太湖花鱉雌雄群體形態及遺傳差異分析
2021-03-15 06:06:41王苗苗梅肖樂馮子偃徐建榮韓曉磊
江蘇農業科學 2021年1期
王苗苗 梅肖樂 馮子偃 徐建榮 韓曉磊


摘要:通過多元統計分析和線粒體DNA測序技術,對中華鱉地方特色品種——太湖花鱉雌雄群體的形態和遺傳差異進行了分析研究。結果顯示,在形態上,對太湖花鱉雌雄群體各項形態參數進行t檢驗差異分析,發現雄性尾部的生長與其體質量的增加相關顯著,呈線性關系;對太湖花鱉雌雄群體形態參數進行主成分分析,并作出主成分1和主成分2的散點圖,得出雌雄群體形態差異不明顯。對太湖花鱉雌雄群體線粒體D-loop控制區序列進行差異分析,雌雄群體間遺傳差異為0.0114;構建N-J聚類系統樹,雌雄群體無單獨聚類,群體遺傳差異不明顯。結果表明,太湖花鱉個體形態差異與其性別存在一定程度的相關性,特別是尾部特征,可用于雌雄性別的初步判斷,但雌雄群體在外觀形態和線粒體DNA上均沒有明顯差異。
關鍵詞:太湖花鱉;多元統計分析;D-loop控制區;形態差異;遺傳差異
中圖分類號:S966.5文獻標志碼:A
文章編號:1002-1302(2021)01-0142-04
作者簡介:王苗苗(1987—),女,江蘇南京人,碩士,水產工程師,主要從事水生生物學研究和水產技術推廣工作。E-mail:miaomiaotgz@163.com。
通信作者:韓曉磊,碩士,高級實驗師,從事淡水水生生物學研究。E-mail:84125241@qq.com。
多元統計分析可根據形態特征的多個參數,對多個互相關聯的指標和對象進行統計分析以揭示其規律,符合農業科學的研究特征[1]。目前,多元統計分析已在水生動物的物種種質評定、種群遺傳變異、性別差異分析等領域開展了大量的研究,然而對龜鱉類物種的研究還鮮有報道[2-6]。線粒體DNA作為細胞中遺傳信息的重要載體,具有長度短、含量高、進化速度快和母性遺傳等特點,特別是D-loop控制區,因其不編碼蛋白,承受選擇壓力較小,是線粒體DNA中進化最快的部分,已成為水生動物種類鑒定、遺傳多樣性、遺傳變異、系統進化、性別差異等方面的研究熱點[7-13]。
太湖花鱉屬于中華鱉(Trionyxsinensis)的地方特色品種,主要分布于長江中下游,太湖流域居多,與普通中華鱉體色差異較大,特別是在自然狀態下體色油綠,背部有對稱小圓黑斑,腹部有明顯塊狀灰黑花斑,由此得名;并因其較高的營養價值和良好的口感風味受到人們青睞,江南地區多有養殖,市場前景廣為看好。太湖花鱉雌雄體在形態規格及生長特性上存在一定差異,本研究通過多元統計分析和線粒體D-loop控制區分析,于表型和基因型多方面對太湖花鱉雌雄特征予以揭示,以期為太湖花鱉雌雄鑒別,以及種質鑒定、資源保護和利用提供一定的理論支持。
1材料與方法
1.1材料來源
太湖花鱉樣品是在2017—2018年取自太湖常州武進段的野生群體,共計取樣30只,其中,雌雄各半,試驗于2019年在長江特色水產工程技術研究中心完成。太湖花鱉活體用于形態參數統計分析,之后取樣品腿部肌肉組織于-70℃冰箱中保存,以備提取DNA使用。
[FK(W11][TPWMM11.tif;S+3mm][FK)]
1.2形態參數測量
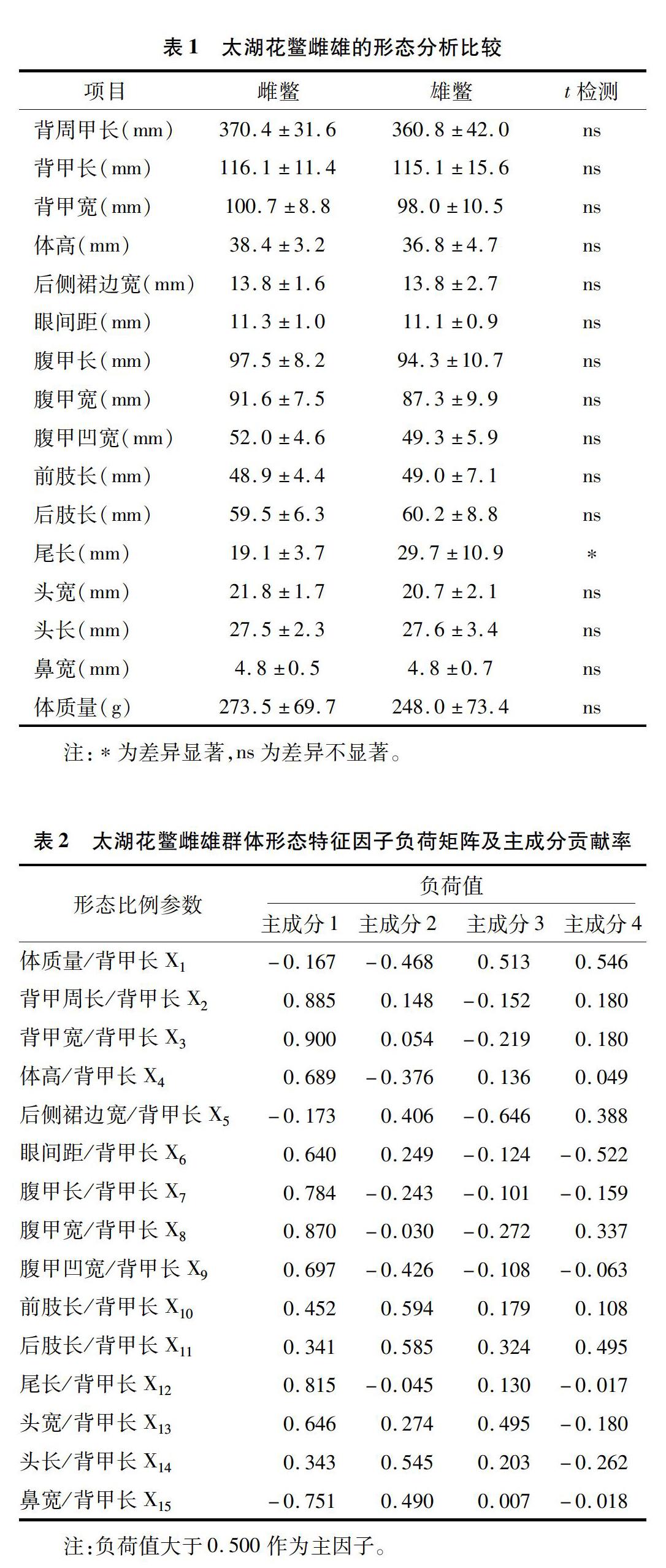
游標卡尺測量每尾太湖花鱉的體高、頭長和頭寬等16項形態指標(精度為0.1mm),電子天平測量每尾太湖花鱉的體質量(精度為0.1g),具體測量方法參考國家標準(GB2104—2007)[14],具體指標見表1、表2。鑒于活體太湖花鱉不便于測量,故可將其放入密閉容器,進行乙醚吸入式麻醉后方可精確測量。
1.3D-loop控制區的PCR擴增及測序
參照韓曉磊等的研究方法[15]提取太湖花鱉樣品基因組DNA并檢測其完整性,測定其濃度后于-20℃保存備用。太湖花鱉2對擴增D-loop控制區的引物序列分別為:T1(5′-3′):[JP9]ATCTTACCCCTACTACACACAT;T2(5′-3′):[JP9]AGACTGGTGATGGAGTATC;T3(5′-3′):[JP9]GCCACAATACTTGTTTATCT;T4(5′-3′):[JP9]AATATCCATCTTGGCGTCTTCA。PCR擴增反應參見陸文浩等的方法[16]具體操作,并測得太湖花鱉樣品線粒體DNA序列。
1.3數據處理
1.3.1形態參數分析
試驗數據采用SPSS19.0軟件進行分析,本研究多元分析方法包括t-檢驗和主成分分析,它們是針對多個研究對象同時進行分析所得數據的運算方法。通過將背甲長作為基數,以取得2個形態參數比值,從而消除鱉體規格大小對多元分析中參數值的影響。
1.3.2D-loop控制區序列分析
所獲得太湖花鱉D-loop控制區序列經過校對處理后,采用軟件ClustalX2.1和MEGA5.1進行序列比對和分析,通過Kimura雙參數模型統計雌雄群體群間遺傳差異,運用鄰接法構建系統發生樹,以bootstrap進行檢驗,重復1000次。
2結果與分析
2.1形態差異分析
由表1可知,太湖花鱉雄鱉體質量與尾長關系的r=0.662(r>r0.01),尾部生長與體質量增加相關性顯著,其一元線性回歸方程為:Y體質量=44.473X尾長+115.837,回歸顯著性檢驗F=10.143,F>F0.01;太湖花鱉雌鱉體質量與尾長關系的r=0.445(r 2.2主成分分析 由表2可知,太湖花鱉雌雄群體方差貢獻率較高的4個主成分貢獻率分別為43.163%、14.427%、8.812%和8.600%,其累積貢獻率為75.003%,包含了群體總變異的大部分,可見以上4個相互獨立的主成分可代表太湖花鱉雌雄群體間的形態差異。在主成分1中,變量X2、X3、X4、X6、X7、X8、X9、X12和X13影響最大;在主成分2中,變量X10、X11和X14影響最大;在主成分3和主成分4中,變量X2影響最大。背甲周長、背甲寬、腹甲長、腹甲寬和尾長是主成分1變異貢獻的主要形態指標,可見太湖花鱉鱉體長度、寬度和尾長是引起其雌雄群體形態差異的主要指標。 由圖2可知,太湖花鱉雌雄群體主成分1和主成分2為相對值,雌雄群體之間存在部分重疊,主成分1和主成分2不能準確將雌雄群體進行區分,故太湖花鱉雌雄群體總體形態差別較小,形態差異統計分析較難將其準確鑒別。 2.3D-loop控制區序列分析 2.3.1太湖花鱉線粒體D-loop控制區序列比較 測序所得序列與GenBank中序列號為AY687385.1的中華鱉線粒體DNA序列進行Blast同源性比對,證實所測序列位于第15321位到15835位之間,大小為515bp,堿基組成分別為G、C、T和A的堿基平均含量分別為10.0%、30.0%、32.9%和27.1%,其中T+A含量(60.0%)高于G+C含量(40.0%),是線粒體DNA典型的反G偏倚特征。太湖花鱉雌雄群體的群內遺傳相似度為99.23%。 2.3.2太湖花鱉分類地位鑒定 根據D-loop控制區序列差異,用MGEA5.1分析2個群體的遺傳距離,由圖3可知,發現雌雄群體間遺傳差異為0.0114。構建聚類系統發生樹,太湖花鱉雌雄群體混為一體,群內個體無單獨聚類。 3討論 t檢驗是用于小樣本(樣本容量小于30)的2個平均值差異程度的檢驗方法,它是用T分布理論來推斷差異發生的概率,從而判定2個平均數的差異是否顯著[17-18]。本研究中,太湖花鱉雄性尾巴的生長與其體質量的增加顯著相關,可用于太湖花鱉雌雄性別的鑒別;主成分分析中尾長也是引起形態差異的主成分變量之一,且需與其他形態變量共同作用進行差異分析。珠水對中華鱉性別鑒別方法進行了研究,指出雄性中華鱉較之雌性尾巴較長,大多能自然伸出裙邊外,故尾部特征可以作為雌雄判別的重要依據[19],此結論與本研究結果基本一致。然而,在太湖花鱉雌雄群體30個樣本的散點圖分析結果中,太湖花鱉雌雄群體并沒有明顯區分。以上推斷表明,太湖花鱉個體形態差異與其性別存在一定程度的相關性,特別是尾部特征,可用于雌雄性別的初步判斷,但其群體的形態差異并不明顯,還需更多的參數乃至更深入的分析研究予以揭示。 本研究通過線粒體D-loop控制區序列比對差異分析,發現太湖花鱉雌雄群體群間遺傳差異僅為0.0114,可以認為2個群體間不存在有效遺傳差異。太湖花鱉雌雄群體聚類系統發生樹中2個群體的群內個體未按照性別差異單獨聚類,同樣揭示雌雄群體群間遺傳差異并不明顯。 綜上所述,太湖花鱉雌雄群體在外觀形態上可通過尾巴長短進行簡單區分,但進行準確的分辨還相對較難,線粒體DNA差異同樣不明顯。由此推斷,太湖花鱉雌雄群體差異在外觀形態只有微小的體現,而在水產養殖中,雄鱉生長速度快于雌鱉,成熟的雄性個體比雌性大且一般為雌性的3倍左右[17],揭示太湖花鱉雌雄差異可能主要體現在生長速度上,故體質量的變化可作為研究方向。 參考文獻: [1]王立賓.多元統計分析:模型、案例及SPSS應用[M].北京:經濟科學出版社,2010:89-91. [2]GolubtsovAS,BerendzenPB.Morphologicalevidencefortheoccurrenceoftwoelectriccatfish(Matapterurus)speciesintheWhiteNileandOmo-Turkanasystems(EastAfrica)[J].JFishBio,1999,55(3):492-505. [3]GibsonAR,BakerAJ,MoeedP.Morphometricvariationinintroducedpopulationsofthecommonmyna(Acridotherestristis):Anapplicationofthejaekknifetopfincipalcomponentanalysis[J].SystematicZoology,1984,33(4):408-421. [4]VossRS,MarcusLF,PatriciaEP.MorphologicalevolutioninmuroidrodentsⅠ.conservativepatternsofcraniometriccovarianceandtheirontogeneticbasisintheneotropicalgenuszygodontomys[J].Evolution;InternationalJournalofOrganicEvolution,1990,44(6):1568-1587. [5]SomersKM.Multivariateallometryandremovealofsizewithprincipalcomponentsanalysis[J].SystematicZoology,1986,35(3):359-368. [6]潘德博,陳昆慈,朱新平,等.黃喉擬水龜(♀)與三線閉殼龜([XZ(20#]♂[XZ)])雜交后代的形態特征及其與父母本的比較研究[J].水生生物學報,2009,33(4):620-626. [7]陳大慶,張春霖,魯成,等.青海湖裸鯉繁殖群體線粒體基因組D-loop區序列多態性[J].中國水產科學,2006,13(5):800-806. [8]肖武漢,張亞平.魚類線粒體DNA的遺傳與進化[J].水生生物學報,2000,24(4):384-391. [9]譚圍.孟加拉笛鯛和四帶笛鯛線粒體基因組特征及分子進化研究[D].湛江:廣東海洋大學,2009. [10]董曉麗.半滑舌鰨(Cynoglossussemilaevis)線粒體基因組全序列和性別相關基因的克隆與表達分析[D].青島:中國海洋大學,2010. [11]李勝杰,白俊杰,葉星,等.基于線粒體D-loop區探討我國養殖大口黑鱸的分類地位和遺傳變異[J].海洋漁業,2008,30(4):291-296. [12]盛軍慶,林巧惠,王軍花,等.池蝶蚌線粒體基因組雙單性遺傳現象分析[J].動物學雜志,2014,49(4):597-604. [13]薛婷.淡水珍珠蚌DUI發生及性腺發育研究[D].上海:上海海洋大學,2016. [14]張軒杰,劉筠,周工健,等.中華鱉國家標準:GB21044—2007[S].北京:中國標準出版社,2007. [15]韓曉磊,徐建榮,李小蕊,等.鳡魚群體遺傳多樣性的AFLP分析[J].南京師大學報(自然科學版),2009,32(1):110-114. [16]陸文浩,韓曉磊,陳梁,等.不同群體中華鱉的形態多樣性分析[J].水產科學,2017,36(6):784-788. [17]秦秋燕.基于動物覓食原理的改進微粒群算法研究[D].太原:太原科技大學,2010. [18]李艷.數據分析軟件SAS實用教程[M].武漢:武漢大學出版社,2015:299-301. [19]珠水.中華鱉雌雄鑒別簡易方法[J].現代漁業信息,2004,19(11):33.
