我國農作物灰霉病殺菌劑的應用現狀
2021-03-14 05:53:58張正煒石小媛武向文戴富明
中國蔬菜 2021年2期
張正煒 陳 秀,2 石小媛 成 瑋* 武向文 戴富明
(1 上海市農業技術推廣服務中心,上海 201103;2 上海市農藥檢定所,上海 201103;3 上海源怡種苗股份有限公司,上海 201314;4 上海市農業科學院,上海 201403)
灰霉病(gray mold)因其致病菌會在病部植物組織表面生長產生大量灰色霉層而得名,是最早被發現和描述的植物真菌性病害之一。早在1729 年雙名制引入以前,意大利植物學家米切利即根據其觀察到的致病菌分生孢子梗的特點用希臘詞“束狀的葡萄漿果”來命名灰霉病菌。1801 年法國植物學家佩爾松將這一致病菌納入雙名法的系統之內,并定名為Botrytis cinerea,即灰葡萄孢菌。這個由葡萄狀分生孢子梗真菌引起的病害,又隨著世界葡萄種植面積的擴張而在全球流行起來。
灰霉病菌源復雜。目前發現的葡萄孢屬真菌有20 多個種,其中多數能夠引起植物灰霉病。如現已明確能夠引起蔥屬植物灰霉病的病原菌就有蔥鱗葡萄孢菌(B.squamosa)、蔥腐葡萄孢菌(B.aclada或B.allii)、大蒜盲種葡萄孢菌(B.porri)、蔥細絲葡萄孢菌(B.byssoidea)、中國蔥葡萄孢菌(B.sinoallii)、大蔥球形葡萄孢菌(B.globosa)、球精葡萄孢菌(B.sphaerosperma)以及灰葡萄孢菌(B.cinerea)等8 個種(崔蘊剛 等,2020)。同時,葡萄孢菌中存在的異核現象和非整倍體分裂現象能引起后代豐富的遺傳及形態表型變異。如不同寄主植物間的灰葡萄孢菌可互相引發致病,不同菌株雖不具有明顯的寄主專化性,但卻具有明顯的致病力分化(張艷杰 等,2017)。后來陸續有研究證實B.cinerea是一種復合體(Giraud et al.,1997;Fournier et al.,2003)。在分子生物學方法引入之后,假灰葡萄孢菌(B.pseudocinerea)得以分離(Walker et al.,2011),此后諸如B.sinoviticola(Zhou et al.,2013)、B.deweyae(Grant-Downton et al.,2014)、B.californica(Saito et al.,2016)和B.euroamericana(Garfinkel et al.,2017)等眾多Botrytissp.的新種又陸續被分離和報道。鑒于葡萄孢屬真菌的這些特點,國際殺菌劑抗性行動委員會(Fungicide Resistance Action Committee,FRAC)將該菌歸為高抗藥性風險的病原菌。
1 我國防治灰霉病殺菌劑的登記現狀
灰霉菌的寄主廣泛。從以獲取糧食、油料和纖維為主的大田作物到生產蔬菜、水果和花卉的保護地經濟作物,灰霉菌在全球范圍內寄生為害200 多種農作物。其中以溫帶和亞熱帶地區的雙子葉植物為主。實際上自然界中灰霉菌的寄主范圍還要更廣泛(Elad et al.,2016),不斷有新的作物寄主被報道。灰霉菌是一種死體營養型的寄生性真菌,即病菌侵染后先殺死寄主的細胞和組織,然后從死亡的細胞中吸取養分。所以灰霉菌對農業的為害從作物田間生長階段一直延續到收獲后的貯藏過程。隨著作物的集約化種植,設施園藝的發展及草莓、藍莓等漿果類水果的普及,灰霉病在我國的發生與為害日益嚴重。化學防治已成為降低灰霉病為害不可或缺的防治措施,且依賴程度與日俱增(胡偉群和陳杰,2002;陳治芳 等,2010)。
截至2020 年4 月,我國登記防治灰霉病的殺菌劑商品共有471 個,其中單劑317 個,混劑154個。登記作物15 種,以防治茄科、葫蘆科、葡萄科以及薔薇科等經濟作物灰霉病為主,其中防治番茄灰霉病的商品最多(242 個),其次為黃瓜(148個)、葡萄(83 個)和草莓(27 個)。除了蔬菜和水果以外,菊花、百合、牡丹等觀賞花卉以及人參等經濟價值較高的保護地作物上也有少量商品登記。
單劑登記有效成分35 種,嘧霉胺登記產品數量最多,有82 個,其次為腐霉利(54 個)、異菌脲(45個)和啶酰菌胺(33 個)(圖1)。混劑復配46 個,實際有效成分32 種。登記最多的混劑為唑醚·啶酰胺(21 個),其次為腐霉·福美雙(20 個)和腐霉·百菌清(12 個)。所有藥劑有效成分合計51 種,其中16 種僅以混劑形式登記(表1)。
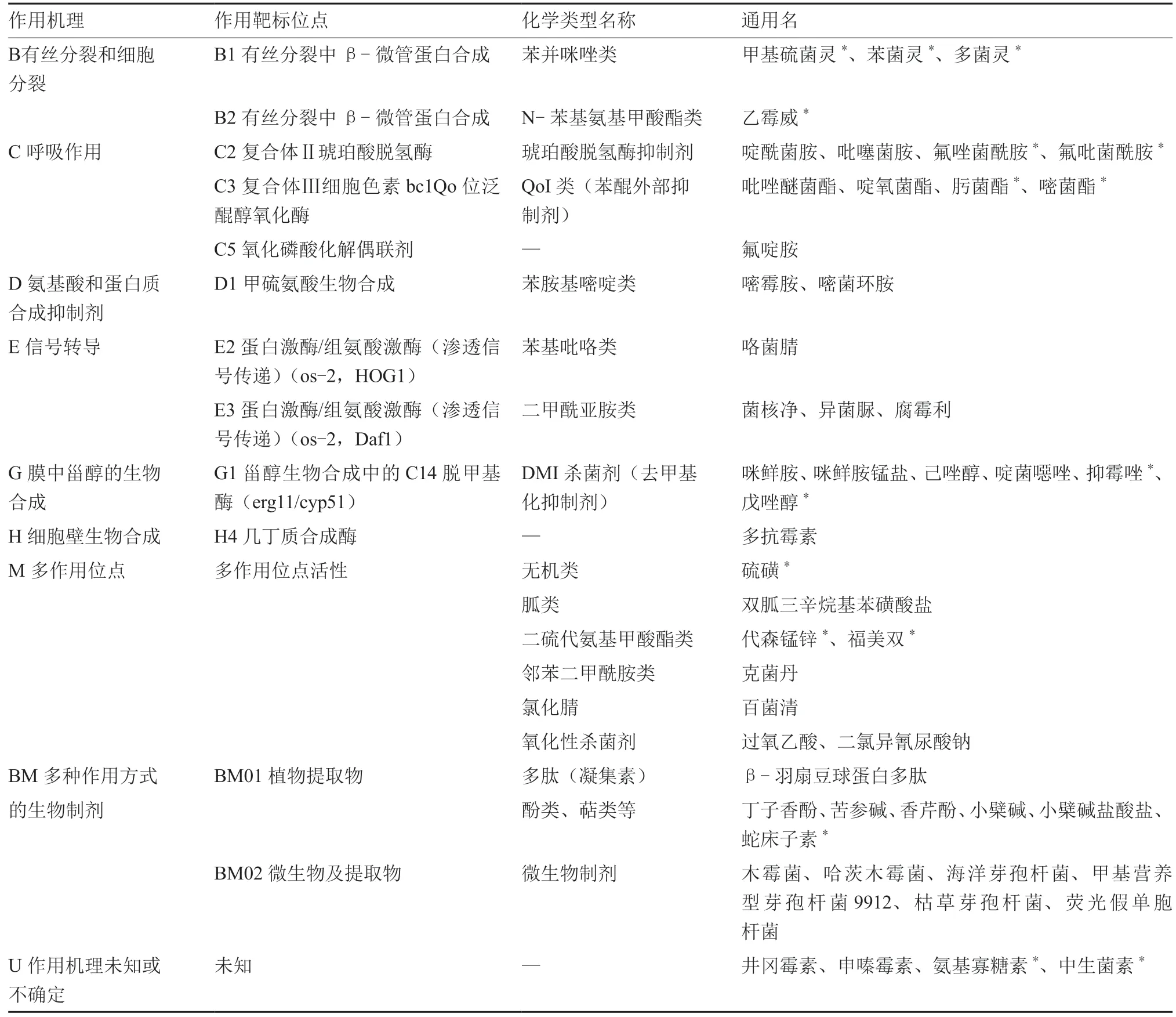
表1 登記殺菌劑的有效成分作用機理分類
由圖2 可知,用于防治番茄灰霉病的殺菌劑主要品種為異菌脲、腐霉利和嘧霉胺,3 個品種登記單劑占比在20%~25%之間;防治黃瓜灰霉病的殺菌劑主要品種為嘧霉胺,目前嘧霉胺登記單劑占比高達50%,腐霉利、啶酰菌胺和過氧乙酸的單劑登記占比均在10%左右;防治葡萄灰霉病登記單劑最多的品種為啶酰菌胺,其次為腐霉利、嘧菌環胺和異菌脲;防治草莓灰霉病的殺菌劑整體登記品種和數量有限,主要有克菌丹、枯草芽孢桿菌、啶酰菌胺和嘧霉胺。
登記最多的植物源殺菌劑為丁子香酚,其次為香芹酚,主要在番茄上登記。微生物防治是防治灰霉病的一種重要手段(肖景惠 等,2019)。目前,用于防治灰霉病的微生物制劑登記數量較多的為木霉菌和枯草芽孢桿菌。木霉菌為拮抗真菌,其制劑在灰霉病發生的多種作物上都有登記。科學合理使用木霉菌防治灰霉病的田間綜合防效甚至優于一般化學藥劑(李萌茵 等,2019)。生防細菌的種類較多,商品化的制劑品種也較真菌豐富,除枯草芽孢桿菌外還有海洋芽孢桿菌、熒光假單胞桿菌等。但除枯草芽孢桿菌以外的細菌制劑品種目前只在黃瓜上登記應用。細菌制劑的田間抑菌效果亦能達到與化學藥劑相當的水平(王紅麗 等,2020)。
2 我國應用化學殺菌劑的主要種類和作用靶標
參照FRAC 對殺菌劑的作用機理分類標準(FRAC Code List 2020),我國目前登記應用的殺菌劑靶標大致可歸為9 類(表1),主要集中在呼吸作用(C)、氨基酸和蛋白質合成抑制劑(D)和信號轉導(E)三大類型。作用于這三類靶標的殺菌劑單劑占登記單劑總數的70%以上,而二甲酰亞胺類和苯胺基嘧啶類殺菌劑是主要的應用品種,二者分別占登記單劑總數的31.5%和28.7%。另外,以啶酰菌胺為代表的琥珀酸脫氫酶抑制劑也是一類比較重要的殺菌劑品種,占登記單劑總數的10.7%。
以有絲分裂和細胞分裂為靶標的苯并咪唑類和N-苯基氨基甲酸酯類殺菌劑,以及二硫代氨基甲酸酯類殺菌劑僅以復配形式登記防治灰霉病。作用于甾醇生物合成中的C14 脫甲基酶(erg11/cyp51)的去甲基化抑制劑(DMI 殺菌劑)也大多以復配形式登記應用,僅有少量產品為單劑。
由于灰霉病菌極易產生抗藥性,登記用于防治灰霉病的殺菌劑更替明顯。以多菌靈為代表的苯并咪唑類殺菌劑多上市于20 世紀60 年代末70 年代初,此類殺菌劑在我國應用時間較早,20 世紀80年代已實現國產化并廣泛應用于病害防治。由于抗藥性的發展,此類殺菌劑早已退出灰霉病防治的主力位置。包括另一類以有絲分裂為靶標的N-苯基氨基甲酸酯類殺菌劑乙霉威在內,目前都僅以混劑形式登記應用。20 世紀70 年代后二甲酰亞胺類殺菌劑開始用于灰霉病的防治,我國于20 世紀80 年代中期開始引進應用。目前從登記商品數量來看,此類殺菌劑仍是防治灰霉病的主力藥劑之一。隨著應用時間的增加,我國灰葡萄孢菌對二甲酰亞胺類殺菌劑的抗性頻率也在不斷上升,但多以低抗菌株為主,并且高抗菌株不具遺傳穩定性(陳夕軍 等,2009),這也是腐霉利、異菌脲等殺菌劑品種得以繼續應用于灰霉病防治的原因之一。苯胺基嘧啶類殺菌劑是另一類用于灰霉病防治的主力藥劑,以20 世紀90 年代開發上市的嘧霉胺和嘧菌環胺為代表。苯胺基嘧啶類殺菌劑是由于灰霉病菌對應用較早的苯并咪唑類和N-苯基氨基甲酸酯類殺菌劑產生抗性后研制而成的(韓之琦 等,2014)。但隨著嘧霉胺等藥劑的大范圍推廣應用,我國蔬菜及葡萄等作物的灰霉病菌對這類藥劑也很快產生了抗藥性(紀明山 等,2003;張瑋 等,2013)。
琥珀酸脫氫酶抑制劑是FRAC 在2009 年劃分出來的一類作用機制相似的化合物。雖然此類化合物早在20 世紀60 年代即有產品面世,但代表性品種多為20 世紀90 年代以后開發上市,并且近年來不斷有新的化合物進行登記。目前應用較多的啶酰菌胺和吡噻菌胺已是相對較早上市的品種。氟唑菌酰胺和氟吡菌酰胺則屬于尚在專利保護期內的新型產品,分別由德國巴斯夫和拜耳公司研發推出,兩者目前僅以混劑形式在我國登記用于防治灰霉病。盡量延緩抗藥性的發展,延長新藥劑的使用壽命,將新化合物的藥效和經濟利益最大化是企業推廣新產品的終極策略。隨著新化合物創制研發成本的提高,新藥劑在抗藥性發展較快的灰霉病防治中的推廣應用要相對更加謹慎。
具有多作用位點活性的殺菌劑以及具有多種作用方式的生物制劑的研發是解決灰霉病菌抗藥性問題的關鍵。雖然登記產品數量有限,但其作為灰霉病化學防治的備選方案在高品質農產品生產中發揮重要作用。我國《綠色食品 農藥使用準則》(NY/T 393—2013)中適用于AA 級綠色食品生產的農藥僅限于植物和動物來源、微生物來源和生物化學產物三大類。其中登記防治灰霉病的單劑品種主要有苦參堿、小檗堿、多抗霉素、井岡霉素、申嗪霉素以及木霉菌、枯草芽孢桿菌、熒光假單胞桿菌等微生物制劑;另有苦參·蛇床素混劑登記用于防治番茄灰霉病。
研究發現對灰霉病菌有抑制作用的植物源提取物不在少數(劉芳 等,2014;葉艷英 等,2016),但目前市場上登記應用的植物源殺菌劑產品仍十分有限。應用較廣泛的有效成分主要是苦參堿和丁子香酚,兩者無論單用還是混配對灰霉病都有較好的防效(蘇克躍 等,2013;楊勇 等,2016;吳祥 等,2017)。蛇床子素對白粉病防效突出(嚴清平 等,2005;李彩霞 等,2007),而目前登記用于防治灰霉病的蛇床子素產品僅限于與苦參堿的混配劑型。
20 世紀70 年代初,科研人員利用放射性土壤桿菌防治桃根癌病取得成功(Kerr &Htay,1974),此后植物病害的微生物防治不斷取得進展(孫廣正 等,2015)。用于防治灰霉病的微生物制劑主要利用生防菌的生長優勢,通過寄生和營養競爭作用,消耗侵染位點的營養物質,使灰霉病病原菌停止生長和侵染。一方面生防菌通過快速生長和繁殖奪取水分和養分,消耗氧氣,擠占生態位,壓縮同一生境中植物病原菌的生存空間;另一方面生防菌通過產生小分子的抗生素和大分子的抗菌蛋白或胞壁降解酶等來抑制病原菌的生長、繁殖和侵染(童蘊慧 等,2003)。木霉菌為病原菌的拮抗真菌,其作用機理復雜多樣。生產中木霉菌多用作土壤處理劑來調控農田微生態。木霉菌防治灰霉病時作為莖葉處理劑,防治時不能與其他真菌藥劑同時使用,也不能與強酸或強堿性物質混用。除木霉菌以外其他灰霉病的生防菌產品皆為細菌制劑,應用菌株主要集中在芽孢桿菌(Bacillus)和假單胞菌(Pseudomonas)。目前登記用于防治灰霉病的細菌制劑主要有枯草芽孢桿菌、海洋芽孢桿菌和甲基營養型芽孢桿菌9912 等3 種芽孢桿菌,以及熒光假單胞桿菌,劑型多為可濕性粉劑。采用菌劑防治灰霉病受產品貨架期、施用環境等因素影響較大。生物防治要獲得穩定的防治效果需建立在完善的病害綜合防治體系之上。微生物菌劑的應用是防治農作物灰霉病的重要補充,對化學農藥減量增效意義重大。
3 灰霉病的殺菌劑應用策略及抗藥性管理
化學防治起效快、防效明顯,能在短時間內高效迅速地控制病害的蔓延。但一方面由于灰霉病菌繁殖速度快、基因突變頻率高、適應性強,這極大地增加了抗藥性發生的概率;另一方面大面積種植單一品種作物、單作或連作的農事安排導致病害發生程度加重,過度依賴化學殺菌劑增加了藥劑選擇壓力和抗藥性發生風險。實際防治中,葡萄孢菌很容易對常用的化學殺菌劑產生抗性。抗性的發展進一步增加了化學殺菌劑的使用頻率,使灰霉病的化學防治陷入惡性循環,引發農藥濫用及農藥殘留超標等一系列問題。另外,殺菌劑的長期頻繁使用不可避免會破壞原生物群落結構,打破農業生態的自然穩態平衡,進一步造成灰霉病加重發生。
灰霉病的特點決定了其很難通過抗性育種手段解決防治問題。生態調控與生物防治雖然能一定程度上減少病菌侵染幾率,但田間一旦發病,化學防治仍是主要手段。化學防治建立在病原學的基礎上,以殺滅病原菌為主要目標。藥劑的殺菌效果是化學防治能否有效控制病情的關鍵。首先,為保證化學防治的效果,對于長期采取化學防控的地區,對田間優勢菌株定期進行抗藥性監測不可或缺。農藥主管部門應加大管控力度,引導農藥企業、農藥經營單位和農藥使用者共同參與抗性風險管理。對高、中等抗性風險的藥劑應規定年使用次數,在產品標簽上標注抗性風險級別和抗性風險管理措施。其次,農藥使用者要建立殺菌劑使用的田間檔案,注意不同作用機理的殺菌劑輪換使用,避免長期施用存在交互抗性的殺菌劑。植保推廣部門應建立混合用藥和輪換用藥的技術方案,推廣使用負交互抗性的藥劑。另外,隨著我國藍莓、獼猴桃、樹莓等新興漿果類水果種植面積的不斷增加,灰霉病的發生范圍也在不斷擴大。當前亟待加強新興作物及小宗作物的農藥登記,改變“非法”用藥的防治現狀。將灰霉病的化學防治納入科學規范的軌道上來。同時仍需積極推進灰霉病的綜合防治和綠色防控,通過建立完善的病害預測防治體系,進一步規范化學殺菌劑的使用,減緩抗藥性的發展。多措并舉,做好我國農作物灰霉病的防控工作。
