小興安嶺天然次生林不同林型下紅松種源試驗
2021-03-02 03:53:06魏志剛夏德安王瑞琪劉瑩瑩李若林楊傳平
植物研究 2021年5期
關鍵詞:差異
魏志剛 夏德安 王瑞琪 張 洋 劉瑩瑩 李若林 楊傳平*
(1. 國家林業與草原局鹽堿地研究中心,中國林業科學研究院,北京 100091;2. 林木遺傳育種國家重點實驗室,東北林業大學,哈爾濱150040;3. 麗林實驗林場,伊春 153031)
闊葉紅松(Pinus koraiensis)林是小興安嶺區域內頂級地帶性群落[1]。同時,闊葉紅松林中主要樹種紅松、水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica)、黃 菠 蘿(Phellodendron amurense)等均為珍貴用材樹種[1],因此它也是我國重要的木材資源儲備基地[2~3]。新中國成立58年間,小興安嶺共生產木材3.28 億立方米,每年采伐量為生長量的2.0~2.5 倍,同時由于森林保護措施不力、毀林開荒、森林火災和森林病蟲害等原因,導致該區域闊葉紅松林資源已近枯竭,紅松蓄積量目前僅存400 多萬立方米[2,4~5]。原始闊葉紅松林破壞后,可衍生出不同類型的天然次生林,如楊樺林[主要由山楊(Populus davidiana)和白樺(Betula platyphylla)組成]、硬闊林[主要由水曲柳、胡桃楸、黃菠蘿、榆樹(Ulmus pumila)、色木(Acer pictum)等組成]和雜木林[主要由毛赤楊(Alnus hirsuta)、白樺和落葉松(Larix gmelinii)等組成]等[4,6]。與原始闊葉紅松林相比,天然次生林遺傳多樣性下降、生態功能脆弱和生產力低下[4,7],已無法滿足該區域社會經濟可持續發展和生態環境安全的基本需求[4]。因此,如何提高現有天然次生林的生產力和生態保護功能,已成為區域內森林經營的首要問題[3,8~9]。研究表明,天然次生林是闊葉紅松林中建群樹種紅松遭到毀滅性破壞演化形成[10~12],通過“栽針保闊”等提高次生林中紅松的優勢度的經營措施可促使其向生長力高和穩定性好的原始闊葉紅松林狀態恢復[9,13~19]。因此,通過天然次生林下間隙和斑塊栽植紅松已成為該區域天然次生森林經營的主要任務[17,19~21]。
不同類型的天然次生林,由于樹種組成不同,其林下土壤理化特性、光照和植被類型均存在較大差異[11,22~24],因此會對造林后紅松的生長產生不同的影響[19,25]。以往研究中,對天然次生林下光照、氣候與土壤因子對紅松細苗的生長等性狀進行過一系列研究[22,26~27]。長期以來,天然次生林下造林用的紅松苗木均為皆伐地選育出的良種或品種[28~30],在環境更為復雜的次生林林冠下其生長與適應性可能會產生嚴重分化,從而對天然次生林向闊葉紅松林的恢復進程造成嚴重影響。然而,目前尚無天然次林下造林用紅松良種選育的研究報道。針對上述問題,本項研究在前期皆伐地紅松種源試驗研究結果的基礎上[28~30],通過分析小興安嶺北部麗林實驗林場3 種主要天然次生林林型對紅松各種源生長與適應性狀的影響,為不同類型的天然次生林林下紅松造林選擇出優良紅松種源。本項研究結果對于今后東北地區天然次生林“栽針保闊”時紅松良種的選擇提供了理論依據。
1 材料與方法
1.1 種源材料
在課題組前期“六五”轉“七五”科技攻關課題紅松種源試驗的基礎上[28~30],選擇在小興安嶺地區表現較好的10 個紅松種源,分別是黑龍江省帶嶺、金山屯、鐵力、黑河、鶴崗、湯原,吉林省的敦化、白河、汪清與遼寧省的本溪種源。
1.2 造林地點與林型
1.2.1 造林地的自然情況
造林地位于小興安嶺南坡北部的麗林實驗林場,湯旺河上游林區。全場地形屬丘陵西北高而東南低,海拔在30~50 m,山脈地勢平緩,坡度為10°~20°。屬大陸性氣候并兼有季風氣候特點。年溫差達4℃、年平均氣溫在0~2℃、年降水量為550~600 mL,多集中在7~9 月。常風方向為西南風,年平均風速3 而秒。土壤分布在海拔30~50 m 范圍內,紅松、云冷杉林地土壤,以暗棕壤為主,多分布在陡、斜、緩坡地,個別地有腐殖土。在海拔300 m 以上的落葉松、白樺林下的土壤,以沼澤化暗棕壤及沼澤土類為主,都分布在平地或谷地。
1.2.2 不同林型的基本情況
在小興安嶺地區麗林實驗林場選擇了楊樺林、雜木林和硬闊林進行林下紅松種源試驗,各林型具體情況如表1所示。
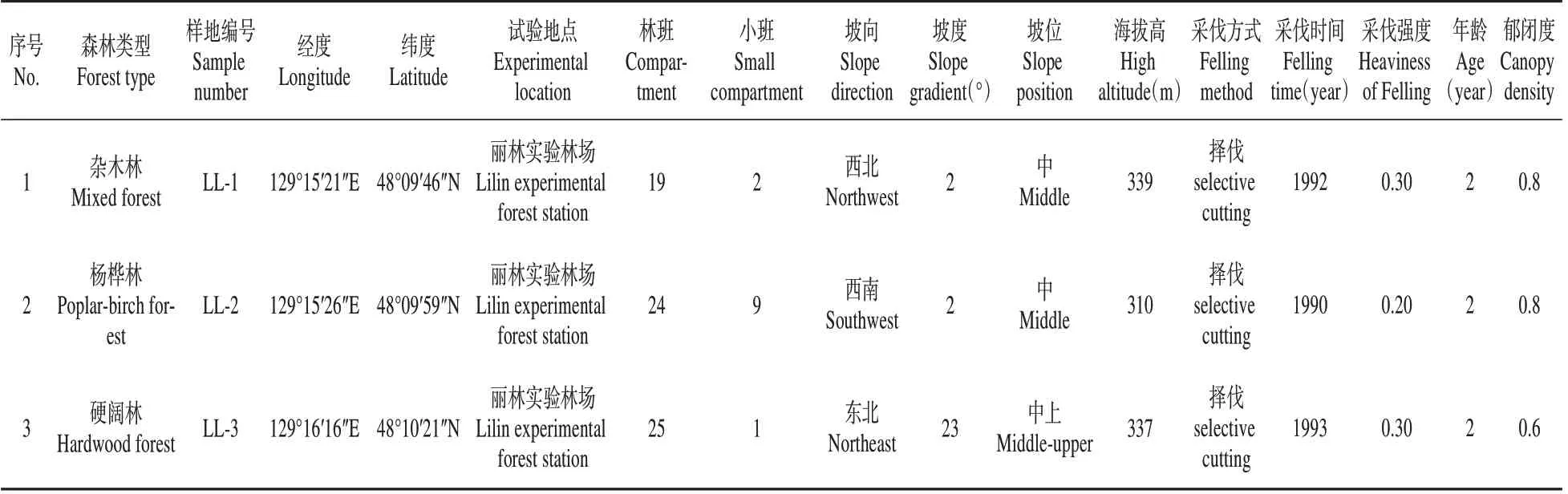
表1 3種天然次生林的基本情況Table 1 Basic information of the three natural secondary forests
1.3 各試驗點造林方案
造林均采用隨機完全區組設計,10 次重復,根據試驗地面積大小確定小區株樹為10~20 株,株行距為2 m×2 m,單行或者雙行小區。區組間用1 行紅皮云杉做隔離行,試驗地四周用1 行紅皮云杉做保護行,保護行和隔離行的株行距與紅松相同。各種源苗木為均為4 年生,2018 年春季造林。
1.4 性狀調查與統計分析
2020 年10 月,林木停止生長后,對各林型下10 個區組內的全部個體進行地徑、幼樹高(7 年生總高)、當年高(2019 年10 月至2020 年10 期間的高生長)、3 年高(造林后3 年期間的高生長)和成活率性狀調查。
在統計分析前,依據拉依達準則去除異常數據[32],并進行以下參數估算。
表型變異系數:
式中:σP為表型標準差,為群體平均值。
種源遺傳力:
式中:MSf為種源均方,MSe為誤差均方,F為方差方差分析中F值。
遺傳增益:
式中:S為選擇差,h2為遺傳力,為某一性狀的均值[33]。
數據計算采用Minitab和SAS軟件進行性狀基本統計、方差分析和多重比較。
2 結果與分析
2.1 不同林型下紅松種源幼林期性狀變異分析
各林型下紅松造林3 年后生長與適應性狀統計分析結果(見表2)表明:楊樺林下紅松各種源地徑、樹高、當年高、3年高和存活率的性狀與變異系數 均 值 分 別 為0.72 cm 和12.42%、0.36 m 和13.52%、0.09 m 和24.29%、0.20 m 和25.17%、48.63%和51.25%;硬闊林下試點各種源上述性狀分別為0.69 cm和14.63%、0.34 m和20.86%、0.08 m和38.80%、0.16 m 和28.63%、19.25%和149.98%;雜木林下各種源上述性狀分別為0.62 cm 和15.66%、0.29 m 和20.46%、0.06 m 和28.93%、0.17 m 和25.17%、18.85%和76.42%。3 種林型對紅松種源性狀的影響不同,其中,楊樺林、硬闊林和雜木林下各性狀變異系數均值分別為25.33%、50.58%和33.33%,表明硬闊林林下環境復雜多變,對紅松生長適應性狀影響較大,而楊樺林林下紅松生長環境相對均一,因此紅松各性狀變異較小。同時,不同林型下,紅松各性狀均產生了較大變異,這為選擇適應好的種源提供了基礎。
2.2 不同林型下紅松種源幼林期性狀差異分析
方差分析發現:不同林型下,紅松種源各性狀存在極顯著差異;各林型下,除3 年高性狀種源間差異不顯著外,其余性狀種源間差異極顯著;林型與種源之間在樹高和當年高性狀上交互作用顯著(見表3)。上述結果表明,小興安嶺麗林實驗林場天然次生林下紅松造林時,必須考慮林型和種源因素對紅松造林效果的影響。
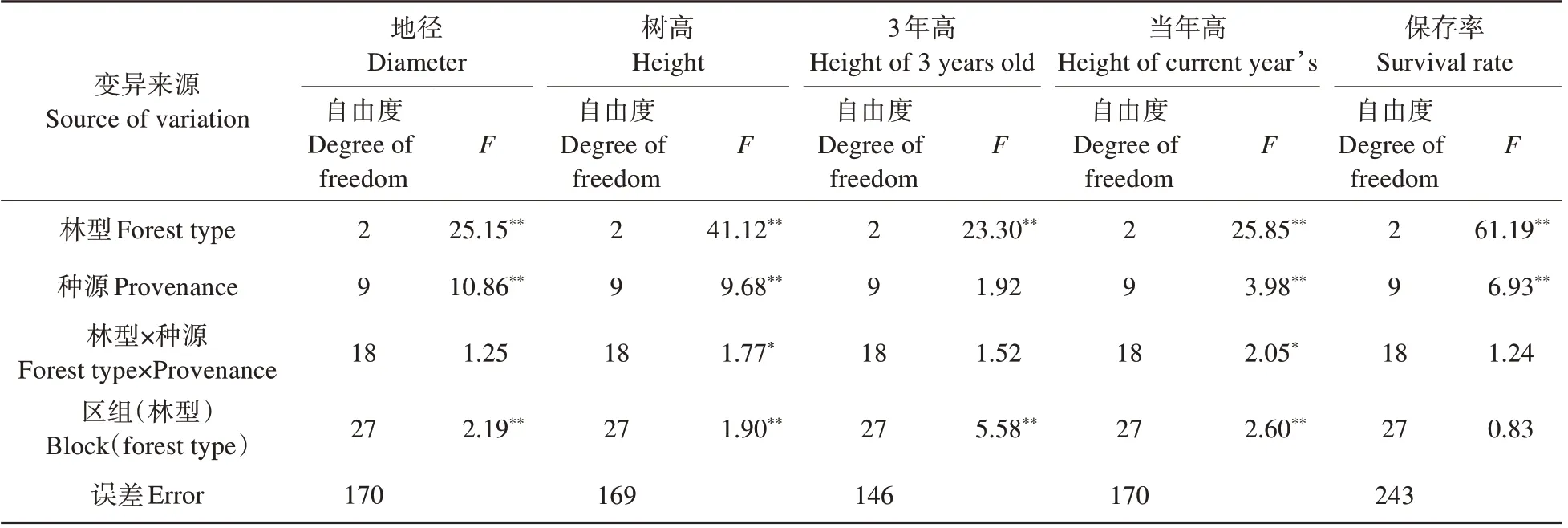
表3 不同林型下紅松幼林期各種源生長與適應性狀的方差分析結果Table 3 Variance analysis results of growth and adaptation traits of various sources in young stand of Korean pine with different forest types
2.3 適宜紅松造林林型與各林型的優良種源選擇
不同林型間紅松各性狀多重比較發現:楊樺林下紅松種源各性狀表現最好,是最適合紅松造林的林型;其次為硬闊林;雜木林下紅松性狀表現最差(見表4)。上述結果表明,有必要為不同林型分別選擇紅松優良種源。
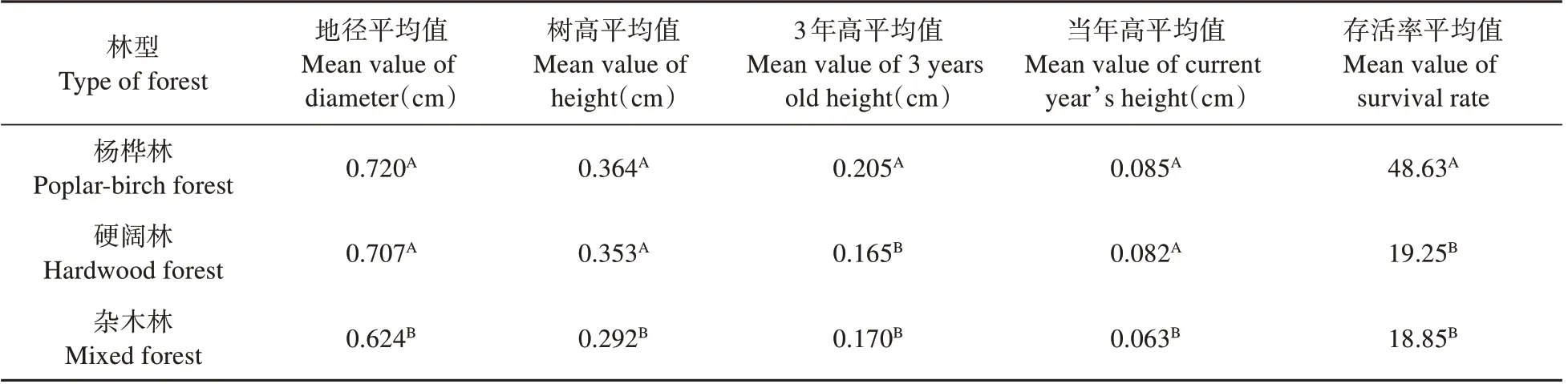
表4 不同林型下紅松幼林期種源生長與適應性狀的多重比較結果Table 4 Multiple comparison results of provenance growth and adaptive traits in young stage of Korean pine under different forest types
不同林型下紅松種源生長與適應性狀方差分析發現:楊樺林下,各種源間3 年高性狀各種源間差異不顯著,當年高性狀差異顯著,其余性狀均存在極顯著差異;硬闊林下,各種源間地徑性狀差異極顯著,而其他性狀各種源間差異不顯著;雜木林下,紅松種源間上述性狀均存在極顯著差異(見表5)。
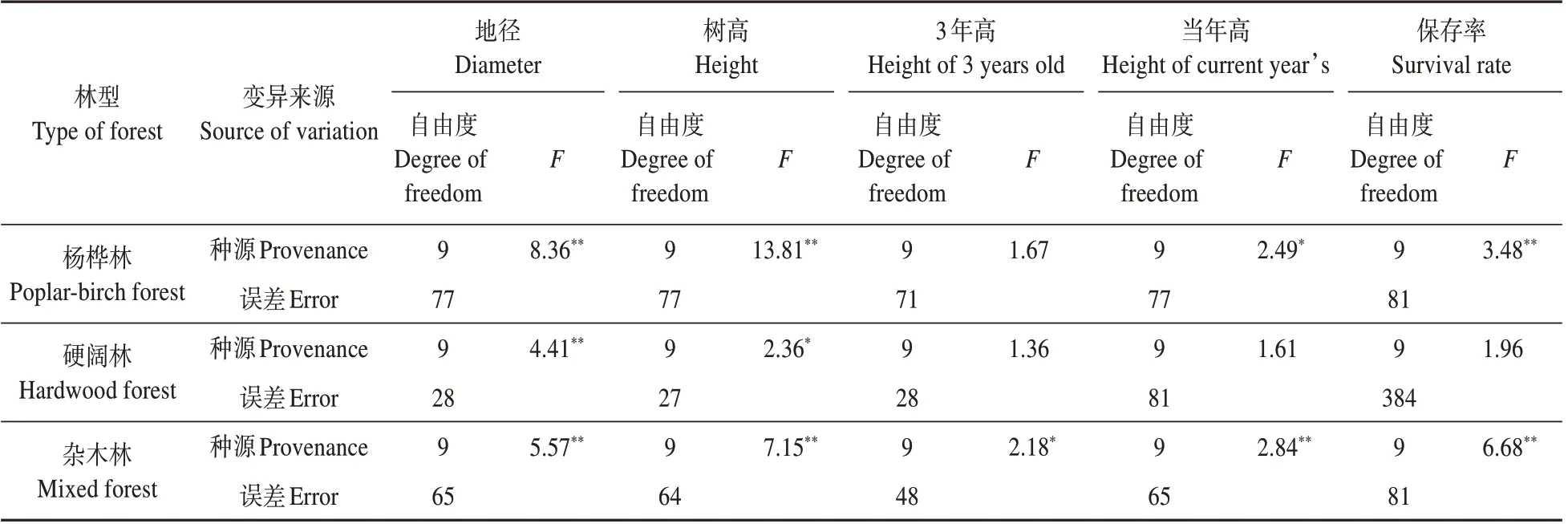
表5 不同林型下紅松幼林期生長與適應性狀的方差分析結果Table 5 Variance analysis results of growth and adaptation traits of young Korean pine under different forest types
種源間差異顯著性狀遺傳力計算分析發現:楊樺林下,種源間存在極顯著和顯著差異性狀的遺傳力分別為0.88、0.93、0.60 和0.71;硬闊林下,種源間存在極顯著和顯著差異性狀的遺傳力分別為0.92 和0.94;雜木林下,種源間存在極顯著差異性狀的遺傳力分別為0.82、0.86、0.54、0.65和0.85。上述結果表明,紅松種源間各性狀的變異受遺傳因素較大,屬中強度遺傳性狀,通過選擇能獲得較好的遺傳增益。同時,不同林型下,同一性狀力遺傳力存在一定差異,進一步證明遺傳力性狀是特定條件下的估算參數。
楊樺林下,種源間存在顯著和極顯著差異性狀多重比較結果(見表6)發現:鐵力種源總體表現最優,其地徑、樹高和當年高分別超出對照(各種源均值)10.00%、21.17%、和12.67%。其保存率雖然不是最高,但與表現最優的金山屯種源間無顯著差異。在楊樺林下選擇該種源造林,在地徑、樹高和當年高性狀的遺傳增益分別為8.8%、19.7%和7.61%。
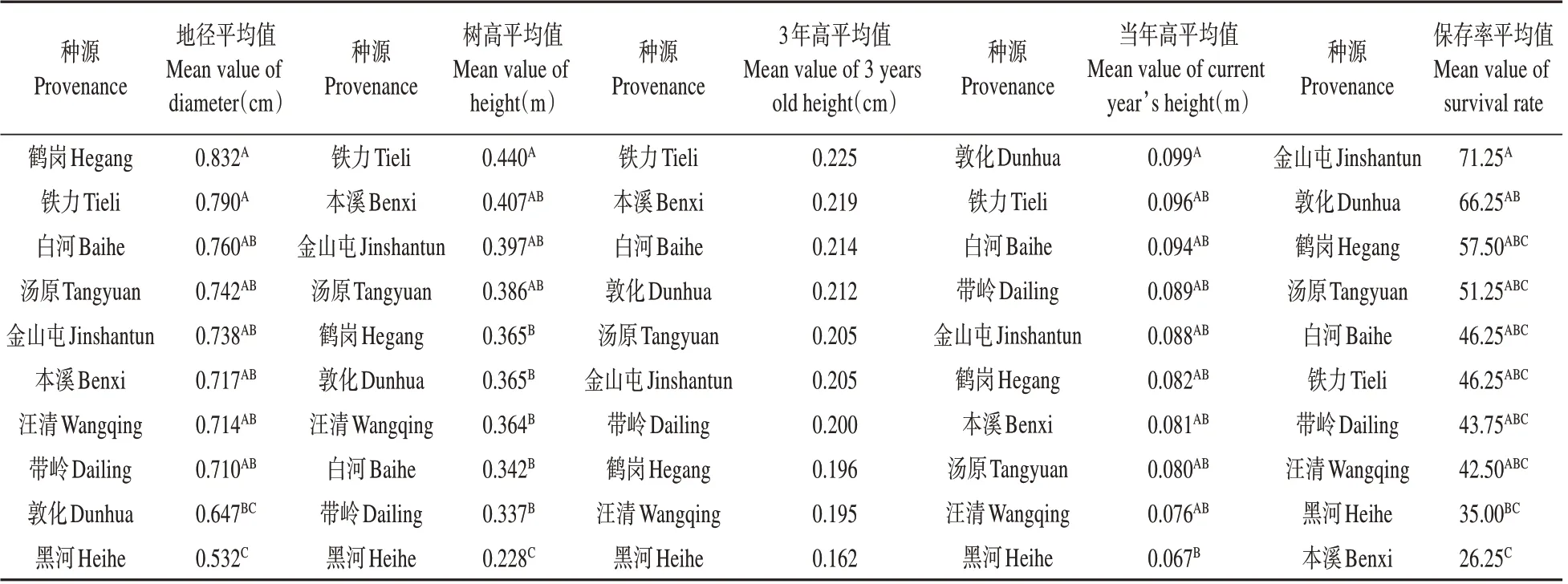
表6 楊樺林下紅松幼林期各種源差異顯著性狀間的多重比較Table 6 Multiple comparison of significant characters with different sources in young stage of Korean pine under poplar and birch forests
硬闊林下,汪清種源性狀綜合表現最好(見表7),其地徑和當年高分別超出對照(各種源均值)24.90%和8.69%,其造林后在上述2個性狀上的遺傳增益分別為22.92%和21.35%。
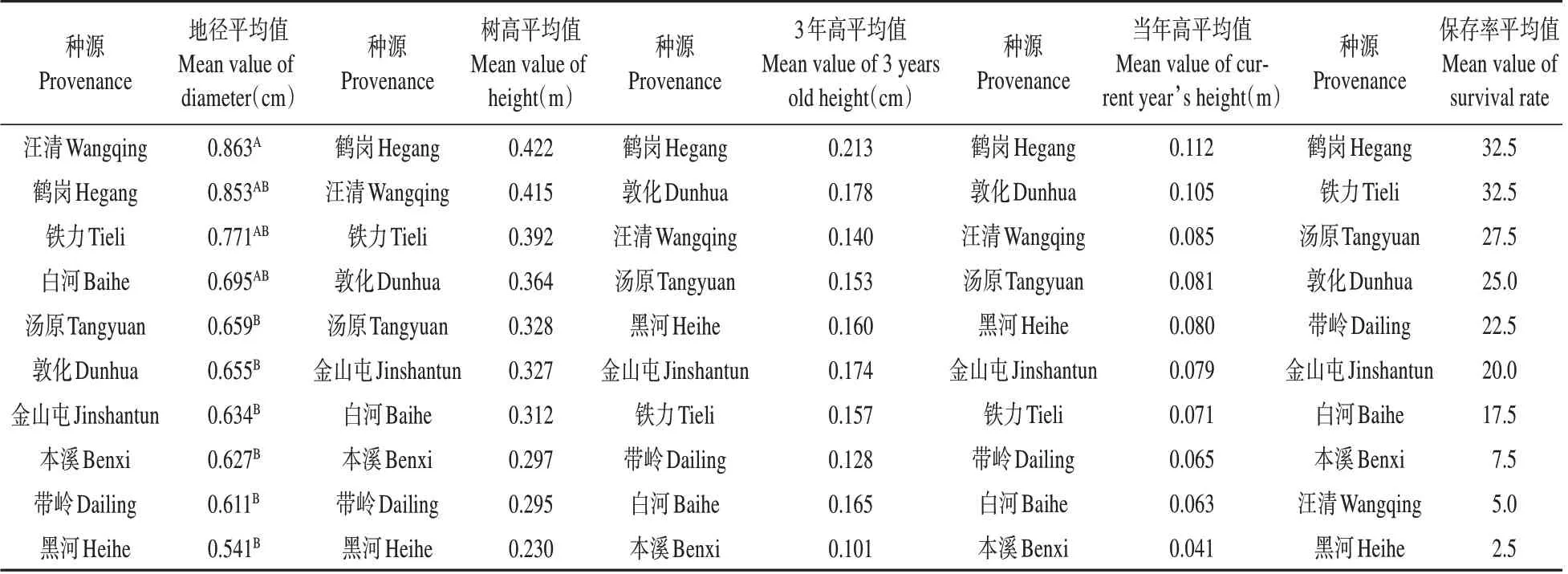
表7 硬闊林下紅松幼林期各種源間差異性狀的多重比較Table 7 Multiple comparison of different characters among different sources in young stand stage of Korean pine under hard broadleaf forest
雜木林下,鐵力種源表現最好(見表8),其地徑、樹高、當年高和保存率分別超出對照(各種源均值)12.42%、29.49%、10.75%和16.71%,其造林后在上述性狀的遺傳增益分別為10.19%、25.36%、6.99%和14.20%。
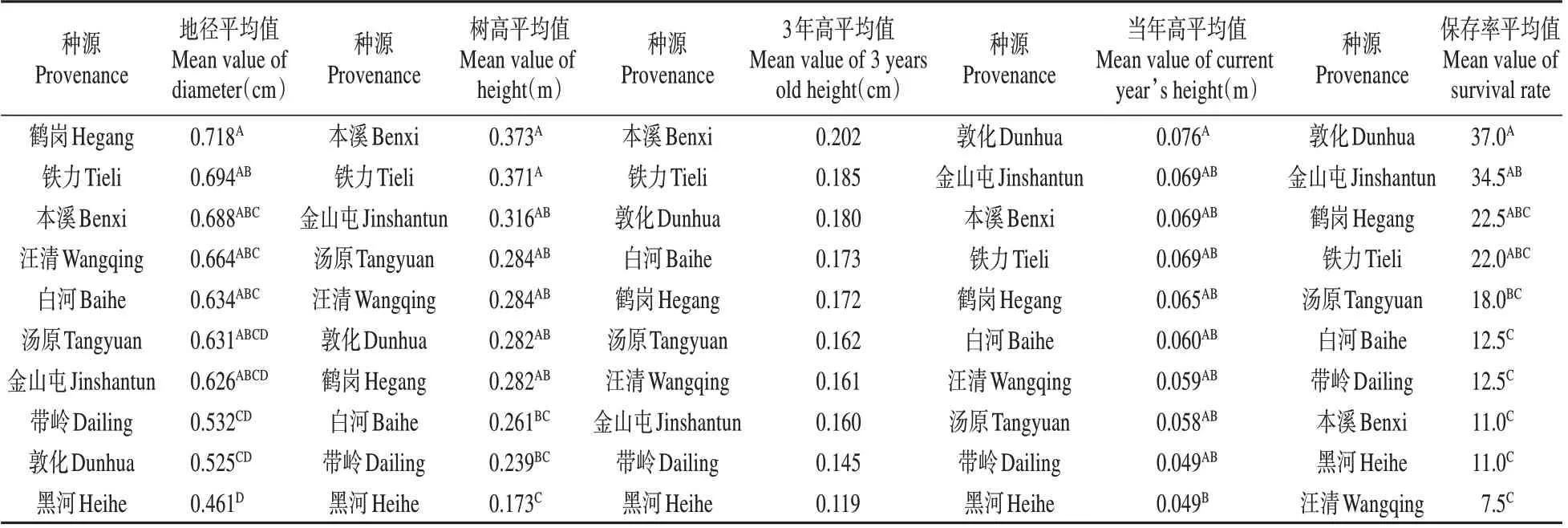
表8 雜木林下紅松幼林期各種源差異性狀多重比較Table 8 Multiple comparison of various sources of different characters in the young stage of Korean pine under miscellaneous forest
3 討論
長期以來,小興安嶺地區天然次生林下栽植選擇的紅松幼苗均是基于前期皆伐地的紅松種源試驗或家系試驗選育出的良種,這些苗木沒有進行過林下更為復雜生長環境的遺傳測定,因此生長過程中性狀可能會產生嚴重分化,從而會對天然次生林的恢復進程造成嚴重影響。因此,開展天然次生林下紅松的良種選育研究工作勢在必行。紅松在我國東北地區主要分布于長白山到小興安嶺一帶,呈新月型走向,并且分布高度隨緯度增高而下降,分布區內不同群體間存在極為廣泛的遺傳變異[28],為不同類型天然次生林下紅松優良種源的選擇提供了豐富的群體育種。
本項研究對不同林型下紅松幼林期各種源性狀基本統計分析結果(見表2)發現,林型對紅松各種源幼林期性狀影響不同。其中,對適應性狀影響最大,其次為3年高、當年高和樹高性狀,而對地徑的影響最小。同時,紅松各種源幼林期性狀在不同林型下的變異幅度不同,其中,硬闊林對各種源性狀總體上影響最大,其次為雜木林,最后為楊樺林。方差分析結果進一步證實,林型對紅松各種源多數性狀的影響達到了極顯著水平,且林型與種源在樹高和當年高性狀上交互作用顯著(見表3)。林型對紅松種源生長與適應性狀產生不同影響的原因可能由于不同林型的郁閉度、林隙大小、小氣候特征和土壤的物理性狀等因素不同[22~24],如不同林型光環境對紅松生長性狀具有顯著影響。然而,上述因素對紅松種源性狀影響的具體機制尚需進一步研究。
從各林型下紅松各性狀的多重比較與綜合表現來看,楊樺林下紅松性狀表現最好,其次為硬闊林,而雜木林最差(見表4)。這可能與楊樺林水源涵養功能最強有關。同時,麗林林場天然次生林下營造紅松時,應盡可能在楊樺林下造林,這樣能保證獲得較高的造林收益。同一林型下,紅松種源生長與適應性狀方差分析發現,各種源間存在極顯著或顯著差異(見表5)。上述結果進一步表明,由于紅松不同種源的遺傳基礎不同,因此同一林型下性狀會產生明顯分化。這一結果與皆伐跡地上紅松各種源生長性狀存在極顯著差異一致[28~30]。因此,必須針對不同林型選擇適宜種源才能保證“適地適樹”的造林法則在營林實踐生產中的落實。
不同林型下具有顯著和極顯著差異的性狀遺傳力不同且多為中強度遺傳性狀,其中,地徑和樹高性狀在3種林型下均具有較高遺傳力,當年高、3年高和存活率性遺傳力較低。在紅松半同胞家系選擇研究中,也發現紅松地徑和樹高性狀遺傳力均超過0.5,但具體值與本項研究結果不同。上述結果進一步證實遺傳力性狀是特定條件下的估算參數。
在10%的入選率條件下,不同林型下紅松優良種源不同且不同性狀獲得的遺傳增益也不同,其中:楊樺林下,鐵力種源表現最好,造林后地徑、樹高和當年高性狀的遺傳增益分別為8.8%、19.7%和7.61%;硬闊林下,汪清種源表現最好,造林后地徑和當年高性狀上遺傳增益分別為22.92%和21.35%;雜木林下,鐵力種源表現最好,造林后在地徑、樹高、當年高和保存率性狀上可獲得的遺傳增益分別為10.19%、25.36%、6.99%和14.20%。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
