豬MAP2K6 基因多態性及其與經濟性狀的關聯分析
2021-03-15 05:40:02張佳麗聶光偉
中國畜牧雜志 2021年3期
張佳麗,聶光偉,劉 軍,左 波
(農業部豬遺傳育種重點實驗室,華中農業大學動物科技學院,湖北武漢 430070)
分子標記輔助選擇(Marker Assistant Selection,MAS)是指在基因克隆或定位的基礎上,通過分析與目標基因緊密連鎖的分子標記,在群體中對含有目標性狀的個體進行選擇[1-2]。對含有目標性狀的個體進行快速、準確的選擇,而且不受外界環境的影響,這在一定程度上解決了育種過程中存在的盲目性、效率低的問題,大大加快了育種進程[3],促進了分子標記輔助選擇和基因組選擇在豬生產上的應用。豬MAP2K6基因位于12 號染色體,編碼絲裂原活化蛋白激酶激酶6(Mitogen-Activated Protein Kinase Kinase 6,MAP2K6)。MAP2K6基因屬于MAPK家族的一員,目前已鑒定出4 種MAPK家族成員,分別是細胞外信號調節激酶1/2(Extracellular Signal-regulated Kinase 1/2,ERK1/2)、ERK(Extracellular Regulated Protein Kinases,ERK)、p38 絲裂原活化蛋白激酶(p38 Mitogenactivated Protein Kinase,p38MAPK)和c-Jun 氨基末端激酶(c-Jun N-terminal Kinase,JNK)[4],分別介導4 條并行的MAPK信號通路。其家族成員主要參與調控多種細胞的關鍵信號級聯反應,能被生長因子、促細胞分裂素、細胞因子及各種形式的環境應激因素所激活,而MAP2K6基因的激活是由于對絲氨酸和蘇氨酸殘基進行雙重磷酸化引起的[5]。p38是MAPK超家族的另一成員,MAP2K6基因是其上游的一個特異激活因子[6],二者之間具有一定的相互作用,Fujishiro 等[7]研究表明,MAP2K6和p38MAPK信號通路的激活,能調節葡萄糖轉運蛋白的表達且對胰島素信號通路具有抑制作用。根據Meng 等[8]的報道,MAP2K6及PLCB1基因共同參與促性腺激素信號通路的傳導,對動物個體的生長發育具有重要作用。
在實際生產中,對QTLs 的定位并進行分子標記是一種行之有效的重要育種手段。如Tang 等[9]通過鑒定CNN3基因的SNP 位點初步確定了該基因與仔豬斷奶重、出生重等生長性狀有關。MAP2K6基因通過調節PPARγ及CEBPβ基因的表達,進而激活p38MAPK,促進脂肪的形成[10],證明MAP2K6基因與牛胴體性狀具有遺傳關聯。Meng 等[8]在大白豬群體中通過GWAS方法鑒定與其生長性狀相關的候選基因,結果表明在10 905 724 bp 位置處存在1 個候選基因MAP2K6,與豬的平均日增重顯著相關。因此,在本研究中將MAP2K6作為豬經濟性狀的候選基因。基于MAP2K6基因的生物學功能和QTL 定位結果,本實驗鑒定了法系大白豬群體中SNPs 位點并分析了豬的重要經濟性狀,期望能夠發現可用于育種實踐的DNA 分子標記,為豬分子育種提供理論依據。
1 材料與方法
1.1 實驗豬群及性狀記錄 采集384 頭法系大白豬前腔靜脈血液,使用血液基因組DNA 中量提取試劑盒(離心柱型)提取樣品DNA,并于-80℃超低溫冰箱中保存。測定的表型性狀及數據收集包括達100 kg 體重日齡(d)、背膘厚(mm)、初生重(kg)、體長(cm)、眼肌面積(cm2)、左乳頭數和右乳頭數。本研究用B 型超聲設備測定豬背膘厚和眼肌面積(背膘厚及眼肌面積測定位置在豬倒數第3、4 根肋骨距離背中線4~5 cm 處),測量數據由育種軟件GBS5.0 校正后得到。
1.2 法系大白豬4 個SNPs 的鑒定與分析 在NCBI 數據庫中查找目的基因DNA 序列(XM_021068116.1),使用Premier 5.0 軟件設計引物,并由上海生工生物工程技術有限公司合成。擴增各基因所用引物及目的片段長度如表1 所示。PCR 擴增反應總體系為20.0 μL:DNA模板1 μL,2×Taq PCR Mix 10.0 μL,ddH2O 8.0 μL,正、反向引物(10 μmol/L)各0.5 μL。PCR 反應程序:95℃預變性5 min;95℃變性40 s,退火30 s,72℃延伸,共36 個循環;最后72℃延伸5 min。PCR 產物于瓊脂糖凝膠電泳檢測,以5 μL 的DNA Marker DL2000作為參照。電泳結束后利用凝膠成像系統觀察擴增結果。
1.3 SNP 質譜分型 本實驗室委托北京康普森生物技術有限公司對MAP2K6基因的4 個突變位點rs345304630(SNP1)、rs325278117(SNP2)、rs332017877(SNP3)和325752048(SNP4)采用質譜分型,檢測法系大白豬DNA 樣本。實驗的主要流程:DNA 提取、質檢;DNA 濃度調試;引物設計、溶解、稀釋;PCR 擴增;PCR 產物堿性磷酸酶處理;單堿基延伸;樹脂純化;芯片點樣;質譜檢測;統計基因型。

表1 MAP2K6 引物序列
1.4 數據分析 采用SAS 統計軟件中的混合線性模型(Mixed),以GLM 程序進行基因型和性狀表型值之間的關聯分析及顯著性檢驗,結果以平均值±標準誤表示(不同基因型表型數值間P<0.01 判定為差異極顯著,P<0.05 判定為差異顯著;若P>0.05 則不顯著)。應用Excel 軟件分別計算基因型頻率和等位基因頻率,并進行χ2檢驗,分析其是否符合哈代-溫伯格平衡(P>0.05表示處于遺傳平衡狀態,P<0.05 則相反)。生長性狀分析模型:
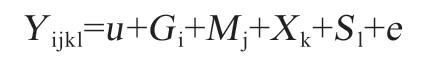
式中,Yijkl為性狀觀察值;u為性狀總平均值;Gi為基因型效應(包括基因加性效應和顯性效應);Mj為年-季度效應;Xk為性別效應及固定效應;Sl為父本效應及隨機效應;e為隨機誤差,假定服從(0,χ2)分布。
2 結果
2.1MAP2K6基因多態性檢測 測序結果表明,在法系大白豬MAP2K6基因內含子8、內含子11 和外顯子12區域共鑒定出4 個SNPs,包括SNP1(rs 345304630)、SNP2(rs 325278117)、SNP3(rs 332017877)和SNP4(rs 325752048)。如表2 所示,采用飛行時間質譜的方法對MAP2K6基因4 個SNPs 位點進行基因型分析,均檢測到3 種基因型。
對MAP2K6基因多態性位點不同基因型與法系大白豬的經濟性狀進行相關性分析(表3),結果發現,法系大白豬的SNP1 與達100 kg 體重日齡和右乳頭數性狀存在顯著相關性,AA 基因型個體達100 kg 體重日齡顯著低于AG 基因型個體,說明AA 基因型個體具有較快的生長速度;GG 基因型個體右乳頭數顯著高于AG 基因型個體;SNP2 與達100 kg 體重日齡和右乳頭數性狀存在顯著相關性。AA 基因型個體達100 kg 體重日齡顯著低于AG 基因型個體,GG 基因型個體具有較快的生長速度,AA 基因型個體右乳頭數顯著高于AG基因型個體;SNP3 與達100 kg 體重日齡和右乳頭數性狀存在顯著相關性,AA 基因型個體達100 kg 體重日齡顯著低于AG 基因型個體,說明AA 基因型個體具有較快的生長速度,GG 基因型個體的右乳頭數顯著高于AG 基因型個體;SNP4 與眼肌面積存在顯著的相關性,AG 基因型個體眼肌面積顯著低于AA 基因型個體。
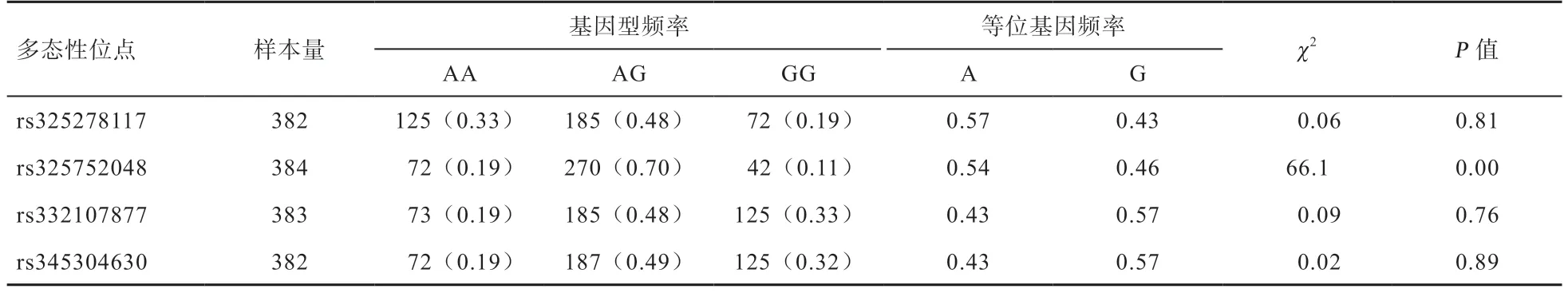
表2 MAP2K6 基因SNPs 基因頻率和等位基因頻率
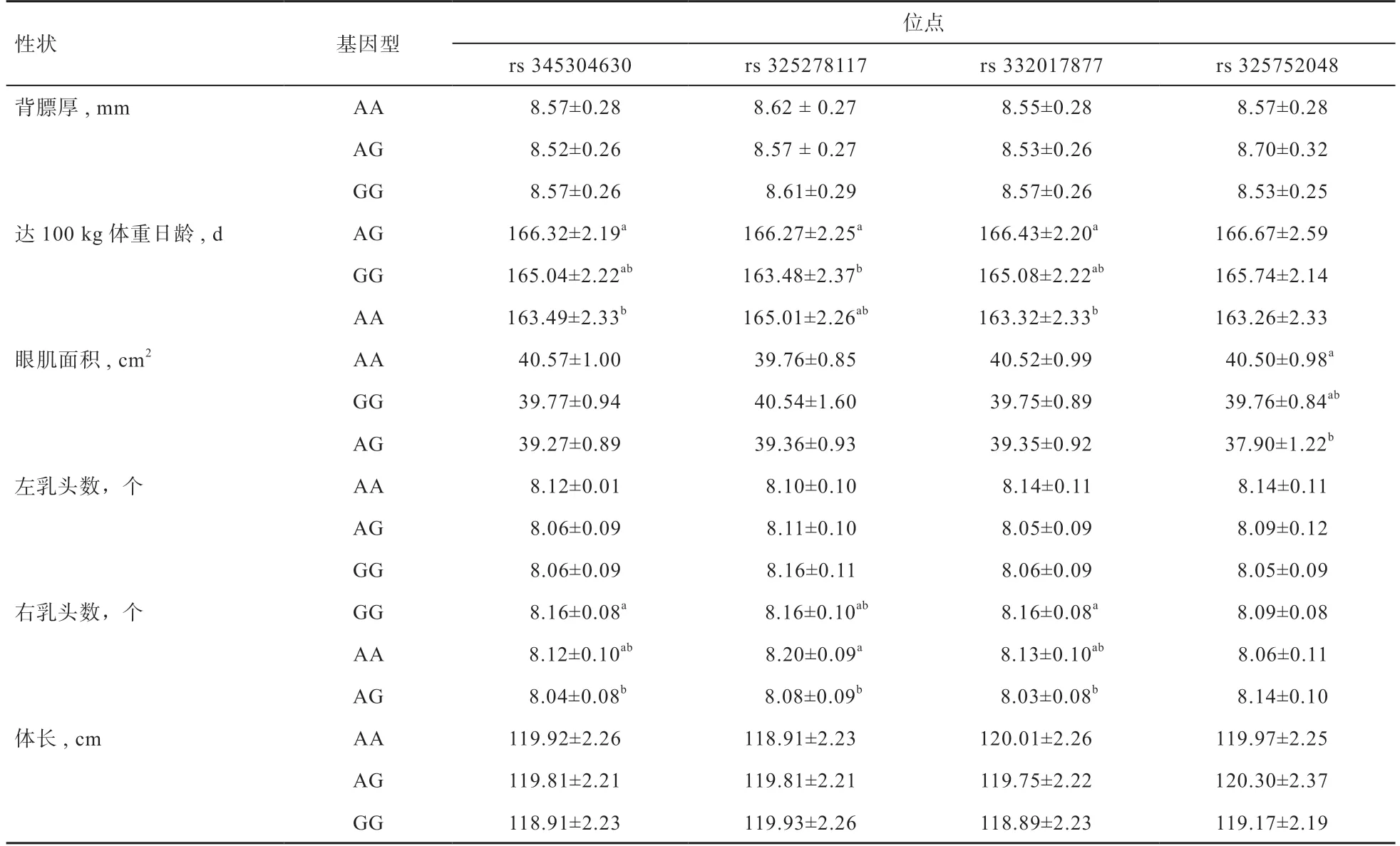
表3 MAP2K6 基因多態性與法系大白豬經濟性狀的關聯分析
2.2 法系大白豬MAP2K6基因3 個SNPs 連鎖不平衡及單倍型分析 如圖1 所示,SNP1(rs345304630)和SNP2(rs325278117)、SNP1(rs345304630)和SNP3(rs332017877)在法系大白豬中處于完全LD(D'=1和r2=1),SNP2(rs325278117)和SNP3(rs332017877)處于強LD(D'=1 和r2=0.99)。如表4 所示,MAP2K6基因3 個SNPs 在法系大白豬中形成1 個單倍型模塊,共產生2 種單倍型,單倍型H2(AGA)的頻率均低于單倍型H1(GAG)。

圖1 法系大白豬MAP2K6 基因SNPs 連鎖不平衡區
2.3 法系大白豬MAP2K6基因單倍型組合與生長育肥和乳頭數性狀關聯分析 由表5 可知,MAP2K6基因在法系大白豬群體中共產生3 種單倍型組合,單倍型組合H1-H1、H1-H2、H2-H2 與體長、乳頭數、達100 kg 體重日齡、背膘厚、初生重和眼肌面積性狀有顯著關聯性。

表4 法系大白豬MAP2K6 基因單倍型頻率
2.4MAP2K6在豬各組織中差異表達分析 由圖2 可知,MAP2K6基因在法系大白豬13 個組織中表達量不同,其中在脾和脂肪中表達量最高,在肝、心、腿肌、腎、舌中的表達量沒有顯著性差異。
3 討 論
已有研究表明,在杜洛克豬中鑒定出MAP2K6與豬的繁殖性狀有關[11],但并沒有做進一步研究。本研究首次在法系大白豬MAP2K6基因上發現4 個SNP 位點,與達100 kg 體重日齡、右乳頭數和眼肌面積有顯著相關;MAP2K6基因在豬各組織中的表達譜結果說明,MAP2K6在脂肪中高表達。MAP2K6在脂肪細胞和葡萄糖轉運蛋白表達中起重要作用[5,7,12],該基因調控機體能量轉化,參與GnRH 信號通路[12],GnRH 信號傳導在哺乳動物青春期開始時的生長中也起著重要作用[13-18]。在其他動物中,Ryu 等[19]研究表明MAP2K6基因與牛的胴體性狀(胴體重、背膘厚和大理石紋評分)具有相關性,且在該基因第1 和第11 內含子上鑒定到4 個單核苷酸多態性位點,其中c.17-10118T>G 多態性位點顯著影響牛的胴體重和大理石紋評分。豬MAP2K6可能是通過影響其胴體性狀、繁殖性狀和生長性狀從而影響脂肪的形成,這還需要進一步研究。另外,本研究出現這種結果可能是由于生存環境、遺傳背景、種質特征等因素的不同,導致該基因的轉錄因子調控差異,進而影響MAP2K6的mRNA 表達水平。MAP2K6基因可能通過調控動物個體的能量利用水平、激素水平等影響其生長發育。因此該基因可作為研究豬生長性狀的一個候選基因。目前,僅有一篇報道表明MAP2K6基因與豬的平均日增重具有相關性[8]。因此,對豬MAP2K6基因功能和SNP 的關聯仍然需要進一步研究。
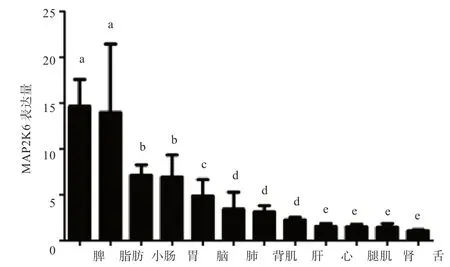
圖2 MAP2K6 基因在法系大白豬不同組織中表達差異
在本研究中,多態性位點rs345304630、rs325278117和rs332017877 均處于遺傳平衡狀態,并未受到遺傳漂變等外界因素的影響,這些多態性位點在群體中可以穩定遺傳。豬生長育肥和乳頭數性狀是受到多基因調控的復雜性狀,單個SNP 不足以闡明基因與性狀間的因果關系[20]。在本研究中,法系大白豬MAP2K6基因3 個SNPs(rs345304630、rs325278117 和rs332017877)處于強連鎖不平衡,且3 個SNPs 形成一個緊密連鎖的整體。單倍型組合與豬生產育肥和乳頭數性狀并無顯著關聯性,推測可能由于實驗樣本量較少導致,下一步將擴大樣本量檢測MAP2K6基因在不同品種豬群的多態性,為分子標記輔助選擇和全基因組選擇提供理論依據和標記資源。
4 結 論
本研究中,MAP2K6基因多態性位點SNP1(rs345304630)、SNP2(rs325278117)和SNP3(rs332017877)與法系大白豬達100 kg 體重日齡和右乳頭數顯著相關,SNP4與眼肌面積顯著相關,其中SNP1(rs345304630)和SNP3(rs332017877)多態性位點的AA 基因型個體生長速度明顯高于AG 和GG 基因型個體,在實際生產中,可以選擇AA 基因型個體的母豬來縮短達100 kg 體重日齡,提高經濟效率;SNP1(rs345304630)、SNP2(rs325278117)和SNP3(rs332017877)多態性位點的GG 基因型個體乳頭數顯著高于AA 和AG 基因型個體,通過選擇GG 基因型個體來增加母豬的泌乳和帶仔能力,對提高哺乳仔豬的存活率和斷奶重具有重大意義。
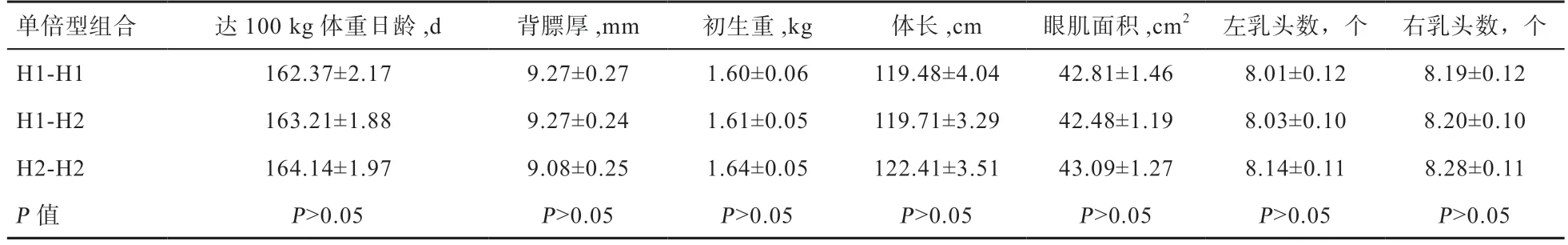
表5 法系大白豬MAP2K6 基因二倍型與生長發育及乳頭數性狀的相關性分析
