水稻單倍體誘導(dǎo)基因OsMATL突變體的創(chuàng)制與分析
2021-03-18 03:16:04賈思思王加峰黃翠紅陳志強
作物學(xué)報 2021年5期
關(guān)鍵詞:水稻
文 欽 賈思思 王加峰 黃翠紅 王 慧 陳志強 郭 濤
華南農(nóng)業(yè)大學(xué) / 國家植物航天育種工程技術(shù)研究中心, 廣東廣州 510642
水稻(Oryza sativaL.)是我國重要的糧食作物之一, 水稻的產(chǎn)量和品質(zhì)直接影響著我國糧食安全與人民生活品質(zhì)[1]。水稻品種的高效選育是提高水稻產(chǎn)量、品質(zhì)和抗性的關(guān)鍵環(huán)節(jié)。傳統(tǒng)育種中需要6~8個世代連續(xù)自交和不斷選擇才能獲得相對穩(wěn)定的材料, 周期較長, 難以滿足水稻育種需求。雙單倍體育種(double haploid, DH技術(shù))只需2個世代就能獲得純系, 大大縮短了育種進程, 是純系選育的一次技術(shù)革命[2]。單倍體的產(chǎn)生是雙單倍體育種的第一步,自然發(fā)生單倍體的頻率很低, 僅為 0.1%, 無法滿足育種的大量需求[3]。在水稻中, 利用花藥進行離體培養(yǎng), 誘導(dǎo)花藥產(chǎn)生愈傷組織再分化成單倍體植株,是產(chǎn)生水稻單倍體的主要方法。但是, 水稻的花藥培養(yǎng)具有嚴(yán)格的基因型依賴且操作復(fù)雜, 如秈稻的花藥培養(yǎng)能力顯著弱于粳稻[4]。因此, 水稻單倍體的產(chǎn)生一直是制約著水稻雙單倍體育種的瓶頸。
近年來, 在玉米中以生物誘導(dǎo)為基礎(chǔ)的 DH技術(shù)已被國內(nèi)外種業(yè)公司及育種單位廣泛的應(yīng)用, 成為了與分子標(biāo)記輔助育種和轉(zhuǎn)基因技術(shù)并稱的現(xiàn)代玉米三大育種技術(shù)[5-6]。生物誘導(dǎo)產(chǎn)生孤雌生殖單倍體是利用單倍體誘導(dǎo)系作父本, 與目標(biāo)選系基礎(chǔ)材料進行雜交, 在雜交當(dāng)代的籽粒上就能獲得一定比例的母本血緣單倍體。利用誘導(dǎo)系產(chǎn)生單倍體具有不受基因型依賴、誘導(dǎo)過程簡單、成本低廉等優(yōu)勢。在玉米中, 誘導(dǎo)單倍體產(chǎn)生的性狀受主效QTL控制,其中qhir1能夠解釋66%遺傳變異[5]。2017年美國、中國和法國科學(xué)家?guī)缀跬瑫r克隆qhir1位點的花粉特異性磷脂酶 A 基因MTL/ZmPLA1/NLD(GRMZM2G471240)[7-9]。在玉米單倍體誘導(dǎo)系中,該基因在第 4外顯子 1572~1573之間有個 4 bp(CGAG)插入, 造成 20個氨基酸移碼突變, 導(dǎo)致翻譯提前終止, 產(chǎn)生單倍體誘導(dǎo)功能。
MTL在作物中高度保守, 利用基因編輯技術(shù)已成功在玉米[7-8,10]、水稻[11-12]和小麥[13-14]中實現(xiàn)以誘導(dǎo)基因MTL為基礎(chǔ)的單倍體誘導(dǎo)。先正達公司[11]對秈稻 IR58025B中MTL同源基因OsMATL(Os03g0393900)第1外顯子和第4外顯子進行編輯,突變體可誘導(dǎo)產(chǎn)生2%~6%的單倍體。王春等[12]對秈粳雜交稻春優(yōu)84中OsMATL第4外顯子進行編輯后單倍體誘導(dǎo)率為4.44%。玉米中MTL基因不同位點突變后單倍體誘導(dǎo)率不同, 水稻中OsMATL基因不同位點的突變是否會影響單倍體誘導(dǎo)率還有待探索。此外, 水稻材料的不同遺傳背景對單倍體誘導(dǎo)率的影響也未知, 因此獲得一個高誘導(dǎo)率的水稻單倍體誘導(dǎo)系對水稻雙單倍體育種有著重要的意義。本研究以粳稻日本晴和秈稻華航48為研究材料, 利用CRISPR/Cas9技術(shù)對OsMATL基因啟動子區(qū)和編碼區(qū)的多個靶點進行編輯, 獲得了秈粳背景下不同類型的OsMATL突變體, 進一步分析了不同位點的突變效應(yīng), 為深入研究水稻孤雌生殖單倍體誘導(dǎo)的機制提供了基礎(chǔ)材料。
1 材料與方法
1.1 材料
研究材料為粳稻日本晴和秈稻華航48, 日本晴是典型的粳型純系品種, 華航48是國家植物航天育種工程技術(shù)研究中心選育的綜合性狀優(yōu)良的秈型純系品種。試驗材料水培種植于人工氣候室(溫度 28℃,光照10 h, 黑暗14 h), 所有材料常規(guī)管理。
1.2 OsMATL生物信息學(xué)分析
利用 Ensembl數(shù)據(jù)庫(https://plants.ensembl.org/index)獲取MTL的基因序列及氨基酸序列。通過NCBI數(shù)據(jù)庫中 Blastp (https://blast.ncbi.nlm.nih.gov/Blast.cgi)在線分析, 搜索MTL在谷物和模式作物擬南芥中的同源基因, 并下載同源基因的氨基酸序列。利用 Clustl Omega (https://www.ebi.ac.uk/Tools/msa/clustalo/)對氨基酸序列進行多序列比對[15-16]; 利用Pfam (http://pfam.xfam.org/)和UniProt(https://www.uniprot.org/)預(yù)測OsMATL基因蛋白結(jié)構(gòu)域。選取OsMATL基因ATG上游2000 bp作為啟動子區(qū)域, 利用順式作用元件在線分析網(wǎng)站PlantCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析啟動子序列, 預(yù)測順式作用元件[16]。利用SWISS MODEL網(wǎng)站(https://swissmodel.expasy.org/)進行蛋白三級結(jié)構(gòu)預(yù)測[17]。
1.3 靶點及引物設(shè)計
利用 CRISPR-GE網(wǎng)站(http://skl.scau.edu.cn/)設(shè)計靶點[18], 在OsMATL編碼區(qū)第1外顯子、第3外顯子和第 4外顯子處設(shè)計單靶點; 在啟動子區(qū)設(shè)計雙靶點對順式作用元件進行敲除, 靶點序列及位置如圖1。根據(jù)選擇的靶點設(shè)計引物(表1), 由金唯智(廣州)生物科技有限公司合成。
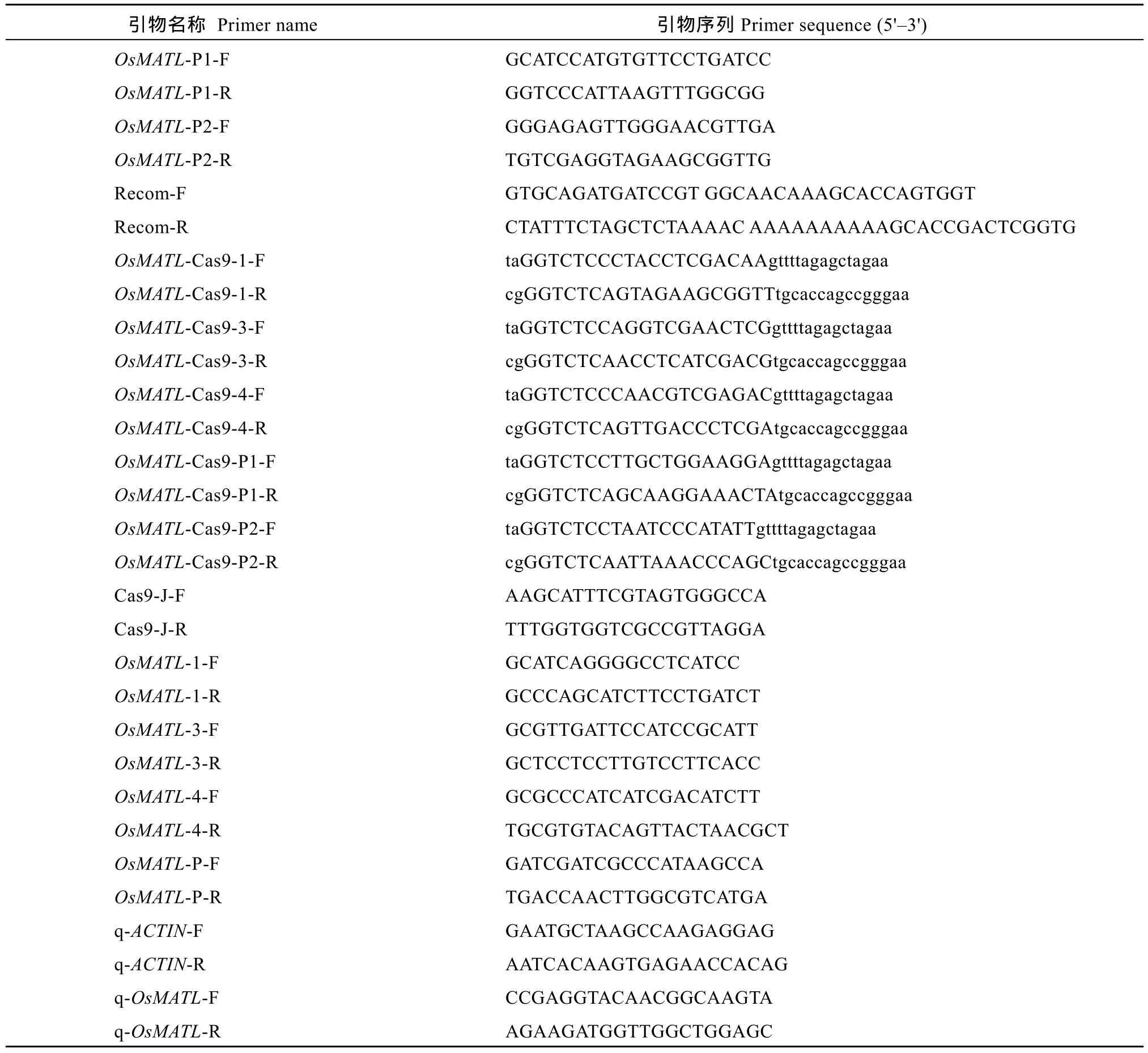
表1 本研究所用引物Table 1 Primer sequences used in this study
1.4 CRISPR/Cas9載體構(gòu)建
參照謝卡斌等[19]利用 TPG 系統(tǒng)構(gòu)建CRISPR/Cas9載體的方法。試驗步驟如下: (1)初級片段獲取。以pGTR質(zhì)粒為模板, PCR擴增出含有靶點和PTG載體的片段; (2) PTG片段組裝。利用Golden Gate克隆方式將初級片段連接起來; (3) pRGEB32載體酶切。利用BsaI限制性內(nèi)切酶酶切pRGEB32載體, 獲得與PTG相同的黏性末端; (4)重組載體構(gòu)建。將PTG片段插入到酶切后pRGEB32載體中BasI克隆位點; (5)重組載體轉(zhuǎn)化。用熱激法將重組的載體轉(zhuǎn)入大腸桿菌感受態(tài)細(xì)胞 DH5α中; (6)陽性克隆檢測。挑取單個菌落接種培養(yǎng), 通過測序驗證載體正確性。CRISPR/Cas9載體構(gòu)建如圖2所示。
1.5 農(nóng)桿菌介導(dǎo)遺傳轉(zhuǎn)化
參照Hiei等[20]建立的水稻高效農(nóng)桿菌介導(dǎo)的遺傳轉(zhuǎn)化法, 將構(gòu)建好的 CRISPR/Cas9載體通過電擊轉(zhuǎn)化農(nóng)桿菌EHA105中, 3個編碼區(qū)的單靶點載體分別轉(zhuǎn)化日本晴和華航48成熟胚誘導(dǎo)的愈傷組織中, 1個啟動子區(qū)的雙靶點載體轉(zhuǎn)化到華航48成熟胚誘導(dǎo)的愈傷組織中, 通過潮霉素抗性篩選T0代組培苗。
1.6 靶點檢測
提取轉(zhuǎn)基因水稻基因組 DNA, 在靶點上下游200 bp處設(shè)計引物, 對包含靶點目的片段進行擴增和測序, 將測序的結(jié)果放入在線解碼工具DSDecode(http://dsdecode.scgene.com/)解碼[21], 確定靶點突變情況。
1.7 OsMATL基因表達量分析
收集OsMATL啟動子區(qū)突變體和野生型散粉前新鮮的花藥, 于-80℃保存。采用TRIzol法提取成熟花粉RNA, 參照EvoM-MLV反轉(zhuǎn)錄試劑盒II說明書反轉(zhuǎn)錄合成cDNA。以cDNA為模板,actin為內(nèi)參, 參照AceQ qPCR SYBR Green Master Mix試劑說明書,在熒光定量PCR儀(ABI StepOnePlus)上進行qRT-PCR分析, 采用2-ΔΔCt法計算基因的相對表達量。
1.8 突變體結(jié)實情況調(diào)查
調(diào)查OsMATL編碼區(qū)純合突變體和OsMATL啟動子區(qū)大片段缺失突變體主穗上結(jié)實情況, 分為實粒(籽粒飽滿透明)、敗育(只剩薄薄種皮或籽粒皺縮)以及空粒(穎殼內(nèi)無籽粒), 并統(tǒng)計結(jié)實率和敗育率。利用SPSS進行單因素方差分析及 Duncan’s法多重比較, 差異顯著性水平為P≤0.01。
2 結(jié)果與分析
2.1 OsMATL生物信息學(xué)分析
對玉米MTL基因與水稻、高粱、小麥、大麥、小米和擬南芥中的MTL同源基因進行氨基酸序列多重比對(圖3)。結(jié)果顯示,MTL在谷類作物中的同源性較高, 達到70%以上, 玉米中MTL基因與水稻中同源基因OsMATL氨基酸序列相似性為 79%, 粳稻和秈稻中OsMATL基因氨基酸序列相似性高達99%。保守的氨基酸序列暗示MTL在谷類作物中具有相似的功能。水稻中OsMATL基因位于3號染色體上, 含有4個外顯子, 編碼432個氨基酸。該基因編碼花粉特異性磷脂酶, 有 1個類磷脂酶 A2結(jié)構(gòu)域、1個酰基轉(zhuǎn)移酶/酰基水解酶/溶血磷脂酶結(jié)構(gòu)域、1個聚乳酸結(jié)構(gòu)域、3個Motif (GXGXXG、GXSXG和DGA/G), 在C430處有S-棕櫚酰化或S-法尼酰化位點, 具有脂類分解代謝和植物防御的功能。
OsMATL啟動子與其他真核啟動子相似, 包含了典型的基礎(chǔ)轉(zhuǎn)錄調(diào)控的順式作用元件(圖4), 有47個TATA-box和36個CAAT-box, 其中TATA-box主要集中在0至-1000 bp處, CAAT-box分散分布在啟動子區(qū)。還有 4種逆境響應(yīng)元件, 分別為厭氧誘導(dǎo)的ARE元件、GC-motif元件、低溫脅迫相關(guān)的LTR元件和應(yīng)激脅迫相關(guān)的 TC-rich repeats元件, 這些元件可能參與了植物防御。5種(8個)光響應(yīng)元件, 如Box 4、G-box、MRE、TCT-motif和 TCCC-motif。此外還有4種(6個)激素響應(yīng)元件、1種(2個)生理調(diào)控相關(guān)元件、1種(2個)蛋白結(jié)合位點相關(guān)元件和若干功能未知的順式作用元件。
2.2 CRISPR/Cas9靶點分析
為了獲得不同位點的OsMATL突變體, 在OsMATL基因編碼區(qū)設(shè)計3個單靶點(圖1), 第1外顯子上的靶點距陳紹江團隊報道的CRISPR/Cas9靶點下游11 bp, 較為徹底的破壞OsMATL的功能; 第3外顯子上的靶點在DGA/G motif處; 第4外顯子上的靶點距玉米天然突變位點上游8 bp處, 與先正達公司和王克劍團隊在第 4外顯子上的靶點接近, 此靶點能夠減少非預(yù)期的影響, 獲得具有單倍體誘導(dǎo)功能的突變體。在啟動子區(qū)域-1200 bp到-1500 bp設(shè)計雙靶點敲除多個順式作用元件, 2個靶點之間距離293 bp (圖1),預(yù)期獲得OsMATL基因表達量具有差異的等位突變。
2.3 OsMATL突變位點分析
所有轉(zhuǎn)化均成功獲得T0代轉(zhuǎn)基因苗, 對T0代轉(zhuǎn)基因苗進行測序, 篩選OsMATL編碼區(qū)純合突變的植株和OsMATL啟動子區(qū)大片段缺失的植株。OsMATL編碼區(qū)上一共獲得了 56種純合突變體(表2), 純合突變率為19%。在第1外顯子的靶點位置, 日本晴中獲得 4種純合突變, 分別為單堿基 A和 T的插入、8個堿基的缺失、3個堿基的替換和10個堿基的缺失; 華航48中只獲得了單堿基 A插入的純合突變。這些突變類型均導(dǎo)致了移碼突變,其中10個堿基的缺失使得翻譯在第112個氨基酸處提前終止, 截短了 303個氨基酸。對比蛋白質(zhì)預(yù)測的結(jié)構(gòu)(圖5), 發(fā)現(xiàn)第1外顯子上突變體的三級結(jié)構(gòu)與野生型的完全不同, 蛋白結(jié)構(gòu)更簡單。
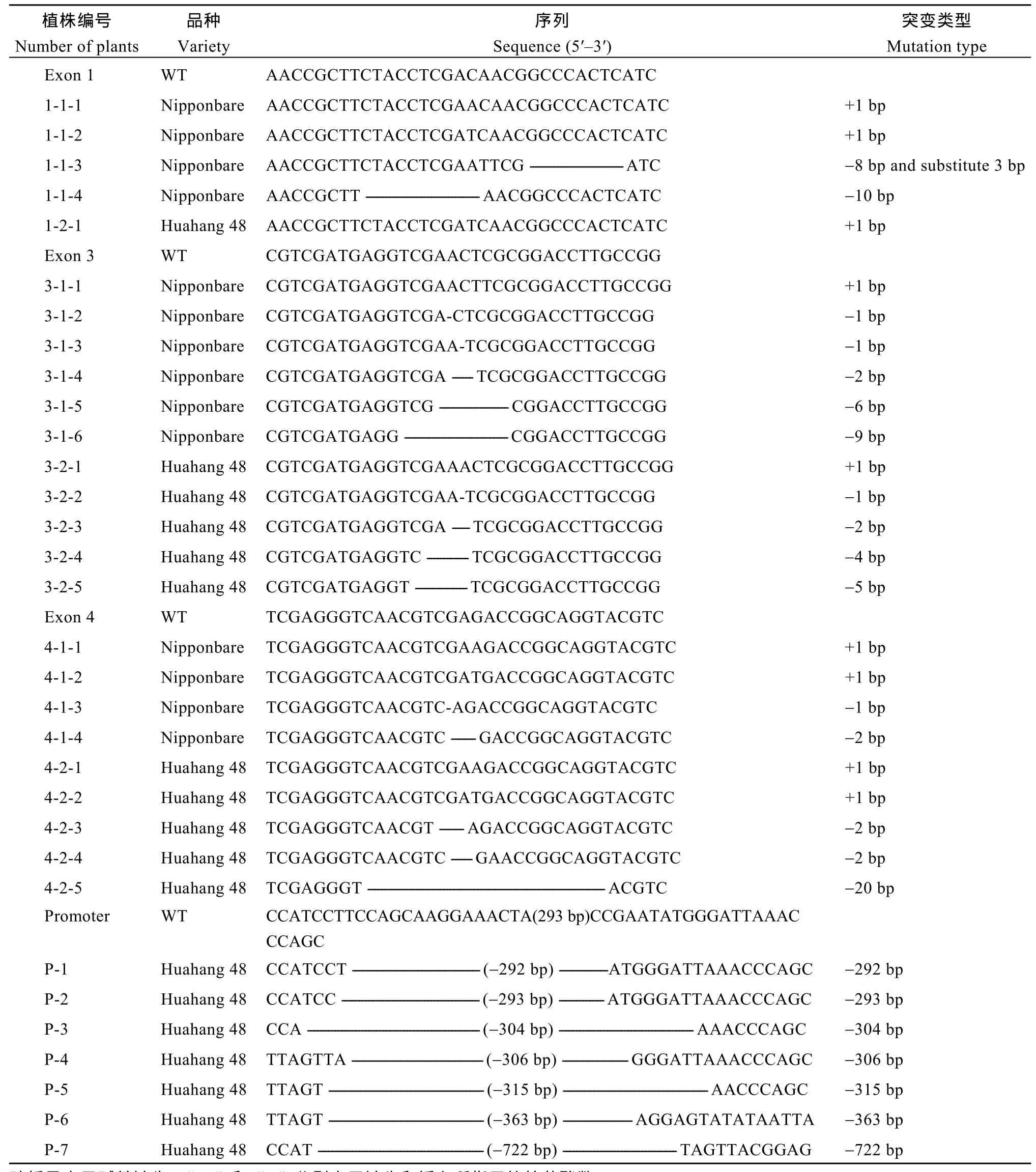
表2 T0代編碼區(qū)純合突變及啟動子區(qū)大片段缺失突變類型Table 2 Homozygous mutation in coding region and large fragment deletion mutation in promoter region types of T0 generation
在第3外顯子靶點位置, 獲得了單堿基A和T的插入、單堿基和多個堿基缺失的純合突變。在日本晴中獲得6種類型的純合突變, 其中6個堿基和9個堿基的缺失導(dǎo)致該位點編碼氨基酸的缺失, 而其他突變類型均引起移碼突變, 使得DGA/G motif喪失, 蛋白翻譯出現(xiàn)不同程度的提前終止。對比蛋白質(zhì)預(yù)測的結(jié)構(gòu)(圖5), 第 3外顯子上突變體的蛋白結(jié)構(gòu)較野生型簡單, 但比第1外顯子上突變體的蛋白結(jié)構(gòu)復(fù)雜。
在第4外顯子靶點位置, 日本晴和華航48中均獲得了單堿基A和T的插入以及2個堿基缺失的純合突變。此外, 在日本晴中獲得了單個堿基缺失的純合突變, 華航 48中獲得了 20個堿基缺失的純合突變。這些突變類型均導(dǎo)致了移碼突變, 其中單個堿基的缺失導(dǎo)致翻譯在第378個氨基酸處提前終止, 其他的移碼突變均未導(dǎo)致翻譯提前終止。第4外顯子上的突變只有少量 C端氨基酸改變, 蛋白三級結(jié)構(gòu)與野生型相似(圖5), 突變后的蛋白結(jié)構(gòu)細(xì)微的差異可能導(dǎo)致催化位點的喪失, 使得蛋白失去了原有的功能。
啟動子區(qū)獲得了7種含有大片段缺失的突變體(表2), 占33.33%。主要為300 bp左右的大片段缺失, 敲除了多個順式作用元件, 分別為 3個TATA-box、2個CAAT-box、4個光響應(yīng)元件和1個激素響應(yīng)元件。 此外, 還有1個缺失722 bp的突變體, 刪除了多個TATA-box和多種順式作用元件。進一步對啟動子區(qū)大片段缺失突變體OsMATL基因進行實時熒光定量PCR分析(圖5-E), 結(jié)果顯示啟動子區(qū)域293 bp的缺失顯著降低了OsMATL基因表達量,該種類型的突變可能影響OsMATL基因的功能。
2.4 OsMATL突變體結(jié)實情況分析
對OsMATL突變體結(jié)實情況進行調(diào)查(圖6和圖7), 日本晴和華航48中OsMATL編碼區(qū)突變體結(jié)實率分別為 2.87%~6.92%和 12.46%~15.26%, 均極顯著低于野生型,OsMATL編碼區(qū)突變后對秈稻結(jié)實的影響比粳稻小, 暗示秈稻更能忍OsMATL基因的功能缺失。日本晴中OsMATL編碼區(qū)突變體敗育率極顯著增加, 高達 51%以上; 華航 48中OsMATL編碼區(qū)突變體敗育率為11%~18%, 其中第4外顯子上突變體敗育率極顯著高于野生型。在日本晴和華航48編碼區(qū)突變體的后代中均發(fā)現(xiàn)不同程度的敗育籽粒, 大部分只剩薄薄的種皮內(nèi)部無內(nèi)含物, 小部分胚乳出現(xiàn)不同程度的皺縮(圖7-C, D)。華航48啟動子區(qū)域大片段的缺失對植株的結(jié)實幾乎沒有影響,暗示OsMATL基因表達量的改變與編碼氨基酸的改變調(diào)控雙受精的機制可能不同。
3 討論
2017年, 控制玉米單倍體誘導(dǎo)的關(guān)鍵基因MTL幾乎同時被 3個國家的科學(xué)家獨立克隆, 單倍體誘導(dǎo)系該基因第4外顯子處發(fā)生了4 bp的插入, 引起氨基酸的移碼突變, 導(dǎo)致MTL蛋白被截短[7-9]。全長的MTL蛋白被定位在精細(xì)胞膜上, 而突變后截短的MTL蛋白在精細(xì)胞膜上并未觀察到信號,MTL基因可能參與信號轉(zhuǎn)導(dǎo)或配子融合的過程, 突變后MTL蛋白在精細(xì)胞膜上的缺失可能是單倍體誘導(dǎo)的原因[22]。利用基因編輯技術(shù)對玉米中的MTL基因進行突變, 證明了編碼磷脂酶的MTL基因功能減弱或者喪失能夠誘導(dǎo)單倍體的產(chǎn)生。Kelliher等[7]利用TALEN技術(shù)對玉米NP2222在自然突變位點附件進行編輯, 使得部分肽段缺失, 單倍體誘導(dǎo)率為6.7%。劉晨旭等[8,23]對ZmPLA1第1外顯子進行編輯,獲得單堿基A插入、單堿基缺失和11個堿基缺失的等位突變, 造成較為徹底的基因突變, 單倍體誘導(dǎo)率為2%左右與Stock 6誘導(dǎo)率基本一致, 胚乳敗育籽粒頻率也顯著升高, 證明了MTL基因突變會導(dǎo)致玉米胚乳敗育。隨后董樂等[10]利用CRISPR/Cas9技術(shù)對玉米自交系ZC01的MTL基因天然突變位點上游5 bp處進行靶向突變, 獲得了一系列的MTL突變體, 以雜交種ZY7、ZY8和單99為受體測定單倍體誘導(dǎo)率, 驗證了所獲得的突變體具有較高的單倍體誘導(dǎo)能力, 平均單倍體誘導(dǎo)率為 7.32%, 此研究結(jié)果與Kelliher等的結(jié)果相近, 推測MTL基因突變位點和材料可能會影響單倍體誘導(dǎo)率。
MTL在作物中功能保守, 為以單倍體誘導(dǎo)基因為基礎(chǔ)的單倍體誘導(dǎo)系統(tǒng)在其他作物中的建立提供了途徑。先正達公司率先在水稻中實現(xiàn)單倍體誘導(dǎo),通過氨基酸同源序列比對在水稻中找到了MTL同源基因OsMATL, 對秈稻自交系IR58025B中OsMATL基因的第1外顯子和第4外顯子上天然突變位點下游7 bp處進行編輯, 突變體的結(jié)實率為20%, 單倍體誘導(dǎo)率2%~6%左右, 且單倍體誘導(dǎo)率受母本遺傳背景的影響, 與玉米MTL突變后結(jié)果相似, 證明了MTL在作物中功能保守[10-11]。王春等[12]對秈粳雜交稻春優(yōu)84中OsMATL基因第4外顯子上天然突變位點上游5 bp處進行編輯, 對單堿基C插入純合突變體以及單堿基C插入和單堿基缺失雙等位突變體進行分析, 突變體的結(jié)實率為11.5%, 單倍體誘導(dǎo)率為4.4%。劉晨旭等[13]將單倍體誘導(dǎo)系統(tǒng)擴展到六倍體小麥中, 利用 CRISPR/Cas9技術(shù)對小麥 CB037中TaPLA第1外顯子進行編輯, 成功獲得了小麥A、D基因組磷脂酶基因TaPLA的突變體, 該突變體結(jié)實率為 30%~60%, 可誘導(dǎo)產(chǎn)生約 5%~15%的單倍體,但同時在自交和雜交過程中還會產(chǎn)生大量的敗育籽粒和一定頻率的非整倍體籽粒。劉會云等[14]利用優(yōu)化的SpCas9系統(tǒng)對小麥Fielder和寧春4號的TaMTL基因設(shè)計雙靶點進行高效的編輯, 獲得了小麥A、B和D基因組3個TaMTL基因的突變體, T0代突變體結(jié)實率明顯降低, 三突結(jié)實率僅為19.8%~22.4%, T1代突變體單倍體誘導(dǎo)率可達到 18.9%, 并在后代中發(fā)現(xiàn)了胚缺失、胚和胚乳雙缺失的籽粒表型。利用CRISPR/Cas9技術(shù)對作物中單倍體誘導(dǎo)基因進行編輯可快速獲得單倍體誘導(dǎo)系, 至此單倍體誘導(dǎo)系統(tǒng)在三大糧食作物上均得以驗證。玉米、水稻和小麥中誘導(dǎo)基因突變后單倍體誘導(dǎo)率并不同, 單倍體誘導(dǎo)率可能與突變位點和遺傳背景有關(guān), 在不同突變位點和遺傳背景的材料中可能存在一些單倍體誘導(dǎo)的修飾位點。本研究對粳稻日本晴和秈稻華航48兩個遺傳背景不同的材料中OsMATL第1外顯子、第3外顯子、第 4外顯子及啟動子進行不同位置的編輯, 獲得了不同位點和不同遺傳背景的OsMATL突變體。本研究中OsMATL編碼區(qū)的移碼突變, 使得氨基酸在不同位置開始發(fā)生改變以及蛋白翻譯提前終止, 導(dǎo)致蛋白結(jié)構(gòu)發(fā)生不同程度的改變, 其中第1外顯子上突變后蛋白結(jié)構(gòu)與劉晨旭[23]對玉米MTL基因第 1外顯子突變后蛋白結(jié)構(gòu)變化相似, 突變后蛋白結(jié)構(gòu)更簡單。蛋白結(jié)構(gòu)不同程度的改變可能會影響OsMATL蛋白與細(xì)胞膜的結(jié)合能力, 從而誘導(dǎo)產(chǎn)生不同頻率的單倍體, 為在水稻中獲得高頻單倍體誘導(dǎo)系奠定了基礎(chǔ)。
單倍體誘導(dǎo)系在誘導(dǎo)單倍體的過程中會引起結(jié)實率的降低, 同時, 會產(chǎn)生較高頻率的敗育籽粒。本研究獲得的OsMATL編碼區(qū)純合突變體的結(jié)實率均顯著極降低, 首次對粳稻中OsMATL基因進行突變,發(fā)現(xiàn)粳稻OsMATL編碼區(qū)突變后對結(jié)實的影響比秈稻大, 粳稻對OsMATL基因缺失的耐受性較差。先正達公司[11]和王春等[12]對水稻中OsMATL基因進行突變后結(jié)實率為 10%~20%, 并可誘導(dǎo)產(chǎn)生 2%~6%的單倍體。本研究中秈稻突變體的結(jié)實率與前人研究結(jié)果相似, 可推測本研究中秈稻突變體的單倍體誘導(dǎo)率約為5%。本研究在水稻OsMATL突變體后代中發(fā)現(xiàn)了不同程度的敗育籽粒, 前人對玉米[7-9]和小麥[13-14]中MTL基因進行編輯后均伴隨著敗育籽粒的產(chǎn)生, 敗育與單倍體的形成密切相關(guān)[24], 說明MTL基因突變后在不同的作物中均發(fā)生了異常雙受精。OsMATL編碼區(qū)突變體結(jié)實率的降低和敗育籽粒的產(chǎn)生, 暗示本研究在秈稻和粳稻上獲得的OsMATL編碼區(qū)突變體均可能具有單倍體誘導(dǎo)的功能。OsMATL啟動子區(qū)域大片段的缺失對植株結(jié)實并沒有顯著影響, 啟動子大片段缺失的純合突變是否會造成結(jié)實率的降低需要進一步的研究。本研究篩選到不同突變位點和不同遺傳背景的OsMATL突變體,為水稻單倍體誘導(dǎo)系創(chuàng)制提供了基礎(chǔ), 也為進一步研究OsMATL基因的功能提供了材料。
4 結(jié)論
以粳稻日本晴和秈稻華航48為研究材料, 本研究利用CRISPR/Cas9基因編輯技術(shù)獲得OsMATL移碼突變和啟動子大片段缺失的突變體,OsMATL編碼區(qū)突變極顯著降低了結(jié)實率, 但突變效應(yīng)受到不同位點及遺傳背景的影響, 秈稻對于突變的忍受程度高于粳稻。此外,OsMATL啟動子區(qū)域大片段的缺失對植株的結(jié)實幾乎沒有影響, 暗示OsMATL基因表達量的改變與編碼氨基酸的改變調(diào)控雙受精的機制可能不同。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
