骨形態發生蛋白-2(BMP2)基因的生理功能和信號通路研究進展
2021-03-15 05:40:02費曉娟金美林盧曾奎魏彩虹
中國畜牧雜志 2021年3期
關鍵詞:小鼠
費曉娟,金美林,盧曾奎,狄 冉,魏彩虹*
(1.中國農業科學院北京畜牧獸醫研究所,北京 100193;2.中國農業科學院 蘭州畜牧與獸藥研究所,甘肅蘭州 730050)
綿羊是重要的家畜之一,擁有11 000 年的馴化史,是史上第一個被馴化的游牧動物。根據綿羊尾型可分為短瘦尾羊、長瘦尾羊、短脂尾羊、長脂尾羊和肥臀羊五類[1]。肥尾羊大約在5 000 年前從瘦尾羊中演化而來,目前肥尾羊數量已占全世界總羊數的四分之一[2]。本實驗室前期的研究表明,在哈薩克羊(肥尾羊)和藏羊(瘦尾羊)脂肪組織中有464 個基因差異表達,其中BMP2基因與綿羊脂尾的發育有關[3-5]。進一步研究表明,BMP2基因能夠促進綿羊前體脂肪細胞的分化,參與綿羊尾部脂肪沉積,從而影響綿羊尾型發育[6]。
1965 年 Urist 在成人的脫鈣骨基質中提取出一種能誘導異位骨發生的活性蛋白,并根據其生物學特性命名為骨形態發生蛋白(Bone Morphogenetic Protein,BMPs),屬于轉化生長因子β(The Transforming Growth Factor-β,TGF-β)超家族[7]。BMPs 家族成員至少有40 個,目前關于骨代謝的研究最多[8]。BMPs 家族被分為5 個亞型,其中,BMP2與骨的生長和分化相關,屬于第一類亞型[9-10]。BMPs 是一種參與生物機體機能發生和調節的分泌蛋白,與細胞增殖和分化、細胞凋亡、形態發生、器官形成等相關[11],BMP2同樣具有廣泛的生物學功能[12-17]。本文就BMP2基因的結構和功能及其參與的信號通路等進行綜述,為進一步研究BMP2基因提供理論依據。
1 BMP2 基因的發現和結構
1988 年Wozney 等[18]首次克隆出了人的BMP2基因,由3 個外顯子和2 個內含子組成,且該基因翻譯起始于第2 外顯子結束于第3 外顯子[19]。研究表明,不同物種的BMP2基因的編碼區序列同源性較高[20]。采用Needle 工具(https://www.ebi.ac.uk/Tools/psa/emboss_needle/)比較不同物種BMP2基因編碼區核苷酸的同源性(表1),綿羊與牛的BMP2編碼區核苷酸的同源性最高;山羊與豬、牛和雞的BMP2編碼區核苷酸序列同源性較高,均在50% 以上;綿羊與雞的BMP2編碼區核苷酸序列同源性最低。
BMP2 蛋白屬于疏水性蛋白,是由信號肽、前體蛋白和成熟結構域組成的較大前體蛋白。蛋白分泌后去除信號肽,前體蛋白發生二聚,然后特異性蛋白水解酶將二聚后的前蛋白裂解,釋放成熟區域。成熟區域可以與自身或者其他BMP 蛋白二聚,生成具有生物活性的二聚成熟蛋白,從而發揮生物學功能[21]。成熟的BMP2蛋白是由2 個114 肽亞單位通過二硫鍵共價連接的同二聚體[22-23]。在BMP2 蛋白的羧基末端有一段由 7 個半胱氨酸殘基形成的保守區域,其中 6 個半胱氨酸形成了3 個分子內二硫鍵,第7 個半胱氨酸殘基通過二硫鍵發揮二聚作用[23]。此外,BMP2單體的核心是一個半胱氨酸結,由8 反向平行的β-折疊和一個α-螺旋形成,且每個單體只有2 個色氨酸殘基,在骨的修復和誘導過程中發揮重要的作用[10-11]。
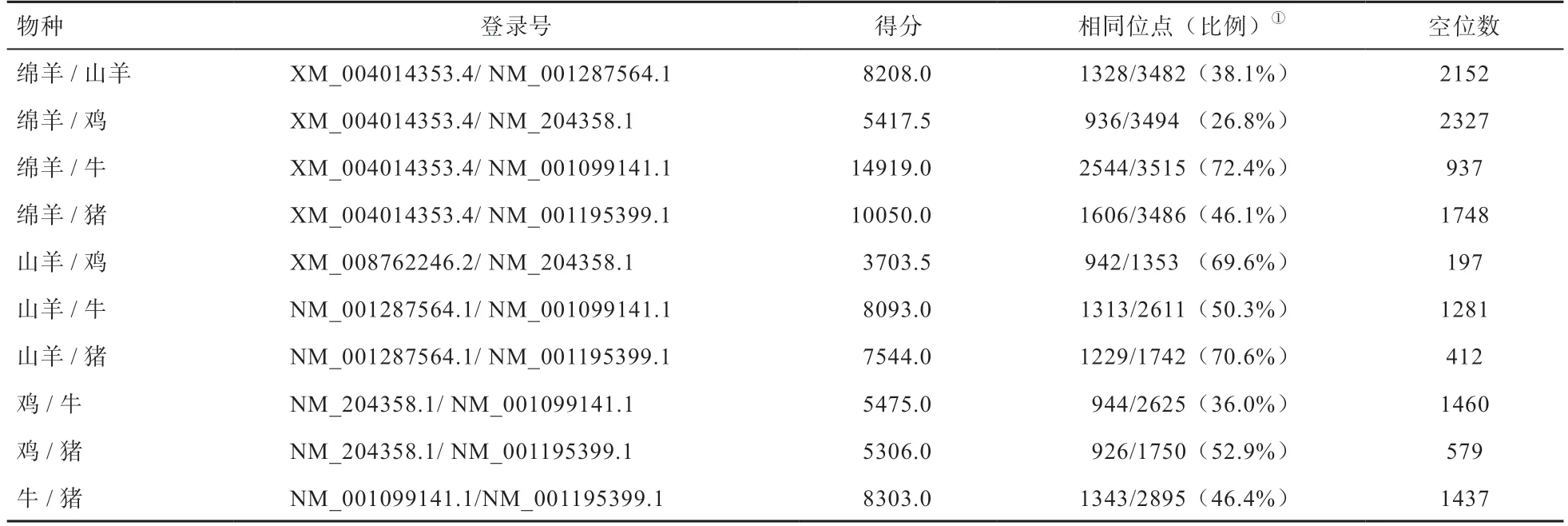
表1 不同物種之間BMP2 基因編碼區核苷酸序列的比較
2 BMP2 基因的表達
BMP2基因在人和動物的不同器官中均有不同程度表達[24-25]。小鼠BMP2基因在胚胎期的表達模式為胚胎外區域在第6 天[28]、羊膜的中胚層細胞在第8.5 天[11]、背部神經管的外胚層在第8.5~9.5 天[11]、心臟的外心肌層和脊椎頂部的外胚層在第9.5~10.5 天[11]、眼睛、牙齒和胡須在第12.5 天[11]表達。與正常組織相比,BMP2在癌細胞中高表達,可作為人類癌癥、腫瘤等疾病的檢測標志物[26]。BMP2基因同樣在健康組織中表達,如在鯉魚的鰓、腸、肝、脾、腦和血液中都有較高的表達水平,與鰭的發育或再生相關[27]。
3 BMP2 的生理功能
BMPs家族通過TGF-β超家族的I 型受體和II 型受體傳遞信號發揮生物學功能,目前在哺乳動物中發現TGF-β超家族有7 種Ⅰ型受體和 5 種Ⅱ型受體,且均具有絲蘇氨酸激酶活性[29]。BMPs 家族受體由一個短的胞外結構域、一個單一的跨膜結構域和一個具有活性絲氨酸/蘇氨酸區域的胞內結構域三部分構成[23]。BMPs家族受體包括3 種Ⅰ型受體BMPIA型受體、BMPIB型受體和激活素受體(如kinase、ACVRI 和 ALK2 等);3 種Ⅱ型受體BMPRII、ActRIIA 和ActRIIB。BMPs 蛋白分泌后,BMPs 配體與II 型受體結合并磷酸化I 型受體,隨后再磷酸化下游轉錄因子,形成復合物并轉移到細胞核,啟動相關基因轉錄,從而調節細胞的增殖和分化等生命活動[30-31]。
3.1BMP2與骨形成BMP2屬于特異成骨基因,現已被美國食品和藥物管理局(Food and Drug Administration,FDA)批準用于脊椎融合的臨床治療中。BMP2在骨形成的過程中,受其他因子影響。Chio 等[32]在大鼠骨質疏松癥體內模型中觀察到維生素C 使BMP2的表達增強,抑制大鼠骨質疏松癥的發生。此外,Kovermann 等[17]研究表明,含有BMP2的體外培養基能夠誘導人滑膜源性干細胞形成軟骨,在含有TGF-β1 和BMP2的聯合培養基中與軟骨形成相關的聚蛋白多糖(Aggrecan,ACAN)的表達也顯著上調,更加促進軟骨形成。此外,BMP2/7異二聚體蛋白聯合低濃度的全反式維甲酸(All-Trans Retinoic-Acid,ATRA)可使小鼠骨髓源巨噬細胞中破骨細胞生成相關基因的表達下調,阻斷破骨細胞生成并降低破骨細胞的吸收活性[33]。綜上,這些因子與BMP2之間形成調控骨發生的網絡,為骨疾病的治療提供新的見解。
3.2BMP2與脂肪沉積 將人的骨髓源性干細胞提取、分離并在三維成骨或成脂培養基中培養,結果顯示在補充了BMP2的三維培養基中形成的脂滴明顯多于其他培養基[34]。對547 例不同體重指數個體的內臟和皮下組織檢測,發現在肥胖人群中內臟組織和皮下組織中BMP2的表達顯著高于健康人群[35]。在體外研究大鼠的肌腱干細胞中脂肪生成機制的過程中,BMP2可磷酸化并激活下游的環磷腺苷效應元件結合蛋白(cAMPresponse Element Binding Protein,CREB)和Smad 蛋白,隨后上調過氧化物酶體增殖物激活受體γ2(Peroxisome Proliferator-Activated Receptorγ2,PPARγ2)的表達,從而增強肌腱干細胞脂肪分化的趨勢[36]。這些結果與Birgit 等[37]的研究結果一致,均表明BMP2與組織中脂肪沉積相關。
本課題組使用Illumina Ovine SNP50 芯片,對10個本土綿羊品種進行群體結構分析和基因組選擇分析,發現這些地方品種分為肥尾和瘦尾2 種類型;用FST和hapFLK 方法分析發現BMP2基因與綿羊尾型的發育有關[4-5],并確定BMP2可作為綿羊尾型發育的關鍵候選基因[38]。此外,有報道稱BMP2可以促進豬前體脂肪的分化[14]。本課題通過構建慢病毒載體,將BMP2基因過表達于綿羊前體脂肪細胞中,發現BMP2基因在mRNA 水平過氧化物酶體增殖物激活受體γ(Peroxisome Proliferator-Activated Receptorγ,PPARγ)和脂蛋白脂酶(Lipoprteinlipase,LPL)均上調,但是蛋白表達水平差異不顯著,推斷BMP2同樣能夠促進綿羊前體脂肪細胞的分化,參與綿羊尾部脂肪沉積,影響綿羊尾型發育。但是其作用機制尚不清楚,需要進一步研究[6]。
3.3BMP2與生長發育BMP2是細胞分化和增殖過程中重要的介質,BMP2的表達水平與細胞的增殖和凋亡息息相關。以腸缺血再灌注大鼠為模型,分別在24 h和48 h 處死,發現灌注大鼠的空腸中BMP2基因和蛋白的表達均顯著上升,且空腸和回腸的細胞數量增加[39]。同時BMP2通過不同途徑促進小鼠星形膠質細胞分化,小鼠大腦皮層星形膠質細胞經過BMP2處理后,激活了轉錄共激活因子相關蛋白(Yes-associated Protein,YAP),并與Smad1/5/8 蛋白相互作用并穩定Smad1/5/8 蛋白的表達,從而促進小鼠大腦皮層星形膠質細胞的分化[39];在小鼠神經干細胞中發生DNA 損傷的模型中,BMP2通過激活Janus 激酶/信號轉導與轉錄激活子(The Janus kinase/signal transducer and activator of tran-ions,JAK-STAT)信號通路,上調星形細胞標記物膠質纖維酸性蛋白(Glial Fibrillary Acidic Protein,GFAP)的表達,同樣促進衰老細胞的星形膠質細胞的分化[40]。此外,當過表達miR-378 時可激活BMP2基因參與的BMP-Smad 信號通路,促進綿羊成肌細胞的增殖[41]。
BMP2還對胚胎發育過程中器官的形成產生影響,對胚胎發育是否畸形起決定性作用。用逆轉錄病毒RASC-BMP2感染雞胚中腸,發現在雞胚腸道形成更緊密的卷曲環,每個循環管的長度和半徑顯著減少;腸系膜背側的細胞密度增加,并且其面積明顯減小[42]。在小鼠胚胎發育過程中,BMP2殘缺會導致前羊膜腔不能閉合,形成畸形羊膜,同時也會造成小鼠心臟發育缺陷[43]。
3.4BMP2與癌癥 近年來的研究表明,BMP2既可以促進癌癥的發生又可以抑制癌癥的發生,這可能是由于細胞種類或者細胞所處的環境不同,從而出現這一矛盾的結果。在乳腺癌細胞系MCF-7 中發現,BMP2基因啟動子甲基化使BMP2表達下調,增強了乳腺癌細胞的耐藥性,促進癌癥發生[44]。而將小鼠和人的乳腺癌細胞MCF-7 分別用BMP2處理,結果顯示BMP2抑制癌細胞的遷移和增殖[45-46]。在鼻咽癌中,sox9與BMP2啟動子結合,增強了BMP2表達,進而激活了BMP2誘導的mTOR 信號轉導,促進鼻咽癌中癌細胞的增殖、遷移和侵襲[47-48]。在小鼠慢性胰腺癌細胞發現BMP2可增強miR-200 的表達,抑制慢性胰腺癌細胞纖維化[49]。此外,激活素A 直接抑制BMP2誘導的多發性骨髓瘤細胞系的抗增殖活性,通過設計BMP2變異體,發現該變異體對特定的II 型受體表現出增強的受體結合親和力,進而促進骨生長,這為多發性骨髓瘤的治療提供新的思路[50]。
4 BMP2 參與的調控通路
4.1BMP2與PI3K/Akt 信號通路 磷脂酰肌醇3-蛋白激酶(PI3K/AKT)信號通路調節細胞的生長發育,同時介導多種癌癥的發生。在含有胎牛血清的培養基中培養小鼠前骨細胞,發現BMP2只需要磷酸化Akt位點Ser473,就可以促進成骨細胞分化;在不含胎牛血清的培養基中,BMP2需要磷酸化Ser473 位點并使Akt的Thr308 位點去磷酸化來增加成骨細胞的分化[51]。在胰腺癌細胞系panc-1 和胃癌細胞中,BMP-2刺激上皮間質轉化(Epithelial-Mesenchymal Transition,EMT)的發生,同樣使PIK3下游因子Akt發生磷酸化,從而促進癌細胞的運動和侵襲[52]。可見,BMP2對Akt的磷酸化,是激活PI3K/Akt 通路的關鍵。此外,在無血清培養基中培養小鼠軟骨細胞系N1511,并加入50 ng/mLBMP2,發現BMP2能夠通過PI3K/Akt 抑制細胞的凋亡,深入研究發現BMP2通過激活PIK3信號,抑制半胱氨酸天冬氨酸蛋白酶-9(Caspase-9)和半胱氨酸天冬氨酸蛋白酶-3(Caspase-3)的表達[53]。
4.2BMP2與Smad 信號通路 Smad 信號通路是BMP2的經典通路,BMP2蛋白分泌后,配體與II 型受體結合并激活I 型受體的絲蘇氨酸激酶活性位點,隨后啟動Smad 家族蛋白質的信號級聯。具體來說是Smad1/5/8被BMP2受體復合物磷酸化,然后與Smad4 結合形成Smad1/5/8Smad4 復合物,復合物再向細胞核內轉運,隨后以組織和發育階段特有的方式啟動基因轉錄,從而調節細胞的生命活動[54-55]。該通路涉及到BMP2介導的多種細胞調節活動。Ahsan 等[56]將惡性外周神經鞘瘤(Malignant Peripheral Nerve Sheath Tumors,MPNSTs)細胞用BMP2 I型受體抑制劑LDN-193189 處理,發現Smad1/5/8 的表達下調,并且MPNSTs 細胞的活性和侵襲性顯著降低,抑制了MPNSTs 細胞的惡性擴散。在脊椎動物胚胎心臟前體細胞中BMP2和Wnt信號通路下游因子糖原合成酶激酶3β(Glycogen Synthase Kinase 3 beta,GSK3β)促進Smad1 磷酸化,調節心臟前體細胞的遷移軌跡,促進心臟的形成[54]。在顱骨缺損模型中發現,胞外鈣離子可增強BMP-2對Smad 信號通路的磷酸化,使骨鈣素、成骨特異性轉錄因子(Runt-related Transcription Factor2,Runx2)和成骨相關轉錄因子(Osterix,Osx)的表達上調,促進了體內骨再生[57]。
4.3BMP2與MAPK 信號通路 促分裂素原活化蛋白激酶(Mitogen-Activated Protein Kinases,MAPKs)包括應激活化蛋白激酶(c-Jun N-terminal Kinase,JNK)、細胞外信號調節激酶(Extracellular Regulated Protein Kinases,ERK)和p383 個亞族[58]。該通路同樣參與癌癥的發生,體內外試驗表明抑制BMP2的表達或者抑制p38 的表達會抑制肝癌細胞增殖、遷移、侵襲以及微血管密度和血管生成[59]。同時BMP2通過該通路調節細胞分化,在C2C12 間充質干細胞中發現Runx2上游因子p38被BMP2激活,使Runx2的表達上調,促進間充質前體細胞的分化[60]。將原代小鼠顱源性成骨細胞在BMP2培養基中誘導培養,并抑制p38的表達,堿性磷酸酶活性也會降低;抑制JNK 的表達,骨鈣素的表達明顯下調,從而影響骨分化[61]。此外,Yang 等[62]研究發現,BMP-2磷酸化JNK,可以增強成骨細胞分化。
4.4BMP2與PPARγ信號通路PPARs是具有調節血糖和脂質平衡的一種過氧化物酶體增殖物激活受體,是一種核激素受體和轉錄因子,包括PPAR-α、PPAR-γ和PPAR-δ3種亞型[63]。PPARγ通路在不同的細胞中調節細胞的增殖和分化,在細胞生命活動中起重要的作用。在間充質干細胞中PPARγ的激活能夠增強Runx2啟動子的組蛋白激活標記來影響BMP2的作用,同時PPARγ靶基因啟動子的組蛋白激活標記的表達在一定條件下也可通過BMP2刺激而提高,因此BMP2與PPAR-γ信號通路是相互交叉的,共同調節細胞活動[64]。通過構建慢病毒載體過表達BMP2與前體脂肪細胞中發現,PPARγ的表達顯著上調,促進前體脂肪細胞的分化[6]。此外,在血管平滑肌細胞中PPARγ通過連接上游因子TGF-β1 和BMP2,協同調節細胞增殖和糖代謝[65]。
5 總結及展望
BMP2基因從發現至今取得了較大的研究進展,其在組織中的生物學功能基本被明確。BMP2作為最活躍的骨生長因子,現已被應用于臨床治療。目前的研究發現BMP2在成骨過程中與其他生長因子或成骨基因之間相互作用形成成骨調控網絡,這為臨床骨治療提供新的思路。同時BMP2在癌細胞內廣泛表達,可作為檢測癌癥的標志物,BMP2基因既可以促進癌細胞的增殖與擴散,又能抑制癌細胞的增長。同樣地,在BMP2介導的癌癥發生的多條信號通路中,也存在抑制或促進癌細胞的遷移、擴散和增殖,針對這一矛盾結果,還需要深入研究。本實驗室前期研究表明BMP2可能與綿羊尾部脂肪沉積相關,影響綿羊尾型的發育。進一步的研究表明BMP2基因能夠促進綿羊前體脂肪細胞的分化,但是機制尚不明確。后期研究將集中于BMP2在綿羊尾部脂肪沉積的調控機理,為培育不同尾型的綿羊提供理論基礎。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
