多毛番茄CIPK 基因家族生物信息學分析及低溫下功能研究
2021-03-14 05:54:00狄成乾王傲雪
中國蔬菜 2021年2期
關鍵詞:分析
柴 暢 狄成乾 汪 楊 王傲雪,*
(1 東北農(nóng)業(yè)大學園藝園林學院,黑龍江哈爾濱 150030;2 東北農(nóng)業(yè)大學生命科學學院,黑龍江哈爾濱 150030)
番茄起源于熱帶,是一種對低溫較為敏感的蔬菜,溫度過低會使番茄生長發(fā)育不良,嚴重時甚至死亡。多毛番茄(Solanum habrochaites)是番茄屬的一種野生番茄,具有耐低溫甚至是抗凍的特性(Venema et al.,1999,2005;Foolad &Lin,2000)。研究表明,與多毛番茄遠緣雜交可以提高栽培番茄的抗性和品質(zhì),因此許多育種工作者將多毛番茄的特性用于番茄抗病蟲性改良和創(chuàng)造抗病蟲種質(zhì)資源(Rick,1973)。然而番茄遠緣雜交不親和,剖析多毛番茄基因調(diào)控的信號轉(zhuǎn)導通路,繼而通過基因轉(zhuǎn)移方法改造栽培番茄,能創(chuàng)造耐低溫的栽培番茄,也可以為其他作物遺傳改良提供基因材料。
鈣離子信號是植物中一種普遍的信號轉(zhuǎn)導途徑,鈣離子作為真核生物中通用的離子幾乎涉及植物生長的各個方面并參與調(diào)控過程。鈣離子濃度升高到某些閾值水平能激活鈣離子傳感器蛋白和離子通道,調(diào)控細胞質(zhì)中游離鈣離子濃度變化,產(chǎn)生相應生理反應。這些傳感器主要包括鈣調(diào)素蛋白(CaM)、鈣依賴型蛋白激酶(CDPK)和類鈣調(diào)磷酸酶B 蛋白(CBL)。CBL 作為傳感器需要與相互作用的蛋白激酶(CIPK)特異性結(jié)合形成功能上的復合物傳遞植物信號,調(diào)控離子流入。
CIPK 蛋白包含保守的N 端的激酶結(jié)構域和C端的調(diào)節(jié)結(jié)構域。前者包含絲氨酸/蘇氨酸蛋白激酶催化結(jié)構域,有可以磷酸化的激活環(huán)。后者包含磷酸酶相互作用域PPI 和自調(diào)節(jié)結(jié)構域NAF(又稱FISL 基序)。特征氨基酸Asn(N)、Ala(A)、Phe(F)、Ile(I)、Ser(S)和Leu(L)組成的NAF基序是結(jié)合CBL 蛋白所必需的(Albrecht et al.,2001)。當CBLs 與CIPKs 相互作用時,自抑制的NAF 結(jié)構域從蛋白質(zhì)結(jié)構域釋放,產(chǎn)生活性激酶構象(Weinl &Kudla,2009)。
CBL-CIPK 復合物在非生物脅迫、生物脅迫以及營養(yǎng)信號級聯(lián)中有重要的作用。許多CBLCIPK 網(wǎng)絡已被證明參與了植物在非生物脅迫條件下的信號傳遞,特別是在鹽脅迫和滲透脅迫下。蛋白激酶CIPK24/SOS2 在鹽脅迫條件下結(jié)合蛋白CBL4/SOS3,進而激活下游成分SOS1/(Na+/H+antipoter),從而改善鹽脫毒過程(Qiu et al.,2002)。過表達SICIPK24/SISOS2增加了番茄的耐鹽性(Huertas et al.,2012)。AtCBL1激活AtCIPK23來調(diào)節(jié)接收器NPF6;3,同時激活K+通道AKT1 抑制銨離子的轉(zhuǎn)運并調(diào)節(jié)植物對銨離子的敏感性,AtCIPK23和AtAKT1的突變體植株葉片氣孔關閉,可以減少植物失水,從而間接響應干旱脅迫(Nieves-Cordones et al.,2012;Straub et al.,2017)。CBL-CIPK 網(wǎng)絡在低溫下也有一些研究。在擬南芥中,AtCBL9-AtCIPK3 和AtCBL1-AtCIPK7 復合物在冷脅迫下有重要的作用(Huang et al.,2011)。AtCBL1、AtCBL9和AtCIPK26互作激活NADPH 氧化酶RBOHF(respiratory burst oxidase homolog F),從而作用于細胞中的H2O2(Drerup et al.,2013)。MdCIPK22-MdSUT2.2 促進了糖積累從而增加了耐旱性(Ma et al.,2019)。
CIPKs 作為鈣離子信號轉(zhuǎn)導途徑中的一個重要蛋白家族,在多毛番茄中還未見報道。為確定多毛番茄CIPK 基因家族成員,本試驗利用生物信息學方法對多毛番茄CIPK 基因家族成員的序列信息進行了鑒定和分析,采用實時熒光定量PCR 的方法研究了冷脅迫下各成員的表達變化,以期為進一步探索冷脅迫下多毛番茄鈣信號轉(zhuǎn)導途徑奠定基礎。
1 材料與方法
1.1 材料
供試多毛番茄(Solanum habrochaites)為東北農(nóng)業(yè)大學園藝園林學院保存的LA1777。將消毒后的種子放在濕濾紙上,發(fā)芽后將其播種到裝有蛭石、土壤混合物(1V∶1V)的塑料盆中,置于20~25 ℃的人工氣候室中,光周期16 h/8 h(晝/夜)。同時對同一批次的多毛番茄幼苗進行低溫脅迫(4 ℃)處理,取樣時間為處理后0、3、9、12、24、36 h。采集植株形態(tài)學上端第2 片完全展開的功能葉,各個處理的樣品均準確稱取0.08 g,3 次生物學重復,將樣品快速轉(zhuǎn)移至液氮中,用于后續(xù)RNA 的提取。
1.2 方法
1.2.1 多毛番茄CIPK 基因家族成員鑒定及基本信息分析 從Pfam 蛋白家族數(shù)據(jù)庫(https://pfam.xfam.org/)下載蛋白酶(PF00069)和NAF 結(jié)構域(PF03822)對應的隱馬爾可夫模型(HMM)文件,再通過hmmsearch 從多毛番茄全基因組數(shù)據(jù)(https://tomato.biocloud.net)中搜索CIPK基因(E-value <1×10-7,相似度>50%)(Finn et al.,2007)。用bio-linux 軟件預測多毛番茄CIPK 蛋白的長度、分子質(zhì)量、等電點。利用MapChart 軟件繪制染色體定位圖。利用SOSUI(http://harrier.nagahama-i-bio.Ac.jp/sosui/)在線軟件預測蛋白跨膜結(jié)構域。利用CELLO(http://cello.life.nctu.edu.tw)預測亞細胞定位。
1.2.2 多毛番茄CIPK 基因家族系統(tǒng)進化分析 為探究CIPK 基因家族內(nèi)的進化關系,使用MEGA軟件的鄰接法(Neighbor-Joining)對多毛番茄、栽培番茄、擬南芥、楊樹的CIPK 基因家族進行系統(tǒng)進化分析,校驗參數(shù)500 次重復。
1.2.3 多毛番茄CIPK 基因家族基因結(jié)構分析 提取多毛番茄CIPK基因CDS 序列及對應的基因組序列信息,運用GSDS 工具(http://gsds.cbi.pku.edu.cn/)進行基因結(jié)構分析并繪制外顯子-內(nèi)含子結(jié)構圖。
1.2.4 多毛番茄CIPK 基因家族保守基序分析 為研究多毛番茄CIPK基因的進化關系,用MEME(http://meme.nbcr.net/meme/)在線分析18 個多毛番茄CIPK基因的保守基序。
1.2.5 多毛番茄CIPK 基因家族基因復制 使用MCSCanX 的默認參數(shù)對多毛番茄和擬南芥、多毛番茄和栽培番茄進行共線性關系分析。利用ClustalW 對全長氨基酸序列進行比對,使用KaKs_Calculator 2.0 計算Ks 和Ka 程序,用同義突變率估計基因?qū)Φ碾x散時間,每年每個同義位點的替換量為T=Ks/2X(X=6.56×10-9)(Yuan et al.,2015)。
1.2.6 實時熒光定量PCR 分析 使用美國OMEGA公司生產(chǎn)的RNA-Solv Reagent 提取RNA,具體步驟參照說明書,RNA 提取過程所需其他試劑均為國產(chǎn)分析純。RNA 逆轉(zhuǎn)錄試劑盒使用日本東洋紡公司ReverTra Ace qPCR RT Master Mix with gDNA Remover。利用NCBI(https://www.ncbi.nlm.nih.gov/)數(shù)據(jù)庫中Primer Blast 功能設計熒光定量引物(Max值設為200),引物序列見表1,利用qRT-PCR 方法檢測多毛番茄CIPK 基因家族各成員在4 ℃處理0、3、9、12、24、36 h 的表達量。實時熒光定量PCR 分析所用熒光染料試劑盒為北京全式金生物技術有限公司TransStart Top Green qPCR SuperMix。使用儀器為QuantStudio1。反應程序為94 ℃ 30 s;94 ℃ 5 s,50~60 ℃ 15 s,72 ℃ 10 s;40 個循環(huán)。以穩(wěn)定的Actin基因作為內(nèi)參,與0 h 未處理葉片(CK)相比并計算,采用2-ΔΔCT方法進行分析。

表1 熒光定量PCR 引物序列
2 結(jié)果與分析
2.1 多毛番茄CIPK 家族成員鑒定
利用hummer 模型分析,鑒定出多毛番茄中24 個CIPK 基因家族成員,將其命名為ShCIPK1~ShCIPK24,在12 條染色體上均有分布,其中6、9、12 號染色體上分布最多,均為4 個,5號染色體有3 個,2 號染色體有2 個,其余染色體上均為1 個(圖1)。從表2 可以看出,多毛番茄CIPK 家族基因長度差距不大,范圍在1 260~1 464 bp,編碼氨基酸的范圍為419~487 aa。等電點最小(ShCIPK21)為 6.37,最大(ShCIPK19)為9.90,除ShCIPK5、ShCIPK6、ShCIPK21外,等電點均大于7,共21 個堿性蛋白。分子量變化范圍為48 497.8~108 454.7 Da,變化較大。亞細胞定位分析表明,大多數(shù)基因定位于細胞質(zhì),少數(shù)基因分布于質(zhì)膜、細胞核、線粒體。大多數(shù)家族成員存在1個跨膜結(jié)構域。多毛番茄CIPK 基因家族的NAF結(jié)構域中殘基A、F、I 和L 是絕對保守的,表明這些殘基可能在與CBLs 的相互作用中起重要作用(圖2),起始位置在300~350 bp 之間,長度為20個氨基酸左右。
2.2 多毛番茄CIPK 基因家族系統(tǒng)進化分析
為了研究多毛番茄與其他物種的進化關系,對多毛番茄、栽培番茄、擬南芥和楊樹的CIPK 基因家族構建進化樹。由圖3 可知,98 個CIPK基因可分為5 個亞族,分別命名為group 1~group 5。其中group 1 包括ShCIPK7、ShCIPK12、ShCIPK13、ShCIPK17、ShCIPK235 個多毛番茄基因家族成員;group 2 包 括ShCIPK2、ShCIPK9、ShCIPK11、ShCIPK18、ShCIPK195 個家族成員;group 3 包括ShCIPK4、ShCIPK10、ShCIPK14、ShCIPK15、ShCIPK16和ShCIPK226 個基因家族成員;group 4 包括ShCIPK5、ShCIPK6、ShCIPK21、ShCIPK244 個家族成員;group 5 包括ShCIPK1、ShCIPK3、ShCIPK8、ShCIPK204 個家族成員。5 個group 中group 1 為3 個垂直同源對,其余4 個group 均有4個垂直同源對,除ShCIPK17和PtCIPK22這對垂直同源對外,其他均為多毛番茄和栽培番茄垂直同源對。結(jié)果表明,在多毛番茄與栽培番茄基因組之間,CIPK 基因家族具有相似的基因。
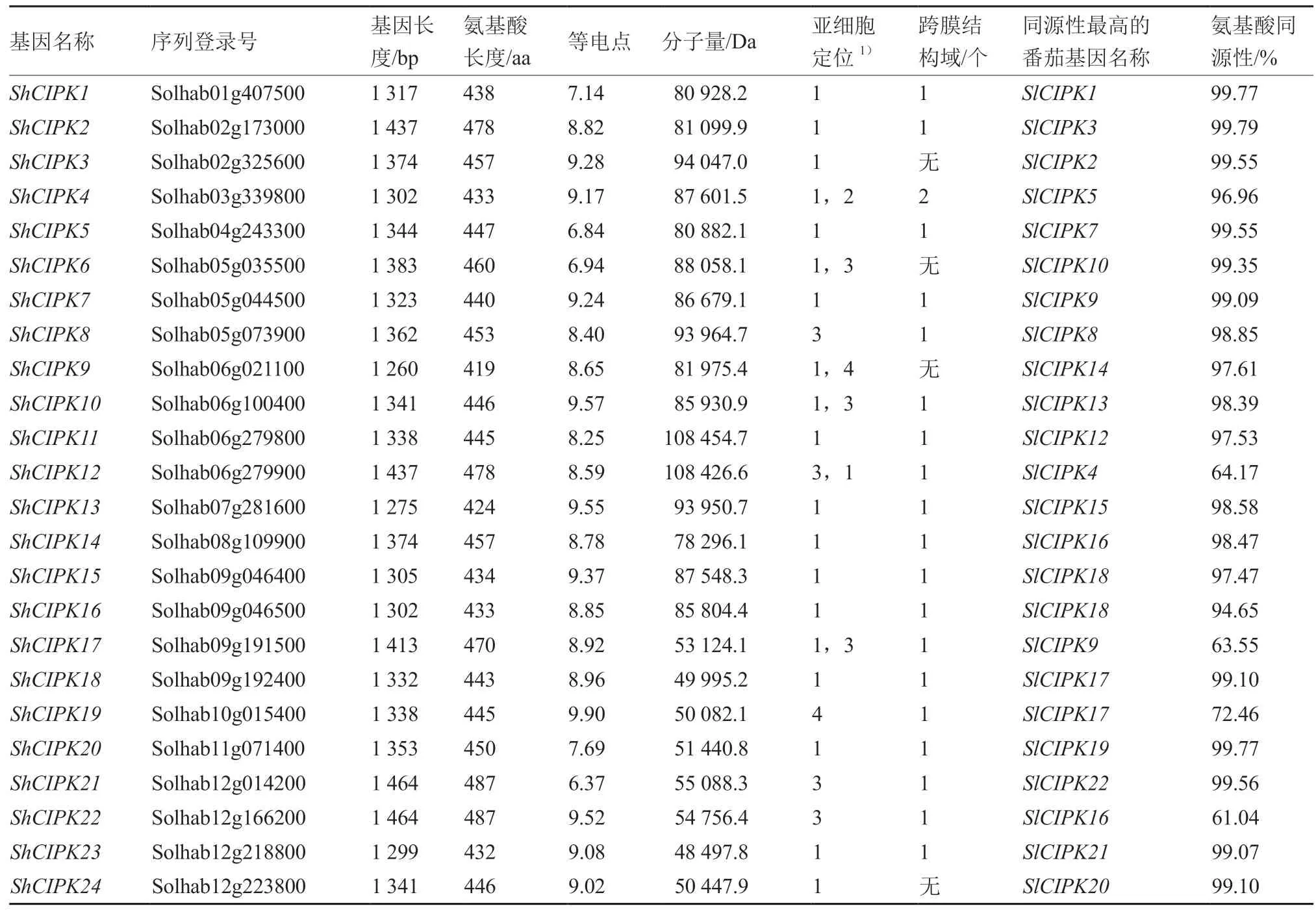
表2 多毛番茄CIPK 基因家族特征分析
2.3 多毛番茄CIPK 基因家族結(jié)構分析
如圖4 所示,多毛番茄CIPK 基因家族分為內(nèi)含子富集和內(nèi)含子缺失兩類,其中ShCIPK4、ShCIPK9、ShCIPK10、ShCIPK13、ShCIPK14、ShCIPK15、ShCIPK16、ShCIPK17、ShCIPK18、ShCIPK22、ShCIPK23無內(nèi)含子,ShCIPK7、ShCIPK11、ShCIPK12、ShCIPK19只有1 個內(nèi)含子,ShCIPK2有3 個內(nèi)含子,屬于內(nèi)含子缺失;ShCIPK6、ShCIPK21有11 個內(nèi)含子,ShCIPK5有13 個內(nèi) 含 子,ShCIPK1、ShCIPK3、ShCIPK8、ShCIPK20、ShCIPK24有14 個內(nèi)含子,以上成員均屬于內(nèi)含子富集組。
2.4 多毛番茄CIPK 基因家族基序分析
運用MEME 軟件在番茄CIPK基因家族中預測到18 個基序,基序類型及排列順序基本一致,同源性越高的基因基序排列相似性越高(圖5)。24 個ShCIPK基因均有Motif1~Motif8 和Motif11。除ShCIPK4、ShCIPK15、ShCIPK9外均有Motif10。ShCIPK4、ShCIPK15和ShCIPK16具有相似基序且排列順序相同,相對于家族中其他成員額外含有Motif14、Motif15 和Motif17。ShCIPK21、ShCIPK6還含有Motif18。
2.5 多毛番茄CIPK 基因家族基因復制與共線性分析
從表3 可以看出,多毛番茄CIPK 基因家族中包括ShCIPK15:ShCIPK16,ShCIPK1:ShCIPK11,ShCIPK23:ShCIPK13,ShCIPK4:ShCIPK15,ShCIPK9:ShCIPK11共5 個基因復制對。4 段重復的Ka/Ks 范圍為0.031 85~0.270 09,可推測出分離時間為8.51~276 Mya(Million years ago,百萬年以前)。這5 個復制基因?qū)Φ钠沃貜偷腒a/Ks 值均小于1,說明它們都經(jīng)過了純化選擇。
為了進一步推斷多毛番茄CIPK基因家族的系統(tǒng)發(fā)育關系,分別構建了模式植物擬南芥和多毛番茄(圖6)、栽培番茄和多毛番茄(圖7)種間共線性分析圖。分析結(jié)果表明,在擬南芥中有14 個基因?qū)εc多毛番茄存在共線性關系。存在1 個基因與2 個基因共線性關系的現(xiàn)象,如ShCIPK2與AtCIPK12和AtCIPK13,ShCIPK15與AtCIPK4和AtCIPK7,ShCIPK19與AtCIPK14和AtCIPK22。擬南芥與多毛番茄中的共線性關系對還有ShCIPK1與AtCIPK3,ShCIPK3與AtCIPK23,ShCIPK4與AtCIPK4,ShCIPK5與AtCIPK8,ShCIPK9與AtCIPK11,ShCIPK10與AtCIPK25,ShCIPK12與AtCIPK10,ShCIPK21與AtCIPK1。

表3 多毛番茄CIPK 基因家族中的基因復制
栽培番茄和多毛番茄中存在的共線性基因?qū)τ?2 個,包括ShCIPK7與SlCIPK9,ShCIPK8與SlCIPK8,ShCIPK9與SlCIPK14,ShCIPK10與SlCIPK13,ShCIPK11與SlCIPK12,ShCIPK13與SlCIPK15,ShCIPK14與SlCIPK16,ShCIPK18與SlCIPK17,ShCIPK20與SlCIPK19,ShCIPK21與SlCIPK22,ShCIPK23與SlCIPK21,ShCIPK24與SlCIPK20。栽培番茄與多毛番茄基因之間的共線性關系存在于同條染色體上。ShCIPK10和ShCIPK21這2 個基因的共線性關系存在于擬南芥和栽培番茄中,ShCIPK10、ShCIPK21、AtCIPK25、AtCIPK1、SlCIPK13、SlCIPK22等可能在祖先分離之前就已經(jīng)存在。
2.6 多毛番茄CIPK 基因家族順式作用原件分析
啟動子的結(jié)構影響啟動子和RNA 聚合酶的親和力,從而影響基因表達水平。對多毛番茄CIPK基因家族成員上游1 500 bp 進行分析,存在多個與植物逆境應答相關的順式作用元件(表4)。在順式作用元件分析當中,除常見的 CAAT-box 和TATAbox 原件之外,可以將其分為3 類。第一類是與激素相關的原件,如ABRE、TCA。第二類是與調(diào)控相關的原件,如光(Light)調(diào)控原件。第三類是與生物脅迫相關的原件,如MBS、LTR。所有成員均有光調(diào)控順式作用元件,超過一半的成員具有脫落酸(ABRE)、水楊酸(TCA)、厭氧誘導(ARE)、茉莉酸甲酯(CGTCA-motif)順式作用元件。9 個家族成員有防衛(wèi)和逆境應答(TC-rich repeats)順式作用元件,8 個家族成員有冷脅迫(LTR)順式作用原件,7 個家族成員有干旱脅迫(MBS)順式作用元件,這些家族成員可能與脅迫應答相關。有防衛(wèi)應答(W-BOX)、赤霉素(P-BOX)順式作用元件的家族成員分別為6 個和5 個。

表4 多毛番茄CIPK 基因家族的順式作用元件分析
2.7 多毛番茄CIPK 基因家族在低溫脅迫下的表達分析
從圖8 可以看出,多毛番茄CIPK 基因家族中各成員在低溫脅迫下的表達量均有一定程度的變化,其中ShCIPK1、ShCIPK2、ShCIPK4、ShCIPK5、ShCIPK7、ShCIPK8、ShCIPK9、ShCIPK13、ShCIPK14、ShCIPK15、ShCIPK16、ShCIPK17、ShCIPK18、ShCIPK19、ShCIPK20、ShCIPK22、ShCIPK24共17 個基因家族成員低溫誘導后上調(diào)表達明顯,ShCIPK19、ShCIPK17表達量上調(diào)尤為顯著。ShCIPK19在低溫誘導下的表達量峰值最大,4 ℃誘導12 h 時的表達量為0 h 未處理葉片(對照)的89.27 倍,36 h 的表達量為對照的23.20 倍;ShCIPK17在低溫誘導12 h 的表達量為對照的70.34 倍,36 h 的表達量為對照的36.61倍。在低溫處理中大多數(shù)家族成員表現(xiàn)出表達量波動的趨勢,只有ShCIPK2表達量持續(xù)上升,在36 h 達到峰值,為對照的54.13 倍。
3 討論與結(jié)論
凍害阻礙了大多數(shù)植物的生長,對番茄的大面積栽培造成越來越大的威脅。多毛番茄具有抗凍特性,可以作為優(yōu)秀的種質(zhì)材料。在以往的研究中,Ca2+在冷脅迫中起著重要的作用,CIPK-CBL 配合物的功能與Ca2+密切相關。空泡Ca2+/H+逆向轉(zhuǎn)運蛋白增強了CBF(CRT/DRE binding factor)基因的表達,使其具有更強的耐低溫能力。ShCBF1在轉(zhuǎn)基因煙草中過量表達,正調(diào)控其下游抗冷相關基因表達和相關蛋白含量改變,表現(xiàn)出更強的耐低溫能力(葛君 等,2017)。AtCIPK3的表達對寒冷、干旱、ABA、高鹽和創(chuàng)傷均有反應,AtCIPK3可作為冷反應信號通路中ABA 依賴、ABA 獨立通路之間的“節(jié)點”(Kim et al.,2003)。AtCIPK7通過與AtCBL1相互作用在冷信號轉(zhuǎn)導中發(fā)揮作用,AtCIPK7在被AtCBL1激活后,可能調(diào)節(jié)參與冷反應的其他蛋白質(zhì)組分的活性(Huang et al.,2011)。BdCIPK31的異位表達增強了低溫下ROS 解毒體系和滲透保護劑生物合成,參與低溫脅迫應答的轉(zhuǎn)錄調(diào)控,是植物低溫適應的一個正調(diào)控因子(Luo et al.,2018)。過表達OsCIPK3提高了轉(zhuǎn)基因植株的抗寒性(Xiang et al.,2007)。小麥TaCIPK7、TaCIPK15、TaCIPK24、TaCIPK32分別誘導對低溫脅迫的應答(Sun et al.,2015)。番茄SlCIPK1在低溫下的表達量顯著高于其他成員(王傲雪和劉思源,2018)。BnaCIPK基因在冷脅迫下轉(zhuǎn)錄水平會顯著提高(Zhang et al.,2014)。
本試驗通過生物信息學分析篩選出多毛番茄中的24 個CIPK 基因家族成員。植物CBL和CIPK基因最早在模式植物擬南芥中被發(fā)現(xiàn),在擬南芥中有26 個CIPKs(Kolukisaoglu et al.,2004)。目前在多種植物中也有了CIPK 基因家族的報道。茄科植物中,栽培番茄有22 個CIPK 基因家族成員(王傲雪和劉思源,2018),茄子中有15 個(Li et al.,2016),辣椒中有21 個(劉思源 等,2017)。禾本科植物中,水稻有31 個CIPK 基因家族成員(Xiang et al.,2007),高粱有32 個(Weinl &Kudla,2009)、玉米有43 個(Chen et al.,2011)、小麥有32 個(Sun,2015)。木本植物中,楊樹有27 個CIPK 基因家族成員(Zhang et al.,2008)。藤本植物葡萄有20 個(Xi et al.,2017)。除此之外,還在藻類、苔蘚、蕨類、葫蘆科等植物中發(fā)現(xiàn)了CIPK基因家族。多毛番茄CIPK 基因家族在6、9、12號染色體上分布較多,在其他染色體上分布較少這種分布不均勻的現(xiàn)象,與禾本科植物谷子、玉米的發(fā)現(xiàn)一致,基因的不平衡分布可能與物種進化和遺傳變異有關。
由多毛番茄、栽培番茄、擬南芥和楊樹構建的系統(tǒng)進化樹分為5 個亞族,每個亞族均有ShCIPK分布。group 1 包括3 對多毛番茄與栽培番茄垂直同源對。group 2、group 3、group 4、group 5 均包括4 對多毛番茄與栽培番茄的垂直同源對。這些結(jié)果表明多毛番茄和栽培番茄相關的同系物基因有著保守的結(jié)構功能。對比而言,擬南芥與多毛番茄沒有垂直同源對,說明多毛番茄的CIPK 基因家族與擬南芥的功能進化上出現(xiàn)了偏離。多毛番茄ShCIPKs分為內(nèi)含子富集組和內(nèi)含子缺失組,內(nèi)含子外顯子的組成都夠反映基因家族的進化關系,內(nèi)含子富集 的ShCIPK1、ShCIPK3、ShCIPK5、ShCIPK6、ShCIPK8、ShCIPK20、ShCIPK21、ShCIPK24、ShCIPK6在系統(tǒng)進化樹中位于group 4 和group 5;內(nèi)含子缺失的位于系統(tǒng)進化樹的group1、group 2、group 3。擬南芥CIPK 基因家族也分為2 個分支,第1 種不含內(nèi)含子或者只含有1 個內(nèi)含子,第2 種含有9 個或者以上的內(nèi)含子。內(nèi)含子獲得的速率相比內(nèi)含子缺失的速率更慢,內(nèi)含子缺失的CIPK基因的系統(tǒng)發(fā)生可能是在內(nèi)含子富集的基因之后,推測是內(nèi)含子富集的基因mRNA 反轉(zhuǎn)錄過程中,重新插入基因組而出現(xiàn)的分支。
在ShCIPKs中存在2 對串聯(lián)重復基因和3 對片段重復基因。研究表明,片段重復源于脅迫。這5 對基因在保守motif 上表現(xiàn)出高度一致,表明串聯(lián)復制和片段復制在ShCIPKs的擴增上有重要的作用。串聯(lián)重復基因?qū)Φ腒s 在0.111 70~0.748 34,分離時間在8.51~57 Mya。片段重復基因?qū)Φ腒s 在1.918 40~3.623 04,分離時間在146~276 Mya。單雙子葉分化的時期在200~205 Mya,推測ShCIPKs的片段復制發(fā)生在單雙子葉分化時期(Wolfe et al.,1989)。
ShCIPKs在冷脅迫下的波動表達與小麥和玉米一致,證明ShCIPKs可能在冷脅迫應答中有重要的作用。SlCIPK1在栽培番茄CIPK 基因家族中受冷誘導表達量上升最大,表達量的峰值出現(xiàn)在3 h,為對照未處理葉片的50.91 倍(劉思源,2018)。本試驗中,ShCIPK19、ShCIPK17上調(diào)表達顯著,ShCIPK19在低溫誘導下的表達量峰值最大,冷誘導12 h 時的表達量為對照的89.27 倍;ShCIPK17的表達量峰值在低溫誘導下12 h,表達量為對照的70.34 倍;只有ShCIPK2的表達量持續(xù)上升,推測其可能對低溫這種非生物脅迫持續(xù)作出相應反應,在36 h 達到峰值,為對照的54.13 倍。ShCIPK2、ShCIPK17和ShCIPK19這3 個基因比SlCIPK1的表達量峰值更高,具有研究價值。ShCIPK19與AtCIPK3的相似度為47.41%,推測ShCIPK19可能具有與AtCIPK3相似的功能。ShCIPK2、ShCIPK17和ShCIPK19含有LTR 冷脅迫應答原件,這與冷脅迫下的實時熒光定量試驗結(jié)果相吻合。
本試驗對多毛番茄CIPK 基因家族進行了全基因組鑒定、生物信息學分析和表達分析,推測ShCIPK2、ShCIPK17和ShCIPK19可能參與低溫脅迫下多毛番茄鈣信號調(diào)控,然而目前對多毛番茄抗冷機制認識尚未明確,接下來的試驗應進一步研究ShCIPKs對冷脅迫的分子機制,打破地理分布對作物種植和園藝植物生產(chǎn)的影響。
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
當代經(jīng)濟研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
