昆蟲抗藥性相關細胞色素P450基因的表達調控機制
2021-03-10 02:24:00邱星輝
昆蟲學報 2021年1期
朱 江,邱星輝
(1.中國科學院動物研究所,農業蟲害鼠害綜合治理研究國家重點實驗室,北京100101;2.中國科學院大學,北京100049)
人類為防治害蟲,相繼研發了有機氯、有機磷、氨基甲酸酯、擬除蟲菊酯、新煙堿以及二酰胺類等殺蟲藥劑。不同類型的殺蟲劑在植物保護和蟲媒疾病控制方面發揮重要作用的同時,也對地球上的生物帶來了新的壓力。昆蟲作為自然界種類極其豐富的生物類群,為應對殺蟲劑選擇壓,進化出多樣化的抗藥性機制,主要包括行為改變(行為抗性)、靶標的不敏感性(靶標抗性)、殺蟲劑表皮穿透能力下降(穿透抗性)以及殺蟲劑代謝解毒(代謝抗性)或扣押(sequestration)作用增強(Coetzee and Koekemoer,2013;Davidetal.,2013;Mugenzietal.,2019;Inghametal.,2020)。
細胞色素P450(簡稱P450或CYP)是存在于幾乎所有生物體中的蛋白超家族,其悠久的進化歷史(約35億年)賦予了其種類和功能多樣性的特征。昆蟲P450參與許多外源性化合物(簡稱外源物)的生物轉化,由其介導的殺蟲劑代謝解毒作用增強是昆蟲對不同類型殺蟲劑產生抗性和交互抗性的重要原因(Lietal.,2007;Feyereisenetal.,2015)。抗性昆蟲中P450的代謝解毒作用增強有兩類可能的機制,一是P450催化活性增強,二是P450酶量增加。催化活性增強的根源在于P450結構的變化,可能涉及氨基酸替換、選擇性拼接或形成嵌合體(Joussenetal.,2012;Ibrahimetal.,2015;Walshetal.,2018),而P450酶量增加涉及到P450基因拷貝數的增加以及P450基因的表達上調(Feyereisenetal.,2015)。
P450基因拷貝數增加涉及的分子事件包括基因復制(gene duplication)或基因擴增(gene amplification),即在基因組中產生一個或多個附加的P450基因(Emersonetal.,2008;Wondjietal.,2009;Puineanetal.,2010;Schmidtetal.,2010;Itokawaetal.,2011;Bariamietal.,2012;Kasaietal.,2014;Zimmeretal.,2018)。P450基因在轉錄層面的上調表達是P450介導抗藥性的最普遍機制(Fujii-Kuriyama and Mimura,2005;Lietal.,2007;Feyereisenetal.,2015),昆蟲抗藥性相關P450基因的轉錄上調由順式調控元件與反式作用因子相互作用得以實現 (Liuetal.,2015)。雖然目前人們對P450基因上調表達分子機制的認識還非常有限,但隨著組學與生物信息學的快速發展以及分子生物學與生物化學研究手段的革新,近年來P450基因調控機制的研究取得了重要的進展,本文試圖綜述這一領域的主要研究發現。
1 調控區DNA序列多態性與順式調控元件
真核生物的順式調控元件是指能與特定轉錄因子結合而調控基因轉錄的DNA序列。P450基因調控區存在著許多順式調控元件,包括啟動子、增強子、沉默子和轉座子(Chungetal.,2009)。P450基因的上調表達可能是由于調控區DNA序列的核苷酸插入、缺失或者堿基替換而增加調控元件與轉錄因子的結合能力,或產生新的調控元件(增強子、啟動子),或破壞沉默子的結合位點(Feyereisenetal.,2015;Liuetal.,2015)。
通過比較抗性與敏感等位基因調控區的序列多態性,已鑒定了一些可以導致P450基因過量表達的順式突變。黑腹果蠅DrosophilamelanogasterCyp6a2基因的過量表達與該基因5′側翼區存在完整的CncC:Maf轉錄因子結合位點相關聯(Misraetal.,2013),該結合位點的存在賦予Cyp6a2基因啟動子更強的轉錄活性(Wanetal.,2014)。家蠅Muscadomestica擬除蟲菊酯類抗性品系LPR的CYP6D1基因的5′側翼區包含一段15 bp的插入,該15 bp插入破壞了一個轉錄抑制子mdGfi-1(為C2H2型鋅指蛋白)的結合位點,從而降低了mdGfi-1對CYP6D1基因轉錄活性的抑制作用,導致CYP6D1 mRNA水平的增加(Gao and Scott,2006)。
桃蚜MyzuspersicaeCYP6CY3基因的過量表達與其啟動子區的微衛星AC(n)重復的長度有關(Bassetal.,2013)。有趣的是,在棉蚜Aphisgossypii和豌豆蚜Acyrthosiphonpisum的CYP6CY3同源基因的啟動子區也檢測到保守的AC(n)重復(Panetal.,2019)。在煙蚜Myzuspersicaenicotianae基因組中至少有5個CYP6CY3基因拷貝,這些CYP6CY3基因上游調控區DNA序列包含AC(n)重復,并表現出AC(n)重復長度的多態性,當AC(n)重復長度達到94 bp時,可以增強CYP6CY3的啟動子活性(Panetal.,2019)。致倦庫蚊Culexquinquefasciatus擬除蟲菊酯抗性相關的CYP9M10基因轉錄起始位點上游第27位的核苷酸發生了由鳥嘌呤向腺嘌呤的突變,由此增強了其轉錄活性(Itokawaetal.,2015)。褐飛虱Nilaparvatalugens吡蟲啉抗性相關基因CYP6ER1 (Bassetal.,2011;Liangetal.,2018)和CYP6AY1(Pangetal.,2014)的啟動子區DNA序列的改變與基因過量表達以及對吡蟲啉的抗性相關。Mugenzi 等 (2019)發現不吉按蚊AnophelesfunestusCYP6P9b基因的5′側翼區缺失3個堿基(ACC)與擬除蟲菊酯抗性以及擬除蟲菊酯殺蟲劑選擇壓相關聯。
轉座子(transposable elements,TEs),也稱跳躍基因(jumping gene),是順式調控元件的豐富來源(Villanueva-Caasetal.,2019;Branco and Chuong,2020)。TEs可以改變昆蟲基因組結構,影響基因的表達(Moschettietal.,2020)。外源物代謝相關的昆蟲P450基因往往擁有更豐富的轉座子(Chen and Li,2007)。有事例表明轉座子插入可以上調P450的表達而產生抗藥性。DDT抗性黑腹果蠅D.melanogasterCyp6g1基因的5′端非翻譯區存在一個轉座子插入[Accord,為長末端重復序列(long terminal repeat,LTR)類型],該轉座子的存在與DDT抗性呈正相關(Dabornetal.,2002)。轉座子Accord自身帶有轉錄增強子,使Cyp6g1基因在果蠅的中腸和脂肪體等組織中特異性過量表達(Chungetal.,2007)。Schmidt等(2010)在果蠅DDT抗性品系Cyp6g1調控區插入的轉座子Accord中發現了兩個附加的轉座子(P-element和HMS-Beagle),它們的存在進一步增強了Cyp6g1的表達。在加州擬果蠅DrosophilasimulansCYP6G1轉錄起始位點上游200 bp處插入一個全長為4 803 bp的非長末端重復(non-long terminal repeat,non-LTR)反轉錄轉座子Doc,也能導致Cyp6g1的上調(Schlenke and Begun,2004)。Salces-Ortiz等(2020)通過RNA-seq和ATAC-seq在基因組層面分析了4種果蠅品系(SE-Sto,RAL-375,RAL-177和iso-1)轉座子插入對馬拉硫磷應答基因表達調控的影響,發現位于應答基因附近的轉座子可能在外源物脅迫響應中發揮重要作用。
2 昆蟲P450基因表達的調控因子與信號轉導途徑
真核生物基因的轉錄調控受很多因子的影響,涉及的信號轉導途徑也多樣。以下介紹的是目前已鑒定的參與昆蟲P450基因表達的調控因子與信號轉導途徑。
2.1 CncC:Maf/Keap1
轉錄因子Nrf2(nuclear factor erythroid 2 p45-related factor 2)是屬于Cnc (cap‘n’collar) 堿性亮氨酸拉鏈家族(basic leucine zipper,b-ZIP)的轉錄活化子。在通常情況下,Nrf2與Keap1(Kelch-like ECH-associated protein 1,一種細胞骨架泛素連接酶)結合存在于細胞質中,而在應激情況下(如殺蟲劑暴露),Nrf2與Keap1解離,并轉移到細胞核中,與Maf(muscle aponeurosis fibromatosis,堿性亮氨酸拉鏈家族成員)形成雜合二聚體,并與基因上游特定的抗氧化物響應元件(antioxidant response element,ARE)結合而起始解毒酶(如P450、谷胱甘肽S-轉移酶等)基因的表達。CncC:Maf/Keap1調控途徑受到作為第二信使的活性氧(reactive oxygen species,ROS)的調控,ROS通過修飾Keap1中的特定半胱氨酸殘基而影響 Nrf2的細胞定位或泛素化,導致Nrf2 在細胞核中累積,活化外源物響應基因的表達(Taguchietal.,2011)(圖1)。
昆蟲的Nrf2直系同源物CncC (cap ‘n’ Collar isoform-C)已知在昆蟲生長發育和應激反應中發揮核心作用(Sykiotis and Bohmann,2008)。果蠅成蟲CncC的過量表達和dKeap1(DrosophilaKelch-like ECH-associated protein 1)的消耗可以活化許多防御相關基因的表達,而dKeap1過量表達則抑制這些基因的轉錄(Misraetal.,2011)。雙分子熒光互補分析發現,dKeap1通過與CncC的相互作用,形成的dKeap1-CncC復合物可以增強或抑制CncC的結合能力,因此對CncC活化某些基因的表達起增效作用,也可以拮抗CncC對另一些基因表達的活化,不同的外源物表現出對不同基因的選擇性表達調控(Deng and Kerppola,2014)。
至今已在許多昆蟲中發現了CncC:Maf/Keap1通路調控P450基因的表達(Palli,2020),表明這一表達調控途徑普遍被昆蟲利用以應對殺蟲劑的選擇壓。理論上CncC/Maf表達量上調、Keap1量的變化或這些蛋白結構的改變,以及ARE序列的差異都可以導致P450的表達增加,但直到目前還沒有發現因轉錄因子CncC,Maf或 Keap1突變導致P450介導的抗藥性的報道(Wilding,2018)。RNAi研究表明,敲低果蠅CncC可以下調Cyp6a2和Cyp6a8的表達,從而降低DDT抗性品系對DDT的耐受性(Misraetal.,2013)。吡蟲啉抗性馬鈴薯甲蟲CYP6BJa/b,CYP6BJ1v1,CYP9Z25和CYP9Z29的過量表達受CncC和MafS (small Maf)的調控(Kalsi and Palli,2017);敲低MafS可以降低岡比亞按蚊AnophelesgambiaeCYP6M2的表達,增加對擬除蟲菊酯殺蟲劑和DDT的敏感性(Inghametal.,2017)。桔小實蠅BactroceradorsalisCYP473A3的表達增加可以導致阿維菌素抗性,抗性品系中MafB (large Maf)過量表達,敲低CncC和MafB,減低了CYP473A3的表達和對阿維菌素的抗性水平(Tangetal.,2019)。在有機磷與擬除蟲菊酯抗性埃及伊蚊Aedesaegypti品系(Oiapoque),Nrf2的轉錄水平與敏感品系沒有差異,但Keap1,CYP6BB2以及GSTX2基因表現為過量表達;Nrf2表達沉默導致Keap1,CYP6BB2以及GSTX2表達量減少,暴露于馬拉硫磷的死亡率增加(Bottino-Rojasetal.,2018),該研究提出了一個Keap1表達與解毒酶基因表達以及抗藥性正相關的事例。Lu等(2020)發現ROS可以通過CncC:Maf/Keap1通路調控斜紋夜蛾SpodopteralituraCYP6AB12的表達。

圖1 CncC:Maf/Keap1-XRE信號通路調控昆蟲P450基因表達(改自Palli,2020)Fig.1 The CncC:Maf/Keap1 pathway involved in the expression regulation of insect P450 genes (adapted from Palli,2020)CncC:cap ‘n’ collar isoform-C;Keap1:Kelch樣環氧氯丙烷相關蛋白-1 Kelch-like ECH-associated protein 1;Maf:肌腱膜纖維瘤蛋白Muscle aponeurosis fibromatosis;ROS:活性氧Reactive oxygen species;XRE:外源物響應元件Xenobiotic responsive element.
2.2 GPCR/Gαs/AC/cAMP/PKA信號途徑
G蛋白偶聯受體(G-protein-coupled receptor,GPCR)是存在于細胞表面的膜結合蛋白,它通過與胞外信號的結合來調控胞內因子(如腺苷酸環化酶、磷脂酶和離子通道等),引發細胞響應而實現信號轉導。GPCR在配體的刺激下構象發生變化,接著與G蛋白(一種由α和β-γ亞基組成的異三聚鳥嘌呤核苷酸結合蛋白)偶聯,使得G蛋白的α亞基結合的二磷酸鳥苷(GDP)轉化為三磷酸鳥苷(GTP),導致α與β-γ亞基親和力降低而與β-γ亞基分離,進而分別與下游效應因子相互作用引發不同的胞內反應,從而轉導配體信號(Lappano and Maggiolini,2011)。GPCR的基本功能是控制G蛋白亞基的活性狀態,其中α亞基序列(Gα)根據其序列同源性和信號活性,分為Gαs,Gαi,Gαq/11和Gα12/13 4個家族(圖2)。

圖2 GPCR/Gαs/AC/cAMP-PKA通路調控P450基因表達(改自Lappano and Maggiolini,2011)Fig.2 The GPCR/Gαs/AC/cAMP-PKA pathway involved in the expression regulation of insect P450 genes (adapted from Lappano and Maggiolini,2011)AC:腺苷酸環化酶Adenylate cyclase;cAMP:環磷酸腺苷Cyclic adenosine monophosphate;GPCR:G蛋白偶聯受體G-protein-coupled receptor;GDP:二磷酸鳥苷Guanosine diphosphate;GTP:三磷酸鳥苷Guanosine triphosphate;Pi:磷酸Phosphate;PKA:蛋白激酶A Protein kinase A;RGS:G蛋白信號轉導調節子Regulators of G-protein signaling.α,β,γ,αs,αi,αq/11和α12/13均為蛋白亞基。α,β,γ,αs,αi,αq/11 and α12/13 are subunits of protein.
在致倦庫蚊抗性品系中發現4個GPCR (rhodopsin-like受體)基因與4個P450基因(CYP9M10,CYP9J34,CYP9J40和CYP6AA7)共同表達上調(Lietal.,2014)。對這4個GPCR基因進行RNAi處理,降低了致倦庫蚊對氯菊酯的抗性,同時也降低了2個依賴環磷酸腺苷(cyclic adenosine monophosphate,cAMP)的蛋白激酶A(protein kinase A,PKA)基因和4個抗性相關細胞色素P450基因的表達(Lietal.,2015)。采用cAMP抑制劑鹽酸布比卡因和RNAi技術,研究了其下游PKA在致倦庫蚊氯菊酯抗性中的作用,發現抑制GPCR信號通路第二信使cAMP和敲低PKA均能導致致倦庫蚊4個抗性相關P450基因和2個PKA基因表達水平以及對氯菊酯抗性程度的降低(Lietal.,2015)。這些結果提示GPCR參與了抗性相關的P450基因的表達調控。敲低GPCR信號通路下游G蛋白α亞基(Gαs)基因能降低腺苷酸環化酶(adenylate cyclase,AC)和PKA以及抗性P450基因的表達,而敲低AC只降低了PKA和抗性P450基因的表達(Li and Liu,2017),表明降低GPCR信號通路中任意一個基因的表達均能增加致倦庫蚊對氯菊酯的敏感性,同時也說明在GPCR通路中G蛋白α亞基(Gs alpha subunit protein,Gαs)位于AC的上游(Li and Liu,2017)。為明確GPCR通路中各個效應子的作用,Li和Liu(2019)在草地貪夜蛾SpodopterafrugiperdaSf9細胞中表達了致倦庫蚊的GPCR(GPCR020021)、Gαs(Gαs006458)、AC(AC007240)和PKA(PKA018257),發現隨著GPCR,Gαs或AC基因的表達增加,Sf9細胞cAMP產量增加,但cAMP的產量不因PKA表達的升高而變化。所有表達異源致倦庫蚊GPCR基因的Sf9細胞表現出更高的PKA活性和更高的CYP9A32表達水平,對氯菊酯耐受性也顯著提高;加入cAMP產生抑制劑(Bupivacaine HCl)和PKA活性抑制劑(H89 2HCl)可以顯著降低cAMP 濃度和PKA活性,并增加氯菊酯對細胞的毒性(Li and Liu,2019)。
赤擬谷盜TriboliumcastaneumCYP4BN6和CYP6BQ11的組成型過量表達或誘導表達可以降低赤擬谷盜對敵敵畏和呋喃丹的敏感性,誘導過表達和RNAi證據顯示這兩個基因的表達受蛛毒素受體(latrophilin,Tclph)的調控:敵敵畏和呋喃丹可以誘導Tclph以及CYP4BN6和CYP6BQ11,沉默Tclph的表達導致CYP4BN6和CYP6BQ11表達量下降,增加赤擬谷盜對敵敵畏和呋喃丹的敏感性(Gaoetal.,2018;Xiongetal.,2019)。最近,Ma等(2020)發現G蛋白偶聯受體Mthl10(LOC101899380) 在家蠅吡蟲啉抗性品系中過量表達,并在轉基因果蠅中證明了Mthl10在抗性相關P450基因的表達調控中發揮作用。
以上事例表明,昆蟲可以利用通用的GPCR/Gαs/AC/cAMP/PKA信號通路調控抗性相關的P450表達,但受該信號轉導途徑調控的、可活化上述P450基因表達的靶標轉錄因子尚未明確。
2.3 MAPK/ERK-p38/CREB信號通路
絲裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)是一類典型的絲氨酸/蘇氨酸類蛋白激酶,在真核生物中非常保守,作為細胞內多種信號通路,對于細胞周期的運行和基因的表達具有重要的調控作用(Cristinaetal.,2010)。MAPK級聯信號傳導是由3個進化非常保守的激酶所組成,即MAPK、MAPK激酶(MAPKK)和MAPKK激酶(MAPKKK)。在外界環境的刺激下,MAPKKK通過磷酸化與Ras/Rho家族一個小的GTP結合蛋白和/或其他蛋白相互作用而被激活,MAPKKK的活化進而引起MAPKK磷酸化而激活,接著引起MAPK的磷酸化,從而將MAPK與細胞表面的受體以及胞外的信號聯系到一起(Cargnello and Roux,2011)。在哺乳動物中主要有4種MAPK通路,即細胞外信號調節蛋白激酶(extracellular signal-regulated kinase 1/2,ERK1/2)、c-Jun氨基末端激酶(c-Jun N-terminal kinase 1/2/3,JNK1/2/3)、ERK5/BMK1和p38 (Cargnello and Roux,2011),它們可以被不同的外界條件所激活,形成不同的信號轉導(圖3)。
最近,Yang等(2020)發現MAPK/ERK-p38/CREB這一激酶激活的信號途徑參與調控煙粉虱BemisiatabaciCYP6CM1基因的表達。CYP6CM1的過量表達可以導致煙粉虱對吡蟲啉的抗性,而CYP6CM1的過量表達受bZIP超家族成員轉錄因子cAMP應答元件結合蛋白(cAMP-response element binding protein,CREB)的調控。絲氨酸MAPK途徑的ERK和p38信號使煙粉虱的CREB蛋白第111位的絲氨酸磷酸化,磷酸化的CREB與CYP6CM1基因啟動子區的CRE-like位點結合活化CYP6CM1的表達。在檢測的吡蟲啉抗性品系中p38和CREB 的表達水平較敏感品系高,敲低ERK或p38的表達,都可以降低CYP6CM1 mRNA和蛋白水平及煙粉虱對吡蟲啉的抗性。
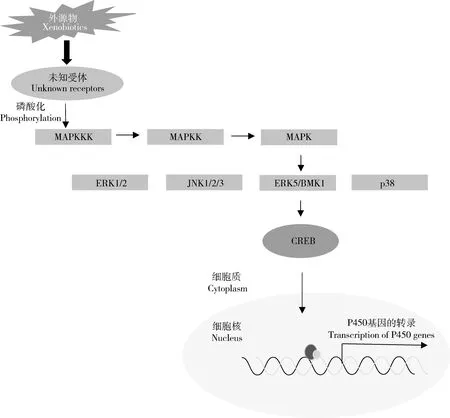
圖3 MAPK/ERK-p38/CREB通路調控昆蟲P450基因表達Fig.3 The MAPK/ERK-p38/CREB pathway involved in the expression regulation of insect P450 genesBMK:大絲裂原活化蛋白激酶Big mitogen-activated protein kinase;CREB:環磷腺苷應答元件結合蛋白cAMP-response element binding protein;ERK:細胞外信號調節激酶Extracellular signal-regulated kinase;JNK:c-Jun氨基末端激酶c-Jun N-terminal kinase;MAPK:絲裂原活化蛋白激酶Mitogen-activated protein kinase;MAPKKK:絲裂原活化蛋白激酶激酶激酶Mitogen-activated protein kinase kinase kinase;p38:p38絲裂原活化蛋白激酶p38 mitogen-activated protein kinase.
2.4 核受體
核受體是一類特殊的、古老的、配體依賴性轉錄因子超家族,具有保守的DNA結合結構域和配體結合結構域(Fahrbachetal.,2012),包括類固醇激素受體、非類固醇激素受體和孤兒核受體。在配體的作用下,核受體與共抑制子(co-repressor)解離并開始招募共激活子(co-activator),然后與靶基因上游的響應元件結合(King-Jones and Thummel,2005),啟動P450基因的轉錄(圖4)。
芳烴受體(aryl hydrocarbon receptor,AhR)是一種配體(如花椒毒素等外源物)激活的轉錄因子,AhR及其芳烴受體核轉運蛋白(AhR nuclear transportor,ARNT)隸屬于含bHLH結構域以及PAS結構域的轉錄因子家族(Guetal.,2000;Fujii-Kuriyama and Mimura,2005)。在未與配體結合的情況下,大部分AhR位于細胞質中,與熱激蛋白90(heat shock protein 90,hsp90)以及熱激蛋白90關聯蛋白(hsp90-associated proteins)結合,形成復合物,使AhR維持在一種非活化可與配體結合的狀態(Berg and Pongratz,2001)。配體與AhR結合后,AhR從復合物中游離出來并且轉移至細胞核中,與ARNT形成異源二聚體。異源二聚體再與P450基因啟動子上特有的XRE-AhR元件結合(Reyesetal.,1992),隨后誘導P450基因的表達。在昆蟲中,花椒毒素通過AhR信號轉導途徑誘導香芹黑鳳蝶PapiliopolyxenesCYP6B1 (Brownetal.,2005)和虎鳳蝶PapilioglaucusCYP6B4 (McDonnelletal.,2004)的表達。在埃及伊蚊氯菊酯誘導的CYP6家族基因的啟動子區也發現了類似于外源物響應元件 (xenobiotic responsive element,XRE)的轉錄因子結合位點(Poupardinetal.,2008)。基于RNAi和轉錄因子共轉染實驗結果表明棉蚜CYP6AD2 (Pengetal.,2016)和煙蚜CYP6CY3 (Panetal.,2019)的表達受轉錄因子AhR/ARNT的調控,具體表現為通過飼喂雙鏈RNA (dsRNA)對AhR和ARNT基因干擾,CYP6AD2或CYP6CY3表達顯著降低,在sf9細胞中共轉染AhR和ARNT基因可以增強CYP6AD2或CYP6CY3基因的啟動子活性 (Panetal.,2019)。
DDT抗性果蠅品系差異表達基因的轉錄因子結合位點分析發現,共調控的多個基因都存在糖皮質激素受體(glucocorticoid receptor,GR)結合位點(Qiuetal.,2013)。在DDT抗性果蠅中發現了GR同源物雌激素相關受體(estrogen-related receptor,ERR)存在一個甘氨酸插入突變,導致果蠅Cyp6g2,Cyp9c1和Cyp12d1基因的組成型上調表達(Sunetal.,2015),這一結果表明核受體ERR在果蠅中參與某些P450的表達調控。
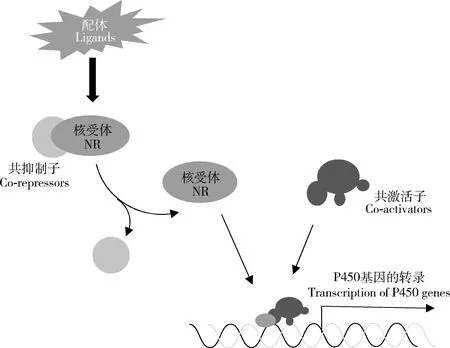
圖4 核受體調控昆蟲P450基因表達Fig.4 The nuclear receptor (NR) involved in the expression regulation of insect P450 genes
果蠅核激素受體96(nuclear hormone receptor 96,DHR96)與哺乳動物的孕烷X受體-組成型雄甾烷受體(pregnane X receptor-constitutive androstane receptor,PXP)是同源物(King-Jones and Thummel,2005)。研究表明,由DHR96介導的Cyp6g1(以及谷胱甘肽S-轉移酶GstE1)上調表達可以增強果蠅對DDT的抗性(King-Jonesetal.,2006;Afscharetal.,2016)。DHR96功能缺失的果蠅對DDT敏感(King-Jonesetal.,2006;Afscharetal.,2016),而DHR96在馬氏管中過量表達可以提高果蠅的DDT抗性(Afscharetal.,2016)。
Li等(2019)發現,小菜蛾Plutellaxylostella的孤兒核受體FTZ-F1和CYP6BG1在抗性種群中過量表達,且FTZ-F1和CYP6BG1的表達可以被氯蟲苯甲酰胺誘導。進一步研究發現CYP6BG1基因的轉錄起始位點上游存在FTZ-F1的結合位點,往3齡幼蟲注射dsRNA干擾FTZ-F1的表達也導致CYP6BG1表達量的下降(Lietal.,2019),這些實驗結果表明CYP6BG1的表達受到FTZ-F1的調控。
3 昆蟲P450表達的轉錄后調控機制
轉錄后調控是指真核生物的mRNA前體到成熟mRNA翻譯起始前這一過程中的調控,包括mRNA剪切、編輯、轉運和穩定性維持。近年來,非編碼RNA對昆蟲P450的調控作用的研究受到了學者越來越多的青睞。
小RNA (microRNA,miRNA)是一類長度大約為22個核苷酸的小非編碼RNA,它通過其5′端的長度為2~8個核苷酸的種子序列(seed sequence)與靶基因mRNA的3′端非翻譯區(untranslated region,3′UTR)互補匹配位點結合,導致mRNA降解或翻譯抑制,由此調控許多基因的表達(Lucas and Raikhel,2013)。miRNA最有效的作用靶標是受體、激酶和轉錄因子等信號級聯反應的成員,也有一些研究表明,miRNA可以調控抗藥性相關的P450的表達。尖音庫蚊CulexpipiensmiR-71(Hongetal.,2014)、miR-285(Tianetal.,2016)和miR-13664(Sunetal.,2019)分別通過調控靶基因CYP325BG3,CYP6N23和CYP314A1的表達而改變對溴氰菊酯的抗性。miRNA可能以簇的方式存在,同簇miRNA之間存在某種方式的內在調節機制,以網絡調控的方式改變抗性相關的P450的表達。例如在擬除蟲菊酯類殺蟲劑抗性的淡色庫蚊Culexpipienspallens雌性成蟲中,MiR-2/13/71簇表達下調,由此調控CYP9J35和CYP325BG3的表達而導致抗藥性,其中MiR-2/13/71簇的miR-2或 miR-13可以直接調控CYP9J35的表達和對溴氰菊酯的抗性(Guoetal.,2017)。煙蚜CYP6CY3的表達及煙蚜對尼古丁的抗性受到let-7和miR-100的調控(Pengetal.,2016),而miR-2b-3p可能降低小菜蛾CYP9F2 mRNA的表達水平(Etebarietal.,2018)。
除miRNA外,其他非編碼RNA也可能參與P450的調控。例如在尖音庫蚊中,piRNA(piwi-interacting RNA)-3878可以靶向CYP307B1來調控擬除蟲菊酯殺蟲劑抗性(Yeetal.,2017);小菜蛾氯蟲苯甲酰胺抗性品系中CYP6B6-like基因的過量表達與11種長鏈非編碼RNA (lncRNA,長度大于200 bp)的表達相關聯,但這些lncRNAs是如何調控CYP6B6-like基因表達的尚不清楚(Zhuetal.,2017)。
4 昆蟲P450基因表達的表觀遺傳調控
哺乳動物的P450的表達受DNA甲基化和組蛋白翻譯后修飾的調控,如CYP2C19的表達受DNA甲基化的間接調控(Burnsetal.,2018),丙戊酸誘導的組蛋白乙酰化可抑制CYP19A1基因的表達(Chenetal.,2015)。有證據表明酯酶E4基因的DNA甲基化在桃蚜抗藥性中發揮作用(Fieldetal.,1989;Hicketal.,1996;Field,2000),但至今有關表觀遺傳調控昆蟲抗藥性相關P450表達的報道很少。Mahmood等(2016)分析了家蠅多殺菌素抗性品系中差異表達的8個P450基因(CYP4G2,CYP6A1,CYP6A36,CYP6A37,CYP6D1,CYP6D3,CYP6G4和CYP12A2)的CpG島的頻數、所在位置和長度,發現其中7個P450存在1個CpG島位點,而CYP12A2包含3個CpG島。不同P450基因的CpG島所在位置差異很大,CYP6A1,CYP6A36、CYP6A37和CYP6D3包含一個靠近5′區的CpG島,CYP4G2和CYP12A2的 CpG島靠近3′區,而CYP6G4的一個CpG島位于基因序列的中間,為外顯子區;3個與外源物代謝相關的P450基因(CYP6A1,CYP6D1和CYP6G4)具有一個相對更大的CpG島。用組蛋白脫乙酰酶抑制劑可以誘導西方蜜蜂ApismelliferaP450基因(CYP9和CYP4G11)的表達,增強對吡蟲啉的耐受性(Huetal.,2017)。
5 結語
雖然殺蟲劑化學結構多樣、作用靶標和作用方式有所不同,但在使用一段時間后,昆蟲毫無例外地能產生抗藥性(Denholmetal.,2002)。通過P450的過量表達以增強對殺蟲劑的代謝解毒作用是昆蟲應對殺蟲劑脅迫使其種群保存下來的一個重要途徑。抗性昆蟲P450基因的過量表達可以是基因組DNA層面的基因重復與擴增,更普遍的是轉錄層面的表達上調。抗性昆蟲P450轉錄上調機制復雜,可以是順式調控元件、反式作用因子,或者順式調控元件和反式作用因子共同作用的結果。現已發現幾種不同類型的轉錄因子(如CncC、CREB和核受體等)和miRNA對昆蟲P450表達的直接調控,也鑒定了間接調控P450表達的作用因子如GPCR及其下游效應子。現有的研究結果充分顯示昆蟲P450基因調控因子和信號轉導通路的多樣性和調控機制的復雜性,一些P450的表達調控甚至是多層面多途徑的。外源物應激響應相關的轉錄因子CncC在調控解毒酶基因表達的中心地位及其在抗藥性形成中的作用已得到很多事例的支持,其他參與抗藥性調控的因子還有待發現。
在基因表達調控水平上,探討昆蟲發生什么遺傳突變,利用何種調控因子或非編碼RNA,以及經由哪些信號途徑而獲得抗藥性,是值得并需要加強的研究方向。通過對P450轉錄調控機制的深入研究,不僅為認識昆蟲適應性的進化機制提供新視角,還可以發現用于代謝抗性檢測的分子標記以及害蟲防治的新靶標,為昆蟲抗藥性監測、治理以及害蟲可持續有效防控提供新的思路與手段。可以相信,綜合采用比較基因組與轉錄組、全基因組關聯分析(genome-wide association studies,GWAS)、染色質免疫共沉淀-測序(ChIP-Seq)、RNAi和規律間隔成簇短回文重復序列(clustered regularly interspaced short palindromic repeats,CRISPR) 基因編輯等現代生物方法與技術,必將在更多的抗性昆蟲中發現更多的調控元件、調控因子和調控途徑,對P450表達調控機制有新的更全面的認識。
