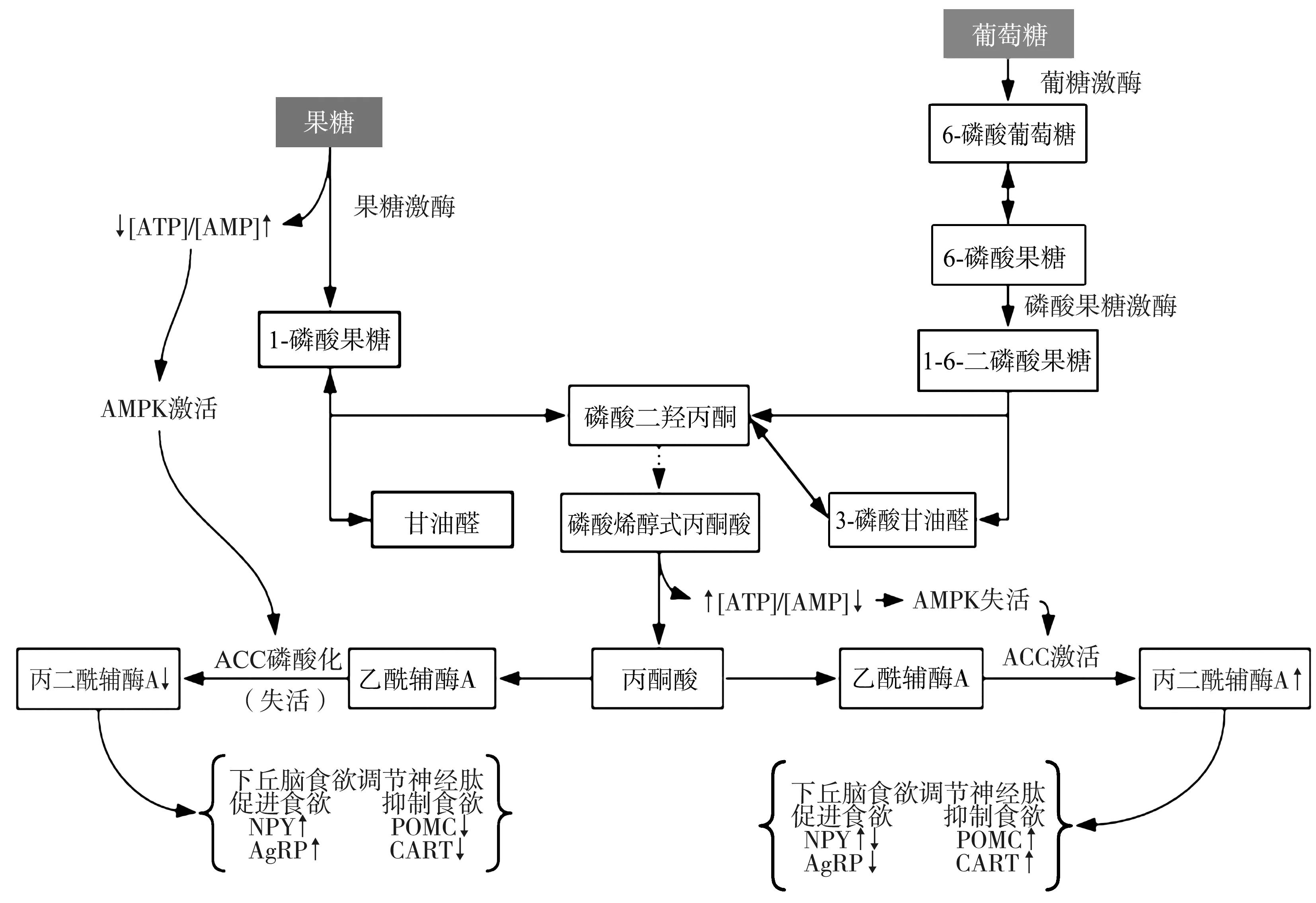
ATP:腺苷三磷酸;AMP:腺苷一磷酸;AMPK:AMP活化蛋白激酶;ACC:乙酰輔酶A羧化酶;NPY:神經肽Y;AgRP:刺鼠基因相關蛋白;POMC:阿片黑素促皮質素原;CART:可卡因-苯丙胺調節轉錄肽;MCoA:丙二酰輔酶A
1.4果糖對大腦區域活動的影響 下丘腦為食欲調節中樞,其與參與調節進食動機及獎賞系統的大腦其他區域(包括紋狀體、眼窩前額皮質、杏仁體和腦島)共同組成綜合網絡調控食欲[8]。有研究采用功能性磁共振成像技術以局部腦血流量作為神經元激活的間接指標,探討大腦區域活動與果糖介導的攝食行為改變的關聯[21,28]。正常生理狀態下,機體從饑餓狀態轉為飽腹狀態時,下丘腦及紋狀體活動減弱,但攝入果糖后,人類丘腦、海馬、后扣帶皮質、梭狀回和視覺皮質的腦血流量顯著減少,但并未降低下丘腦活動,反而引起下丘腦活動短暫且小幅度的升高,同時下丘腦與紋狀體功能連接未見增強[21]。此外,海馬體的記憶過程也參與了進食行為的調節,海馬體損傷的嚙齒動物可失去識別饑餓和飽足狀態的能力,導致食欲亢進[29]。人類海馬體的激活與食物渴求、饑餓狀態等有關,提示攝入果糖后海馬體活動減少可能誘導食欲亢進的發生[21]。另一方面,果糖與享樂攝食及獎賞機制有關,相對于葡萄糖,果糖攝入導致大腦視覺皮質和左眼眶額葉皮質對食物線索的反應性增強,表明果糖增強了對食物線索的獎賞和動機信號的傳遞,并刺激大腦中負責處理注意力和獎賞的區域,最終表現為對食物強烈的渴望和饑餓感[30]。
2 果糖對外周食欲信號的影響
攝食行為刺激胃腸分泌細胞分泌約20種多肽激素,部分激素與CNS相互作用組成腦-腸軸調控攝食行為及能量穩態,這些胃腸激素被稱為腦腸肽,分為促食欲因子(胃饑餓素)和飽腹因子[瘦素、胰高血糖素樣肽1(glucagon-like peptide-1,GLP-1)]兩類,除腦腸肽外,胰島素也在人體代謝和飲食行為中起重要作用。
2.1胃饑餓素 胃饑餓素由胃腸的內分泌細胞和大腦弓狀核分泌,是唯一在外周刺激食欲的胃腸激素,與短期饑餓、食物攝入和體重有關,與血糖、胰島素及瘦素水平呈負相關[8,31]。胃饑餓素調節進食和能量穩態依賴于其與生長激素促分泌素受體的結合,生長激素促分泌素受體廣泛分布于脂肪組織、腎上腺、生殖腺、胰腺、胃以及CNS(如下丘腦、腹側被蓋區、海馬體、大腦皮質、背側迷走神經復合體)[32]。下丘腦內生長激素促分泌素受體的激活可直接提高促食欲神經元的活性,引起突觸連接方式的改變,從而促進食欲回路的激活,同時抑制促進飽腹感的通路[33]。研究指出,攝入果糖后引起外周循環中胃饑餓素水平升高可導致食欲亢進[34-35],這可能與外周胃饑餓素穿過血腦屏障作用于多個大腦回路有關。胃饑餓素可直接激活弓狀核內神經肽Y/AgRP及外側下丘腦內增食欲素神經元,并抑制POMC神經元[36];還可通過激活Gq/11蛋白介導的磷脂酶C/三磷酸肌醇信號通路及Gs蛋白介導的環磷酸/蛋白激酶A信號通路激活AMPK[32,37],磷脂酶C/三磷酸肌醇信號通路及AMPK自身可促進細胞內鈣水平升高,有利于2-花生四烯酸甘油生成,EC可間接激活下丘腦AMPK,AMPK的激活(磷酸化)抑制乙酰輔酶A羧化酶和脂肪酸合酶,介導細胞質內MCoA和長鏈脂肪酰輔酶A的減少,促進下丘腦內脂肪酸氧化[33,38]。AMPK介導的脂肪酸氧化過程中,線粒體內解偶聯蛋白-2水平上調,神經肽Y和AgRP的轉錄水平升高,最終驅動攝食行為[39]。另外,胃饑餓素可作用于邊緣獎賞系統,亦可同時作用于突觸前和突觸后的生長激素促分泌素受體以激活多巴胺神經元[33],在動物外周或腦室內注射胃饑餓素均能引起腹側被蓋區及伏隔核中多巴胺水平升高[31],可觀察到動物攝食量增加,以及更傾向選擇高熱量的食物[40]。正常體重人群進行享樂性進食時血漿中胃饑餓素和2-花生四烯酸甘油水平升高,提示胃饑餓素與ECS在獎賞相關反應的調節中存在聯系[24]。胃饑餓素亦可激活人體邊緣獎賞系統的其他區域(如杏仁核、眼窩額葉皮質和紋狀體),增加進食快感及攝食渴求[8-9]。
2.2胰島素 中腦邊緣多巴胺獎賞回路主要由腹側被蓋區、伏隔核、杏仁核、海馬等構成,腹側被蓋區是獎賞索求和抑制攝食的關鍵部位[41]。已知胰島素作用于CNS內相應受體可產生飽腹感,并削弱邊緣獎賞系統及大腦皮質對食物刺激的反應[5,21],其機制為胰島素直接抑制腹側被蓋區中多巴胺能神經元的活性,或間接通過EC介導的長時程抑制多巴胺能神經元上的興奮性突觸傳遞[22],還可通過抑制下丘腦食欲調節神經肽的表達[5,8],實現飽腹效應。研究發現,與普通飲食以及加用葡萄糖飲料的動物和人類的胰島素水平相比,加用果糖飲料胰島素水平更低[11-12,35],且血糖水平無明顯上升[42],攝入果糖不引起外周胰島素水平上升可能與胰島β細胞內低表達葡萄糖轉運體5有關,認為果糖基本不刺激胰島素分泌[35],并指出長期果糖攝入可引起胰島素抵抗及下丘腦胰島素信號紊亂,抑制CNS內胰島素信號轉導,從而降低飽腹感[8]。
2.3瘦素 瘦素是脂肪組織分泌的抑制食欲性激素,通過瘦素受體調控下丘腦內多個與食欲相關的系統。瘦素可通過激活乙酰輔酶A羧化酶、降低AMPK活性增加弓狀核內MCoA的合成,通過AMPK/MCoA信號通路調節下丘腦神經肽的分泌和表達[8,43-44];瘦素是下丘腦EC水平的強調節劑,通過降低細胞內鈣水平抑制EC合成和信號轉導,間接影響下丘腦神經肽的表達[24]。在下丘腦CB1R部分敲除的小鼠中發現,瘦素介導的抑制攝食的作用減弱,提示瘦素通過下丘腦CB1R信號抑制食欲[24]。瘦素可直接介導下丘腦食欲神經肽的表達,直接刺激弓狀核中的POMC神經元,促進蛋白水解產物α-黑色素細胞刺激素釋放,抑制AgRP及神經肽Y分泌,還可刺激鄰近POMC神經元釋放阿片類物質以抑制外周釋放胃饑餓素[8]。果糖對瘦素水平影響的研究結果并不一致,可能與瘦素水平受體脂分布與體脂含量影響有關[4,45]。研究發現,長期喂食果糖的小鼠出現瘦素敏感性下降及瘦素抵抗,并與果糖飲食引起的高三酰甘油血癥有關,三酰甘油可導致瘦素血腦轉運障礙、瘦素受體基因表達下降及細胞因子信號轉導抑制劑基因表達增加[11]。外周循環中瘦素水平下降可刺激AgRP及神經肽Y的表達,抑制POMC和可卡因-苯丙胺調節轉錄肽表達[46],而神經肽Y表達增加與瘦素抵抗直接相關[11]。故果糖介導外周瘦素水平下降以及瘦素抵抗,影響CNS內瘦素相關信號轉導,從而削弱了瘦素介導的飽腹效應。
2.4GLP-1 GLP-1是腸道L細胞和孤束核分泌的一種短效飽腹肽類激素,與胰島β細胞中GLP-1受體結合刺激胰島素分泌,同時具有延遲胃排空、抑制食欲和攝食等效應[47]。其中,GLP-1的抑制食欲作用是最早發現的胰外效應,GLP-1受體激活介導食欲抑制及攝食量降低效應主要是通過抑制下丘腦的活性實現[18],此外也可通過谷氨酸受體α-氨基-3-羥基-5-甲基-4-異唑丙酸/紅藻氨酸介導的多巴胺能神經元突觸興奮性增加實現[48]。果糖可降低餐后GLP-1的上升幅度,延遲GLP-1釋放[21,35],削弱GLP-1激活對AMPK活性的抑制作用[18],從而降低GLP-1介導的飽腹效應。
3 展 望
果糖不僅作用于下丘腦的食欲調節相關信號通路,還可通過介導大腦不同區域神經元的活動實現促食欲效應,通過研究果糖對CNS和外周胃腸激素形成的復雜食欲調節網絡的影響,有利于理解果糖在分子學層面與攝食行為改變之間的關系。目前關于果糖在調控食欲方面的研究較少,對于予以果糖后各種食欲調節相關因子及下丘腦神經肽基因水平的變化尚未完全了解,目前研究方向集中在糖類對食欲調節相關通路的酶類及相應受體、轉運蛋白受體、糖代謝相關酶類基因水平的影響,但基因表達水平上調并不一定意味著編碼蛋白質表達水平的增加,為準確判斷具體基因或編碼產物調節食欲的機制,應進一步探討果糖攝入后食欲調節相關通路基因在蛋白轉錄水平的變化;同時對于食欲因子在食欲調控機制中的作用亦需進一步研究。
猜你喜歡
人人健康(2023年26期)2023-12-07 03:55:46
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
中國生殖健康(2019年2期)2019-08-23 08:12:10
人大建設(2019年12期)2019-05-21 02:55:32
學苑創造·A版(2015年11期)2016-01-14 09:03:27
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
中國醫藥科學(2015年15期)2015-02-27 12:32:27
中國全科醫學(2013年36期)2013-01-25 06:20:58
