壇紫菜泛素連接酶PhCUL1 基因克隆與功能驗證*
2021-03-19 06:19:16林鍵章王文磊紀德華陳昌生謝潮添
漁業科學進展 2021年1期
林鍵章 王文磊 徐 燕 許 凱 紀德華 陳昌生 謝潮添
(農業農村部東海海水健康與養殖重點實驗室 福建省水產生物育種與健康養殖工程研究中心集美大學水產學院 廈門 361021)
紫菜是一類具有重要經濟價值、營養價值和生態價值的大型海藻,不僅可以食用,還被廣泛用作肥料、藥物和化學物質的來源(Blouin et al, 2011),而且作為“藍碳”物種和生物燃料,具有巨大的潛力(Nellemann et al, 2009; Xu et al, 2016)。經估算,大型海藻每年可固定約173 Tg 的碳,并作為碳供體將90%固定的碳輸送至深海,其余則埋藏在沿海沉積物中,該估算值超過沿海生境中被子植物的碳埋藏估值(Krause-Jensen et al, 2016、2018),表明大型海藻在固碳方面有著巨大潛力。此外,由于紫菜等大型海藻含有較高的結構多糖和低木質素、不占用耕地等原因,其為生物燃料的優質原料(Ghadiryanfar et al, 2016; Sudhakar et al,2018)。Duarte 等(2017)指出,養殖海藻全部用于生物燃料生產,每平方千米養殖海藻每年可減少1500 t CO2排放。2018 年,我國栽培紫菜產量超過201779 t(干重),占養殖海藻產量的8.6%,其中,以壇紫菜(Pyropia haitanensis)為主要栽培對象的福建省紫菜產量已超過 74628 t (干重)(中國漁業統計年鑒,2019),帶來了可觀的經濟效益和生態效益。
然而,近年來,隨著全球氣候變暖,溫度成為限制紫菜產業發展的主要因素(陳偉洲等, 2015; Wang et al, 2018)。盡管一些壇紫菜耐高溫品系(Z-26、Z-61和 ZS-1)已廣泛栽培(Chen et al, 2008; Yan et al,2010),但有關壇紫菜的耐高溫機制仍然知之甚少。因此,闡明壇紫菜的耐高溫機制對耐高溫品系的選育具有重要意義。目前,對于壇紫菜耐高溫機制的研究主要集中在比較組學分析,例如,Wang 等(2018)比較了Z-61 和野生型壇紫菜的轉錄組,發現Z-61 可以通過更為積極的轉錄調節抵御高溫環境。Shi 等(2017)使用iTRAQ 技術在壇紫菜中鑒定了151 種應激反應蛋白,其中,參與清除未折疊蛋白或變性蛋白的泛素蛋白酶體系統在壇紫菜應答高溫脅迫過程中顯著上調表達,但其具體功能有待進一步驗證。
泛素蛋白酶體系統是最重要的脅迫響應系統之一,通過降解受損或多余的蛋白質來維持細胞中蛋白質的平衡。泛素化過程由3 種酶介導:E1(泛素激活酶,UBA);E2(泛素結合酶,UBC);E3(泛素連接酶)(Zhang et al, 2015)。其中,E3 連接酶識別配體蛋白并決定蛋白質降解的高特異性,因此,在泛素化過程中起關鍵作用。許多研究表明,E3 是植物響應非生物脅迫的“調節劑”,甚至單個E3 可以調節植物響應多重非生物脅迫(Moin et al, 2019)。Cullin-RING 連接酶是真核生物中被廣泛研究的多亞基E3 連接酶,在植物中有3 種類型:CUL1、CUL3a/b 和CUL4 (Stone,2014)。CUL3 可以促進DREB2A 的降解來提高擬南芥(Arabidopsis)的耐熱性(Morimoto et al, 2017)。此外,水稻Cullin 家族基因在應對非生物脅迫時呈現上調表達的趨勢(Moin et al, 2019)。以上結果說明,Cullin E3 連接酶可能在植物響應非生物脅迫中發揮重要作用。本實驗室在前期研究中發現,壇紫菜的CUL1 基因也積極響應高溫脅迫(Wang et al, 2018),但其具體功能有待進一步實驗驗證。
由于壇紫菜的特殊結構及缺乏啟動子和轉化方法等問題,目前尚未在壇紫菜中建立一套穩定的遺傳操作體系,對這些候選基因和蛋白的功能還不明確。萊茵衣藻(Chlamydomonas reinhardtii)作為一種真核單細胞綠藻,被廣泛用于驗證基因的功能和基本的生物學進程。同時,隨著基因工程技術的發展,已開發出一些可應用于萊茵衣藻的遺傳操作技術,包括核轉化、葉綠體和線粒體轉化系統(Dejtisakdi et al, 2016)。到目前為止,許多異源基因已在萊茵衣藻中成功表達,包括微藻Chlorella zofingiensis (Cordero et al,2011)和大型海藻Porphyra seriata (Kim et al, 2011)。Jin 等(2017)成功將甘紫菜(Pyropia tenera)HSP19.3 轉入萊茵衣藻,并證實了其在耐熱方面的作用。以上結果表明,萊茵衣藻可能是表達壇紫菜基因的良好載體。因此,本研究將壇紫菜的PhCUL1 基因轉入萊茵衣藻進行耐熱性分析,探究壇紫菜泛素連接酶(PhCUL1)在抗高溫中的作用,為壇紫菜耐高溫品系的選育提供基因信息。
1 材料與方法
1.1 實驗材料培養
本實驗所選用的壇紫菜耐高溫品系Z-61 由集美大學壇紫菜種質改良和應用實驗室選育和純化。培養條件:溫度(21±1)℃,光照強度50~60 μmol/(m2?s),光周期12 L∶12 D,每2 d 更換1 次新鮮的Provasoli's enrichment solution (PES)培養液,并充氣培養。培養藻體長度至15 cm 左右時,選取表面光滑、無破損、無扭曲的健康藻體進行后續實驗。
萊茵衣藻細胞壁缺失株系“CC-400 cw15 mt+”被用于PhCUL1 轉基因功能驗證。萊茵衣藻細胞在Trisacetate-phosphate (TAP)培養液中培養:溫度25℃,光照強度50 μmol/(m2?s),光周期14 L∶10 D。每隔2 h 輕搖1 次培養液,防止細胞貼壁生長。
1.2 總RNA 的分離純化和cDNA 的合成
采用E.Z.N.A 植物RNA 提取試劑盒(OMEGA,德國)提取壇紫菜和萊茵衣藻的總RNA。使用1%凝膠電泳檢測所提取總RNA 的完整性,并在紫外分光光度計上分別測定OD260nm和OD280nm值,計算出RNA的濃度,判斷其純度。
用于實時熒光定量PCR 分析的cDNA 用PrimeScript RT reagent kit (TaKaRa)進行反轉錄,合成的cDNA 稀釋10 倍備用。
1.3 壇紫菜PhCUL1 基因的全長克隆
根據壇紫菜轉錄組的注釋結果,篩選出一條注釋結果為臍形紫菜(Porphyra umbilicalis) CUL1 的Unigene0010232。根據序列設計1 對擴全長引物(正向引物加KpnⅠ酶切位點序列,反向引物加XbaⅠ酶切位點序列,便于后續實驗) PhCUL1F 和PhCUL1R(表1),PCR 擴增PhCUL1 基因的保守序列,預期長度為2481 bp;擴增體系為25 μl:2×Mix 12.5 μl,ddH2O 9.5 μl,每條引物0.5 μl,cDNA 模板2 μl。PCR擴增程序:95℃變性5 min;95℃ 40 s;62℃ 30 s;72℃ 2 min 30 s,35 個循環;72℃,延伸10 min。將擴增產物割膠回收、轉化和測序,驗證全長克隆的正確性。

表1 實驗用到的引物名稱和序列Tab.1 Primers and sequences used in this experiment
1.4 PhCUL1 基因的生物信息學分析
使用在線軟件NCBI(http://www.ncbi.nlm.nih.gov)分析PhCUL1 基因核苷酸序列,推導氨基酸序列和開放閱讀框(ORF)。通過 SMART (http://smart.emblheidelberg.de/)來識別保守結構域。使用MEGA6 軟件的Neighbor-Joining 法構建PhCUL1 系統進化樹。
1.5 PhCUL1 衣藻表達載體構建
使用KpnⅠ和XbaⅠ內切酶雙酶切目的基因和衣藻表達載體V3,載體圖譜見圖1。酶切體系:目的基因2 μg,10×M 緩沖液3 μl,KpnⅠ內切酶1 μl,XbaⅠ內切酶1 μl,用ddH2O 補足體系。V3 載體1 μg,10×M 緩沖液2 μl,KpnⅠ內切酶1 μl,XbaⅠ內切酶1 μl,用ddH2O 補足體系。酶切程序:37℃,140 min,65℃,10 min。將酶切后的基因和V3 載體16℃過夜連接,轉化和涂板。挑取陽性菌落進行PCR 驗證,獲得含有目的基因的V3 衣藻表達載體。
1.6 萊茵衣藻核轉化
萊茵衣藻核轉化采用“玻璃珠轉化法”。萊茵衣藻“CC-400 cw15 mt+”在TAP 培養液中培養至對數生長期(細胞數約為2×106個),室溫6000 r/min 離心5 min,棄上清液,倒置1 min。用300 μl TAP 培養液重懸細胞,加入到含300 mg 滅菌并烘干的玻璃珠的2 ml 離心管中,再加入0.1 ml 20% PEG 6000,2 μg 線性重組載體,5 μl 鮭魚精DNA。置于漩渦振蕩器上震蕩30 s,停歇30 s,重復2 次。把混合液加入到40 ml新鮮TAP 培養液,在100 r/min 搖床過夜培養。室溫6000 r/min 收集細胞,去上清液,用400 μl TAP 培養液重懸細胞,混勻后倒在TAP 平板上(含10 μg/ml 潮霉素B),24℃光照培養7~10 d,平板上長出單個克隆藻。
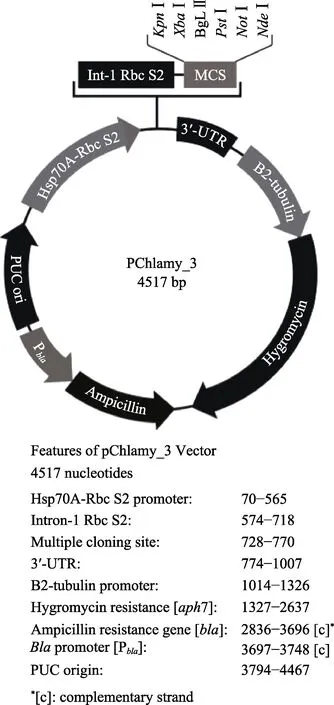
圖1 衣藻表達載體V3 圖譜Fig.1 V3 expression vector of C. reinhardtii
1.7 PhCUL1 基因表達水平的qPCR 分析
根據PhCUL1 的序列,設計qPCR 正反向引物,并以 β-tubulin 基因作為內參(表 1),進行高溫下PhCUL1 在萊茵衣藻中的表達水平分析。20 μl 體系:10 μl 2×SYBR green Master Mix(TaKaRa, 日本),0.4 μl正反向引物(20 mmol/L),2 μl 稀釋模板,0.4 μl ROX Dye 和6.8 μl ddH2O。擴增程序:95℃ 30 s,95℃ 5 s,60℃ 31 s,40 個循環。循環結束后,檢查擴增的熔解曲線,確保擴增的特異性。熒光定量PCR 擴增在ABI7300 型定量PCR 儀(Applied Biosystems, 美國)上進行。
1.8 數據統計分析
所有實驗均設置3 個重復。利用Excel 和SPSS 17.0 對實驗數據進行統計分析,并采用單因素方差分析(One-way ANOVA)和最小顯著差異法(LSD)比較不同數據組間的差異,P<0.05 表示存在顯著差異,P<0.01表示差異極顯著。
2 結果
2.1 PhCUL1 基因的克隆及序列分析
以壇紫菜cDNA 為模板,PhCUL1F/PhCUL1R 為引物進行擴增,獲得1 條擴增條帶,長度約為2500 bp(圖 2a)。通過 BLAST 比對發現,其與 Porphyra umbilicalis (GenBank No.: OSX79849.1)的cullin-1 基因核酸序列同源性達到85.14%,進而推斷該片段為PhCUL1 的基因片段。
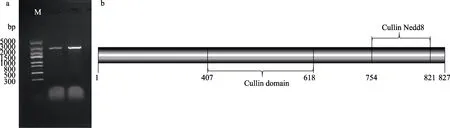
圖2 PhCUL1 基因克隆產物凝膠電泳和保守結構域Fig.2 Agarose electrophoresis of PCR products of PhCUL1 gene and protein domain analysis of PhCUL1
通過ORF Finder 軟件分析發現,PhCUL1 基因的ORF 為2481 bp,起始密碼子為ATG,終止密碼子為TGA,編碼827 個氨基酸,預測分子量為91.34 kDa,理論等電點為6.43。通過SMART 分析其保守結構域,發現PhCUL1 存在1 個Cullin (407~618 aa) 結構域和1 個Cullin Nedd8 (754~821 aa)結構域(圖2b)。
2.2 PhCUL1 基因系統進化樹分析
為了確定PhCUL1 的系統進化關系,采用Neighbor-Joining 法構建了PhCUL1 氨基酸序列系統進化樹。進化樹結果顯示(圖 3),PhCUL1 與紅藻門角叉菜(Chondrus crispus)、龍須菜(Gracilariopsis chorda)和臍形紫菜(Porphyra umbilicalis)聚為一側,而與高等植物明顯區分開來,這說明PhCUL1 基因在紅藻門中較為保守。而在紅藻門中,PhCUL1 與臍形紫菜聚為一支,這說明PhCUL1 基因在紫菜中較為保守。
2.3 PhCUL1 基因在高溫脅迫下的差異表達變化
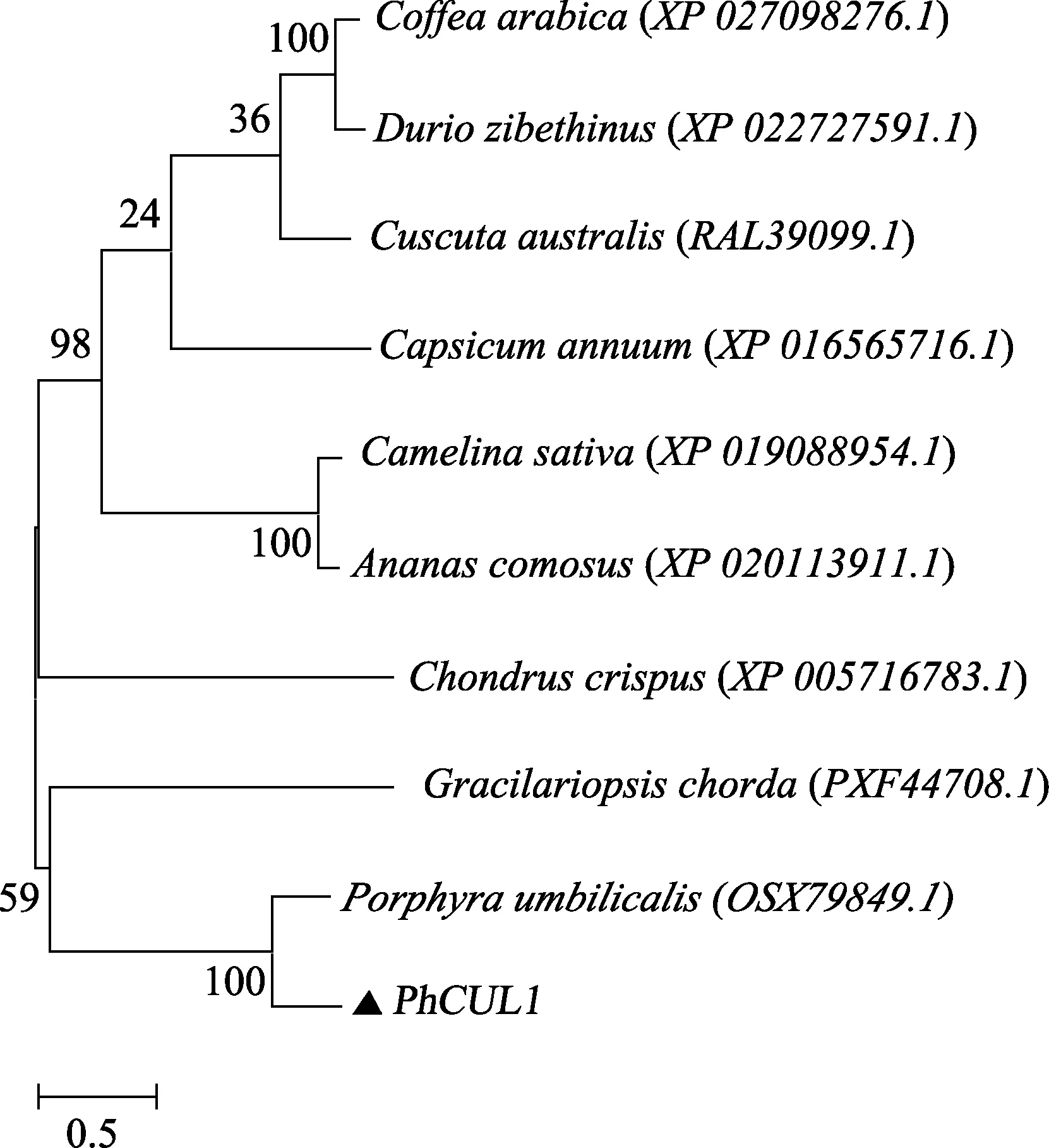
圖3 采用NJ 法構建的基于PhCUL1 基因所編碼氨基酸序列的系統進化樹Fig.3 Phylogenetic tree based on PhCUL1 amino acid sequences by NJ method
為了研究壇紫菜PhCUL1 對高溫脅迫的響應,本研究采用熒光定量PCR 技術檢測PhCUL1 基因在高溫31℃處理不同時間水平(0、3、6、12 和24 h)下的相對表達水平變化。結果顯示(圖4),高溫對PhCUL1基因表達水平有顯著影響,高溫處理3 h,PhCUL1的表達量是0 h 的11 倍;高溫處理6 h 和12 h,表達水平較之于高溫處理3 h 有所下降,但顯著高于0 h,其表達量分別是0 h 的5.1 倍和8.0 倍。高溫處理24 h后,PhCUL1 的表達量達到最高值,是0 h 的16.9 倍。
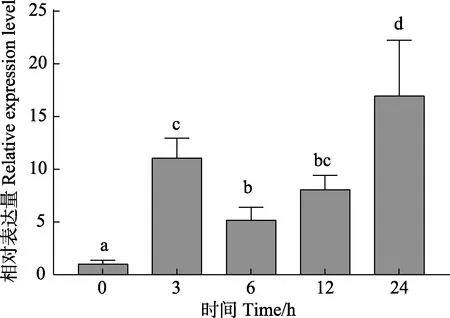
圖4 壇紫菜PhCUL1 基因在高溫31℃脅迫過程中相對表達變化Fig.4 The relative expression of PhCUL1 gene in P. haitanensis under 31℃ treatment
2.4 轉PhCUL1 萊茵衣藻陽性克隆子的篩選
隨機挑選8 個潮霉素B 抗性品系的衣藻,提取DNA,使用PhCUL1 特異引物(表1)進行PCR 驗證。瓊脂糖凝膠電泳結果顯示,所有挑選出的衣藻都檢測到陽性PCR 產物,但在野生型品系中未發現陽性克隆,表明PhCUL1 基因已成功轉入衣藻基因組(圖5)。選擇第4 個轉基因品系(T4)進行后續功能分析。
2.5 轉PhCUL1 衣藻對高溫脅迫的耐受性分析
為了研究轉基因衣藻對高溫脅迫的耐受程度,在33℃下培養野生型和轉基因衣藻36 h,每天觀察其生物量(圖6),并測定其在750 nm 的吸光度(表2)。結果顯示,高溫33℃脅迫12 h 后,轉基因衣藻(T4)的生物量高于野生型衣藻(WT)。高溫33℃脅迫36 h 后,轉基因衣藻(T4)的生物量明顯高于野生型衣藻(WT)。相應的OD750nm也與此結果吻合(表2)。

圖6 高溫33℃脅迫下衣藻的生物量變化Fig.6 Biomass of C.reinhardtii under 33℃ treatment
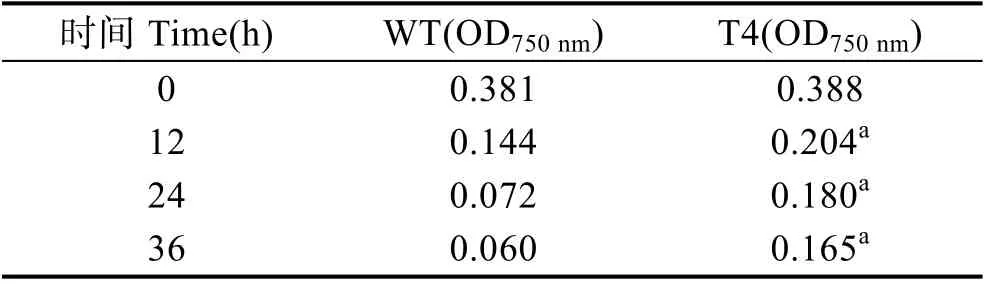
表2 高溫33℃脅迫下衣藻濃度的變化Tab.2 Concentration of C. reinhardtii under 33℃ heat stress
2.6 PhCUL1 基因在萊茵衣藻中的差異表達變化
高溫33℃脅迫1~6 h 后,PhCUL1 基因的表達水平顯著升高(P<0.05)(圖7),且在脅迫3 h 時,基因的表達水平最高,約為對照組的3 倍。脅迫9 h后,PhCUL1 表達水平下降,與對照組無顯著差異。
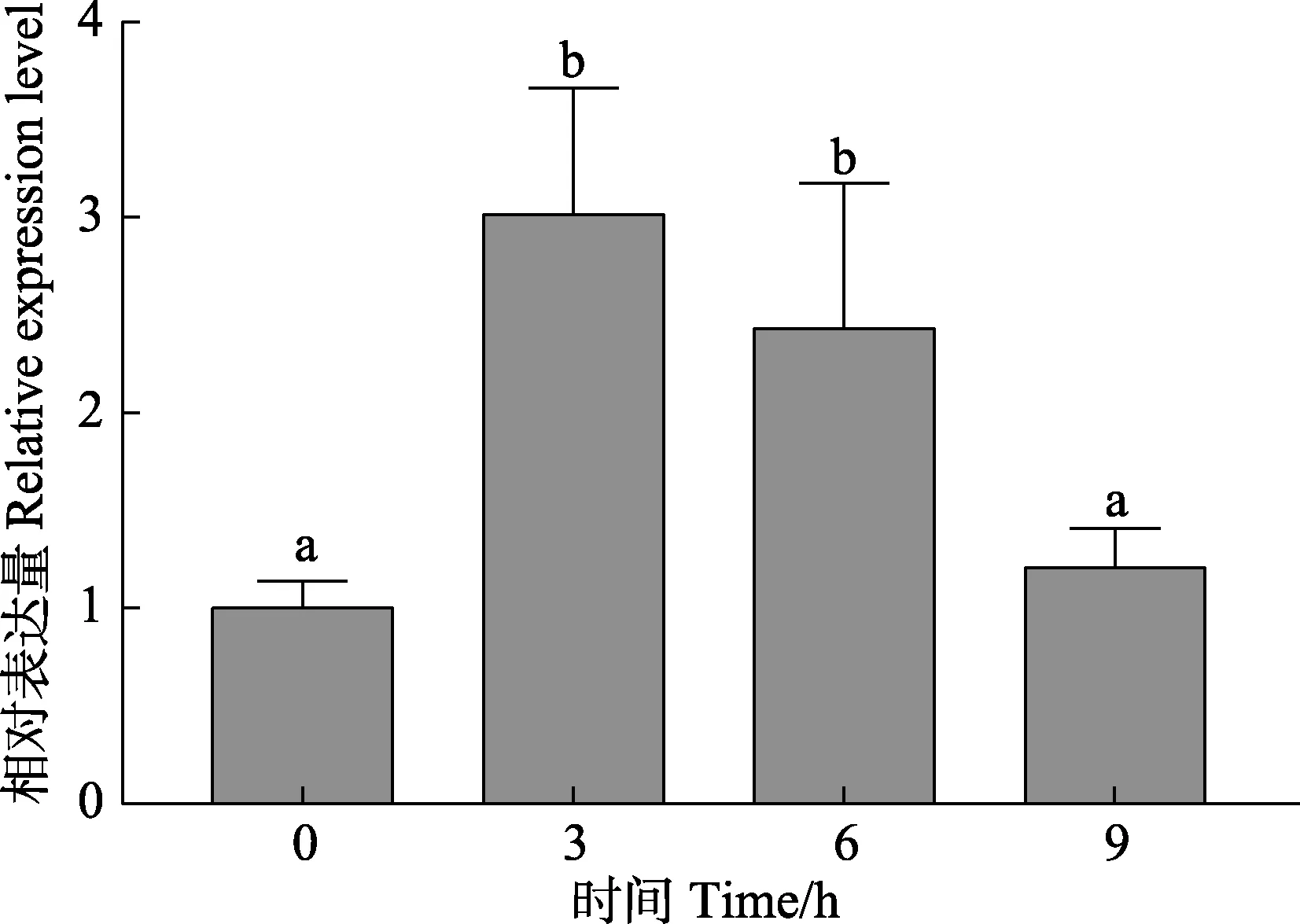
圖7 高溫33℃脅迫下PhCUL1 基因在萊茵衣藻中的相對表達水平Fig.7 The relative expression level of PhCUL1 gene in C. reinhardtii under 33℃ treatment
3 討論
溫度是影響紫菜產業發展的一個主要因素。近年來,頻繁出現的秋季高溫回暖天氣嚴重影響了壇紫菜栽培工作的有序開展,水溫過高導致壇紫菜幼苗或成菜發生病爛,造成嚴重減產(陳玉婷等, 2015)。進一步研究壇紫菜的耐高溫機理對于快速選育耐高溫品系具有重要的理論意義。前期研究已證實,泛素系統在響應高溫、高鹽和干旱等非生物脅迫中扮演著重要角色(Zhang et al, 2015)。其中,E3 連接酶因其決定靶蛋白的特異性而被廣泛研究。Cullin-RING E3 連接酶是真核生物中一個被充分研究的基因家族,Cullin 蛋白一般含有1~3 個結構域(Moin et al, 2019)。本研究克隆獲得的壇紫菜PhCUL1 基因含有2 個保守結構域,407~618 位點是cullin 結構域,754~821 位點是Cullin Nedd8 結構域。其中,Nedd8 結構域是蛋白融合位點,它能賦予cullin E3 泛素連接酶活性(Hori et al,1999),表明PhCUL1 也可能具有泛素連接酶活性,Nedd8 是PhCUL1 介導泛素連接酶活性的體現。
已有大量文獻報道和證據表明,cullin 家族基因可以積極響應非生物脅迫。例如,Moin 等(2019)在研究水稻的逆境脅迫表達譜時發現,水稻cullin 家族基因在多種逆境脅迫下上調表達。本研究也分析了壇紫菜PhCUL1 基因在高溫脅迫下的表達模式。研究結果顯示,在高溫31℃脅迫下,PhCUL1 基因在不同時間點均呈現上調表達趨勢;高溫脅迫 3 h 時,PhCUL1 基因的表達量是0 h 的11 倍,說明PhCUL1基因對高溫脅迫反應非常迅速,短時間內可以大量積累表達;隨著脅迫時間的延長,PhCUL1 基因的表達量有所下調,但仍然高于0 h,高溫脅迫6 h 和12 h的PhCUL1 基因表達量分別是0 h 的5.1 倍和8.0 倍;當脅迫時間持續到24 h 時,PhCUL1 仍然處于高表達狀態,其表達量是0 h 的16.9 倍。以上結果表明,PhCUL1 在高溫脅迫過程中均上調表達,可能是壇紫菜響應高溫脅迫的一個關鍵基因,但具體功能需進一步驗證。
隨著基因工程技術的發展,許多研究人員利用遺傳操作手段對E3 連接酶的功能進行了驗證。Peng 等(2019)發現,過表達擬南芥E3 連接酶AtPUB48 可以增強擬南芥種子發芽和幼苗生長階段的耐熱性。此外,過表達水稻E3 連接酶OsHIRP1 使水稻在高溫下具有比對照組更高的萌發率(Kim et al, 2019)。然而,有關E3 連接酶基因在大型海藻抗逆過程中發揮的具體功能尚未發現報道。本研究在萊茵衣藻中轉入PhCUL1 基因來研究其耐熱功能。qRT-PCR 結果顯示,高溫33℃脅迫下轉基因衣藻的PhCUL1 基因顯著上調表達,表明該基因能被高溫脅迫誘導。生理實驗結果顯示,在持續的高溫脅迫下,轉基因衣藻比野生型具有更高的存活率,證明PhCUL1 基因可能通過維持蛋白穩態調控了壇紫菜的耐熱過程。當然,一個機體對非生物脅迫的調控機制是非常復雜的,至于PhCUL1 基因是如何在這個復雜的網絡中起作用,還有待進一步研究。
4 結論
本研究成功克隆了壇紫菜的泛素連接酶PhCUL1基因,而且該基因被高溫顯著誘導。轉入PhCUL1 基因的萊茵衣藻也比野生型衣藻具有更強的耐熱性,證明了PhCUL1 基因的耐熱功能。這為壇紫菜耐高溫機制的解析提供了有用的基因信息,也為耐高溫品系的培育奠定了理論基礎。
