酵母發酵飼料對瘤胃菌群數量及多樣性的影響
2021-03-01 11:34:44郝凌魁劉世雄李冬芳于春微李松建胡紅蓮劉大程
中國畜牧雜志 2021年2期
郝凌魁,劉世雄,李冬芳,于春微,李松建,陳 灰,高 民,胡紅蓮,劉大程*
(1.內蒙古農業大學獸醫學院,內蒙古呼和浩特 010018;2.農業部動物疾病臨床診療技術重點實驗室,內蒙古呼和浩特 010018;3.內蒙古農牧業科學院動物營養與飼料研究所,內蒙古呼和浩特 010031)
近幾年生物飼料日益受到研究者關注。生物飼料不僅可以改善飼糧適口性、增強動物機體免疫功能、調節消化道微生態平衡、提高動物的生長性能,而且可以提高飼料的利用率、改善畜舍環境和降低飼料中的抗營養因子[1-4]。酵母發酵飼料為本課題組研制,由酵母菌為主的高活性菌株經液態富集培養、固態厭氧發酵及酵母破壁等發酵工藝生產而成,主要含有甘露聚糖、β-葡聚糖、多肽、有機酸和氨基酸5 種主效成分。前期試驗表明酵母發酵飼料不僅能提高肉羊的采食量和生產性能,而且可以增強肉羊免疫功能和抗應激能力[5-9]。反芻動物瘤胃的功能是通過瘤胃微生物來實現的[10-11],肉羊采食酵母發酵飼料后對瘤胃內主要功能菌的影響需要進一步闡明。基于此,本試驗旨在研究酵母發酵飼料對瘤胃菌群數量的影響,為其在生產中的應用提供理論依據。
1 材料與方法
1.1 酵母發酵飼料的制作 選取麩皮、玉米、棉籽粕、米糠、玉米胚芽粕、白玉米皮等原料作為發酵底物[5-6]。將活性釀酒酵母菌液(BC、XR4)以15%的接種量接入發酵底物中,加水使最終含水量為45%,然后進行固態堆積發酵。料高60 cm、發酵時間48 h、翻料控制發酵溫度在35℃左右。其主要技術指標:葡聚糖≥145 mg/100g,甘露聚糖≥71 mg/100 g,多肽≥1.3 mg/g,氨基酸≥15.27 mg/100g,有機酸≥1.48 mg/g。
1.2 試驗設計及日糧 選用10 只安裝永久性瘤胃瘺管(內徑4 cm)且體重約40 kg 的14 月齡蒙古羯羊,單籠飼養,自由飲水。將試驗動物隨機分為對照組和試驗組,每組5 只羊。對照組飼喂基礎日糧,試驗組飼喂基礎日糧+酵母發酵飼料,自由采食。課題組前期實踐證明,酵母發酵飼料的最適添加范圍是10%~15%[5],本試驗中添加量設計為日糧的13.37%。預試期15 d,正試期5 d,每天06:00 和18:00 飼喂。參考《肉羊飼養標準》(NY/T 816—2004)配制試驗飼糧。飼糧組成及營養成分見表1。
1.3 試驗方法
1.3.1 采集樣品 預試期結束后,正試期開始采樣,每天通過瘤胃瘺管在晨飼后0、3、6、9、12 h 采樣,每天采集一個時間點,共采5 次。用無菌鑷子夾取瘤胃內容物中的飼草料殘渣,并用負壓裝置抽取瘤胃中上層的瘤胃液和食糜,共計50 mL。放入經無菌水沖洗的自動勻漿機中攪拌勻漿15 s,后裝入無菌自封袋中,加入等體積磷酸緩沖液(pH 6.0,0.2 mol/L),冰浴拍打揉搓5 min,經2 層紗布過濾后4℃、1 000×g、15 min 離心取上清;所得上清液經4℃、10 000×g、25 min 再次離心后棄上清得沉淀加50 mL 緩沖液懸浮,經過液氮速凍后帶回實驗室-80℃保存。
1.3.2 實時熒光定量法的檢測
1.3.2.1 DNA 基因組提取及PCR 擴增 采用磁珠法土壤和糞便基因組DNA 提取試劑盒(TIANGEN,DP712)提取樣品基因組DNA,然后以其為模板進行PCR 擴增。PCR 反應體系:PCR Mix 10 μL,上、下游引物(10 μmol/L)各 0.5 μL,DNA 模板2 μL,ddH2O 12 μL;反應條件:95 ℃,3 min 變性;95 ℃,30 s;Tm,30 s;72℃,延伸1 min,40 個循環。查找GenBank 中目的基因的序列,利用Primer premier 等基因編輯軟件并結合參考文獻[8]設計目的基因的引物和探針序列,PCR 引物、探針序列和退火溫度見表2。PCR 擴增產物經瓊脂糖凝膠電泳檢測目的片段大小后,采用膠回收試劑盒(AXYGEN,AP-GX-50)回收PCR產物。

表1 飼糧組成及營養成分(干物質基礎)
1.3.2.2 qPCR 試驗 實時熒光定量PCR 采用Taqman探針法絕對定量。根據目的片段大小,選用pMD-19T為載體質粒進行目的片段的連接,隨后轉化進入DH5-α感受態細胞,經Amp 抗性LB 平板鑒定含有各目的片段的陽性克隆菌落后,接種于LB 液體培養基(含Amp)中,37 ℃振蕩過夜培養,用質粒小提試劑盒(TIANGEN,DP103)提取重組質粒。委托華大公司測序,在GenBank 上使用Blast 比對測序結果。利用分光光度計檢測比對結果相符的各質粒的核酸濃度,以10 倍倍比稀釋,作為標準品以制作標準曲線。
應用Applied Biosystems 7500 Fast Real-Time System 進行實時熒光信號值定量檢測。反應體系總體積為50 μL:Premix Ex Taq(Probe qPCR)(2×)25 μL,上、下游引物各(10 μmol/L) 1 μL,探針2 μL,ROX Reference Dye II(50×)0.5 μL,DNA 模板4 μL,ddH2O 16.5 μL。反應條件:95℃,30 s;95℃,5 s;Tm 34 s,40 個循環。
1.3.3 Illumina MiSeq 16S rRNA 高通量測序技術的檢測根據E.Z.N.A.?soil 試劑盒(Omega Bio-tek,Norcross,GA,U.S.)對各樣品進行DNA 抽提,用338F(5′-ACTC CTACGGGAGGCAGCAG-3 ′)和806R(5′-GGACTAC HVGGGTWTCTAAT-3′)引物對V3~V4 可變區進行PCR 擴增,擴增程序:95℃預變性3 min,27 個循環(95℃變性30 s,55℃退火30 s,72℃延伸30 s),最后72℃延伸10 min(PCR 儀:ABI GeneAmp?9700 型)。擴增體系為20 μL:4 μL Fast Pfu 緩沖液(5×),2 μL dNTPs(2.5 mmol/L),0.8 μL 引物(5 μmol/L),0.4 μL Fast Pfu 聚合酶;10 ng DNA 模板。
使用2%瓊脂糖凝膠回收PCR 產物,利用AxyPrep DNA Gel Extraction Kit(Axygen Biosciences,Union City,CA,USA)進行純化,Tris-HCl 洗脫,2% 瓊脂糖電泳檢測。利用QuantiFluorTM-ST (Promega,USA)進行檢測定量。根據Illumina MiSeq 平臺(Illumina,San Diego,USA)標準操作規程將純化后的擴增片段構建PE 2×300 的文庫。利用Illumina 公司的 Miseq PE300平臺進行測序(上海美吉生物醫藥科技有限公司)。
原始測序序列使用Trimmomatic 軟件質控,使用FLASH 軟件進行拼接,使用UPARSE 軟件(version 7.1 http://drive5.com/uparse/),根據 97%的相似度對序列進行 OTU 聚類;使用 UCHIME 軟件剔除嵌合體。利用 RDP classifier(http://rdp.cme.msu.edu/)對每條序列進行物種分類注釋,比對Silva 數據庫(SSU123),設置比對閾值為70%。
1.4 統計分析 實驗數據通過Excel 2010 整理后,使用SAS 9.0 統計軟件進行單因素方差分析。結果以平均值± 標準差表示。P<0.05 表示差異顯著,P<0.01 差異極顯著。
2 結果與分析
2.1 實時熒光定量PCR 結果
2.1.1 PCR 擴增結果 如圖1 所示,各陽性克隆菌落所提取質粒擴增的PCR 產物條帶單一,與各自對應的目的片段大小相符。進行序列同源性分析比對,相似性均大于99%,可滿足后續實驗要求。

表2 瘤胃細菌引物、探針及退火溫度
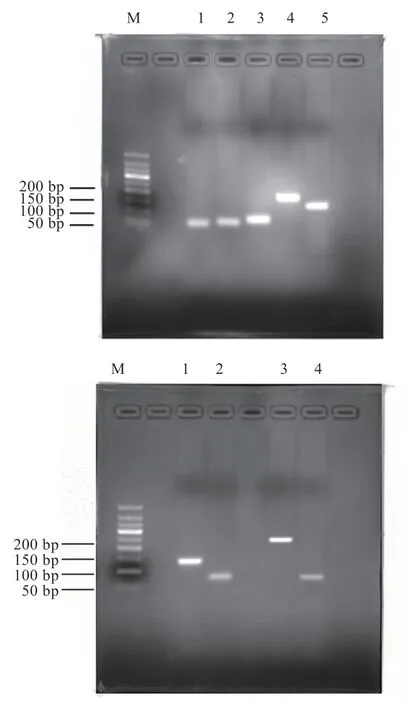
圖1 瓊脂糖凝膠電泳圖
2.1.2 熒光定量PCR 標準曲線方程 各菌的熒光定量PCR 標準曲線方程:Y溶纖維丁酸弧菌=-3.506logX+46.338;Y黃色瘤胃球菌=-3.637logX+46.800;Y產琥珀酸絲狀桿菌=-3.586logX+49.889;Y埃氏巨型球菌=-3.616logX+49.603;Y白色瘤胃球菌=-3.341logX+41.441;Y棲瘤胃普雷沃氏菌=-3.341X+41.044;Y嗜淀粉瘤胃球菌=-3.319X+41.417;Y反芻獸甲烷短桿菌=-3.625X+44.434;Y牛鏈球菌=-3.282X+45.000。式中,X表示拷貝數,Y表示Ct 值,各方程的R2值均大于等于0.999。
2.1.3 熒光定量PCR 測定結果 由表3 可知,試驗組的溶纖維丁酸弧菌在晨飼后3 h 數量最大(27.57×107copies/mL),且在晨飼后3、9、12 h 均高于對照組(P<0.05);但在晨飼后6 h 試驗組中溶纖維丁酸弧菌數量低于對照組(P<0.05)。試驗組中黃色瘤胃球菌在晨飼后9 h 數量最大(4.32×107copies/mL),且在晨飼后6、9、12 h均高于對照組(P<0.05)。試驗組產琥珀酸絲狀桿菌在晨飼后6 h 數量最大(4.27×107copies/mL),3、6 h 均高于對照組(P<0.05)。試驗組中埃氏巨型球菌的數量在晨飼后6 h(P<0.01)、在晨飼后9 h(P<0.05)高于對照組。試驗組白色瘤胃球菌的數量最高值在晨飼后9 h(22.30×104copies/mL),高于對照組中的最高值(18.67×104copies/mL)(P>0.05)。試驗組中牛鏈球菌的數量在晨飼后3 h 高于對照組(P<0.01)。在試驗組中嗜淀粉瘤胃桿菌的數量與對照組差異不顯著。試驗組棲瘤胃普雷沃氏菌的數量在晨飼后3 h 和6 h 均高于對照組(P<0.01)。試驗組中反芻獸甲烷短桿菌的數量在晨飼后0 h 高于對照組,但在6 h 低于對照組中(P<0.01),且3、9、12 h 均小于對照組的數量(P>0.05)。
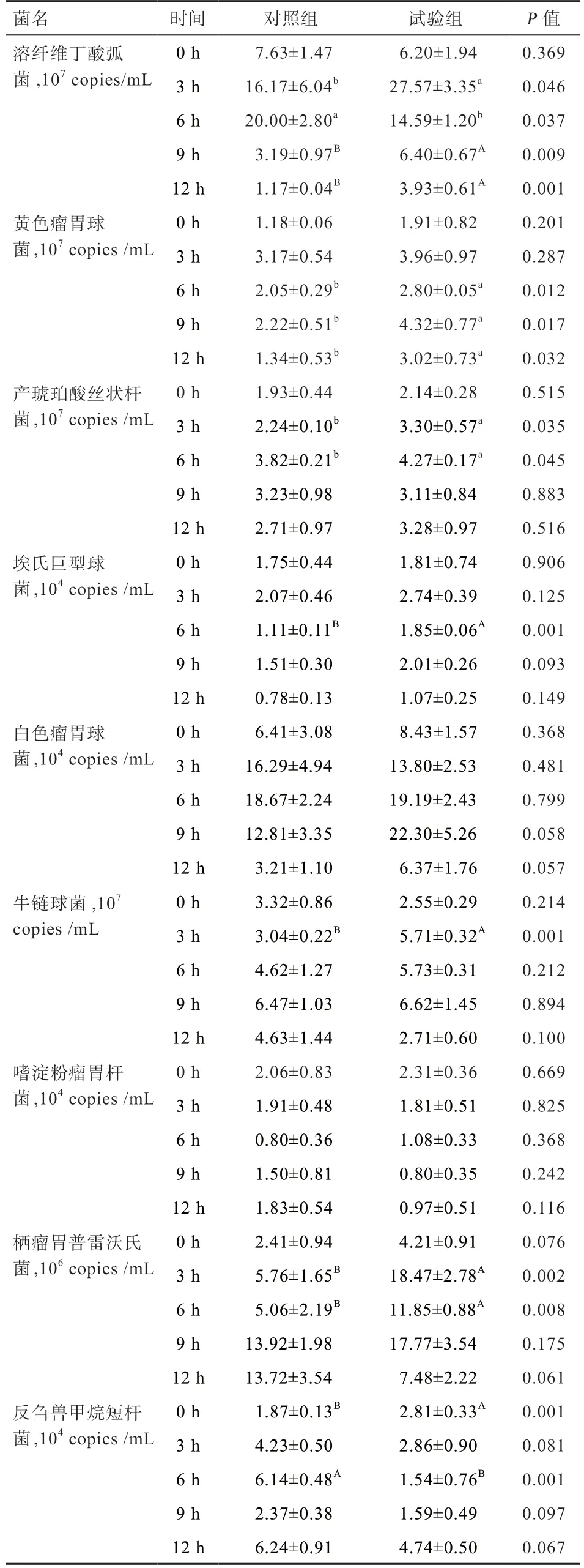
表3 熒光定量PCR 結果
2.2 Illumina MiSeq 16s rRNA 高通量測序結果
2.2.1 樣本DNA 和測序數據評估 瓊脂糖凝膠電泳檢測DNA 質量結果顯示,提取的DNA 純度良好,PCR產物目的條帶大小正確,濃度合適。通過16S rRNA基因測序質控篩選后進行測序深度檢測,所有樣本的Shannon 稀釋曲線顯示曲線趨向平坦(圖2),說明本次測序數據量足夠大,已經基本覆蓋到樣品中所有的物種,可以反映樣本中絕大多數的微生物多樣性信息。
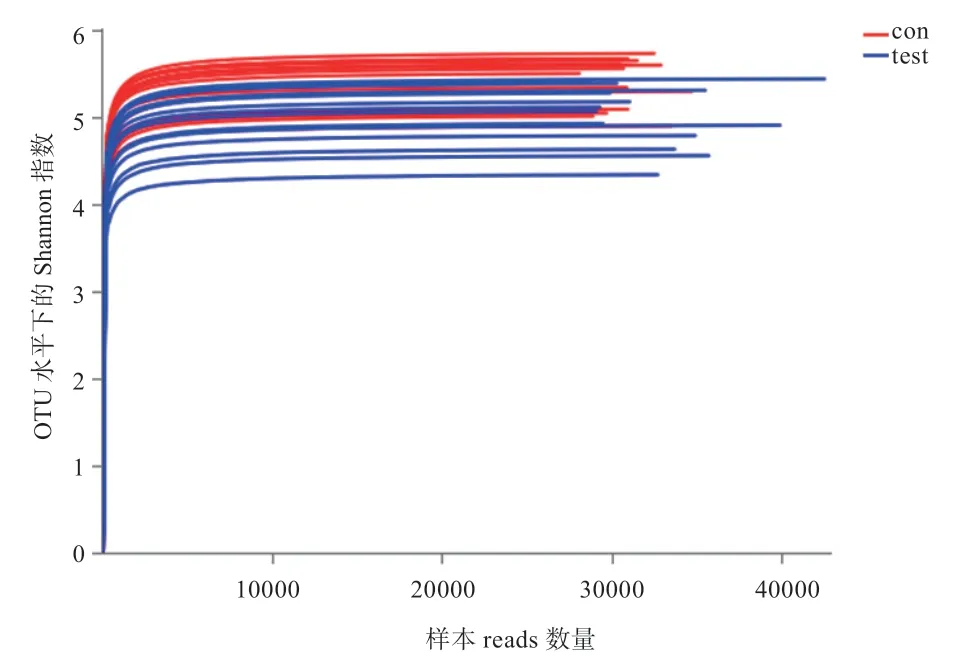
圖2 Shannon 稀釋曲線圖(相似閾值為 97%)
2.2.2 Alpha 多樣性分析 由表4 可知,試驗組的Shannon指數低于對照組(P<0.01);試驗組的Simpson 指數高于對照組(P<0.05);Sobs 指數、Ace 指數和Chao 指數在兩組中沒有顯著差異。說明試驗組的多樣性顯著低于對照組,而豐富度沒有差異。
2.2.3 Venn 圖分析 如圖3 所示,在相似度為97% 的水平下,2 組共聚類得到2 306 個OUT,對照組和試驗組分別為2 179 個和2 116 個,共用1 989 個OUT,占OUT 總數的86.25%,獨有的OUT 數分別為190 個和127 個,分別占OUT 總數的8.23% 和5.51%,表明2組 OUT 組成具有一定的相似度,但也存在差異。

表4 Alpha 多樣性分析結果
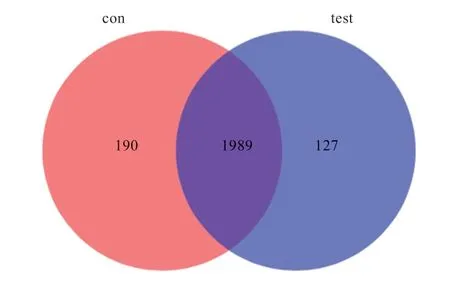
圖3 OUT 水平下瘤胃微生物Venn 圖
2.2.4 群落組成分析 經過物種分類學比較,樣品共包含20 門,34 綱,63 目,93 科,235 屬。
2.2.4.1 門水平 如圖4 所示,在門水平分類上,占主導的為擬桿菌門(Bacteroidetes)、厚壁菌門(Firmicutes)、螺旋體門(Spirochaetae)和纖維桿菌門(Fibrobacteres)。此外,軟壁菌門(Tenericutes)、疣微菌門(Verrucomicrobia)和變形菌門(Proteobacteria)占比均高于1%。擬桿菌門在試驗組中最大值為64.05%,高于在對照組中的最大值57.48%(P>0.05)。厚壁菌門在試驗組中最大值為32.29%,低于對照組中的最大值34.06%(P>0.05)。纖維桿菌門與螺旋體門在對照組與試驗組中的相對豐度最高值均在0 h。纖維桿菌門在試驗組中的平均值(4.02%)高于對照組(3.40%);螺旋體門在試驗組中的平均值(5.08%)高于對照組(4.41%)(P>0.05)。其余相對豐度低于0.1%的門類被聚集到一起(others<0.1%)。
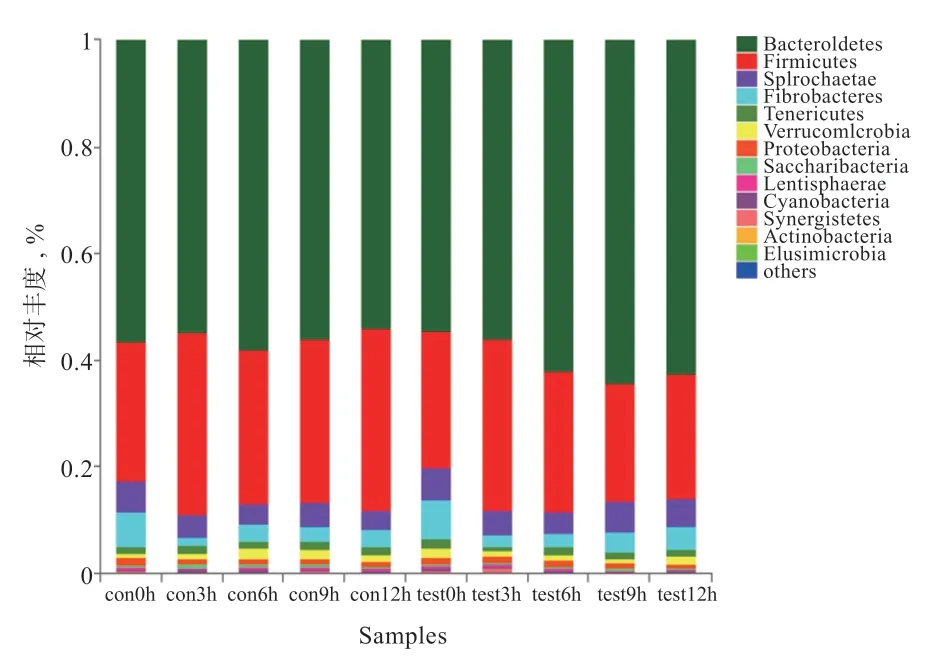
圖4 瘤胃菌群門水平相對豐度
2.2.4.2 屬水平 如圖5 所示,優勢菌屬為普雷沃氏菌屬_1(Prevotella_1)、理研菌科_RC9 菌屬(Rikenellaceae_RC9_gut_group)、未排位擬桿菌_BS11 菌屬(norank_f_Bacteroidales_BS11_gut_group)、密螺旋體菌屬_2(Treponema_2)和纖維桿菌屬(Fibrobacter)等,其余相對豐度低于1%的門類被聚集到一起(others<1%)。普雷沃氏菌屬_1 在試驗組中平均值(34.34%)高于對照組(33.52%)(P>0.05)。理研菌科_RC9 菌屬在試驗組中平均值(12.03%)高于對照組(6.91%)(P<0.05)。纖維桿菌屬在對照組的平均值(4.14%)高于試驗組(3.39%)(P>0.05)。瘤胃球菌科_NK4A214 菌屬(Ruminococcaceae_NK4A214_group)在對照組的平均值(3.14%)高于試驗組(3.06%)(P>0.05)。丁酸弧菌屬_2(Butyrivibrio_2)在試驗組的平均值(1.72%)高于對照組(1.20%)(P>0.05)。丹毒進菌科_UCG-004 菌屬(Erysipelotrichaceae_UCG-004)在試驗組的平均值(1.47%)高于對照組(0.77%)(P<0.05)。韋榮球菌科_UCG-001 菌屬(Veillonellaceae_UCG-001)在試驗組的平均值(0.65%)低于對照組(1.31%)(P<0.05)。克里斯滕森菌屬_R-7(Christe nsenellaceae_R-7_group)在試驗組中的平均值(0.79%)低于對照組(1.12%)(P<0.05)。
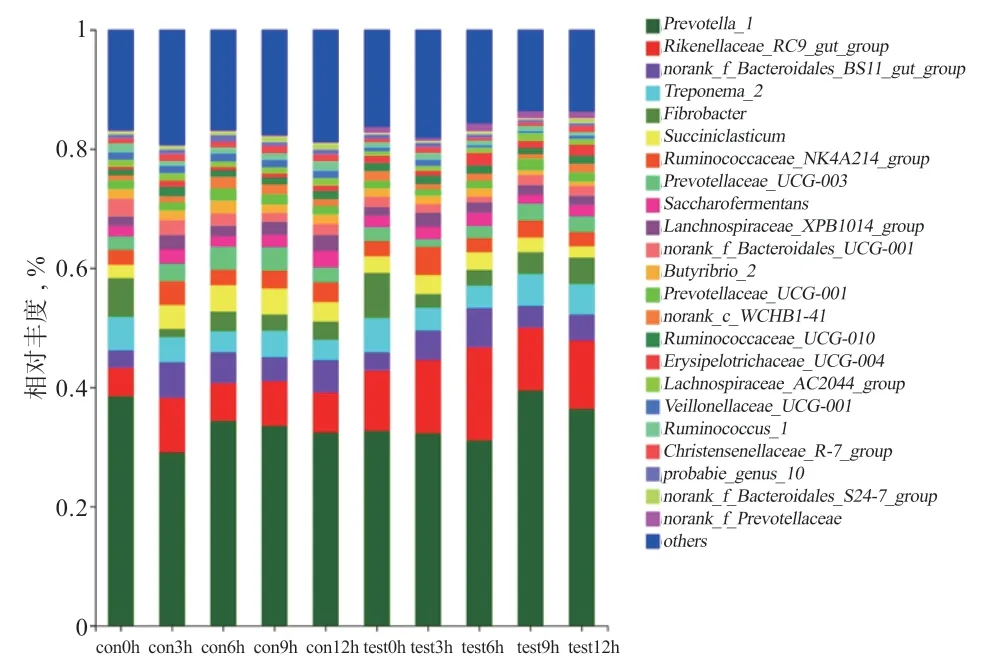
圖5 瘤胃菌群屬水平相對豐度
2.2.5 PLS-DA 分析 PLS-DA 分析作為樣本分組分析中的一種,具有忽略組內的隨機差異,突出組間系統差異的特點。各點之間的距離代表各個樣本組成的相似性與差異性,其距離越近,樣本的相似性越高;距離越遠,樣本的相似性越低。如圖6 所示,2 個組內各點的距離接近,具有相似性,而兩組間的距離偏遠,具有一定的差異性。
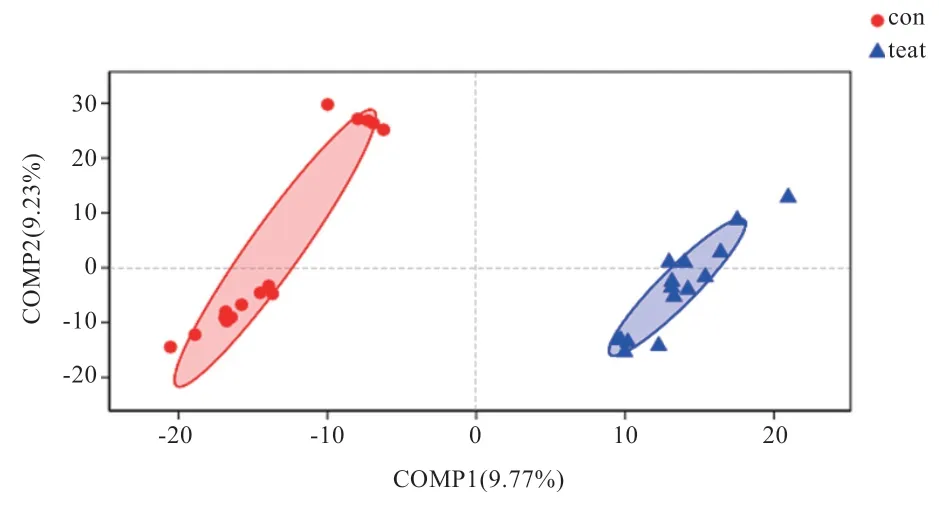
圖6 PLS-DA 分析結果圖
3 討 論
3.1 瘤胃液采樣方法的分析 反芻動物瘤胃中微生物種群根據寄生的位置不同可分為固相瘤胃微生物、液相瘤胃微生物和貼附于瘤胃壁的微生物三類。在瘤胃液采集工作中,如單一抽取瘤胃液相微生物作為樣本進行試驗分析,固態內容物上黏附的大量細菌就可能沒有被取樣,樣品出現偏差,整體數據的準確性會受到影響,因此,要根據微生物寄生的不同位置,采集不同瘤胃內容物混勻后才可作為樣品[12-13]。本試驗采用對瘤胃內容物殘渣、食糜和瘤胃液結合,并用緩沖液清洗的方法,獲得較高代表性的瘤胃菌群樣品。
3.2 9 種瘤胃菌數量變化分析 反芻動物的瘤胃器官是一個高效的發酵系統,其獨特優勢在于可以降解植物源類纖維素。大量的微生物寄生于瘤胃中,參與消化和吸收纖維素類營養物質的過程。其中,黃色瘤胃球菌、白色瘤胃球菌和產琥珀酸絲狀桿菌占據著主導地位。經過植物乳酸菌、枯草芽孢桿菌、地衣芽孢桿菌、或酵母菌等微生物發酵處理的飼料飼喂反芻動物后,該3 種菌的數量均有極顯著提高[14-16]。Hoang 等[17]試驗表明,由枯草芽孢桿菌、布拉迪酵母菌和乳酸菌組成的復合菌活性添加劑可提高機體對粗纖維等營養物質的消化率,進而提高飼料轉化效率。本試驗中,酵母發酵飼料增加了黃色瘤胃球菌和產琥珀酸絲狀桿菌的數量,可提高機體對纖維素類營養物質的消化率。除了纖維素類分解菌之外,本研究中溶纖維丁酸弧菌、棲瘤胃普雷沃氏菌和埃氏巨球型菌的數量也在試驗組中有所增加。溶纖維丁酸弧菌和棲瘤胃普雷沃氏菌具有淀粉降解酶活性和蛋白降解酶活性,可以降解蛋白質和淀粉等多糖類大分子的營養物質[18]。溶纖維丁酸弧菌還可參與體內的氫化作用,將不飽和脂肪酸轉變成飽和脂肪酸,提高機體的增重效果[19]。酵母發酵飼料中富含甘露聚糖、葡聚糖和氨基酸,可誘導溶纖維丁酸弧菌和棲瘤胃普雷沃氏菌的大量增殖。在現代肉羊養殖業中,大比例使用精飼料以達到短時間增重的目的,使得瘤胃中牛鏈球菌快速增殖生長并產生乳酸,易造成肉羊瘤胃酸中毒的現象。以氨基酸為氮源的埃氏巨球型菌可代謝乳酸,該菌在防止乳酸性酸中毒、提高淀粉的利用率、參與氨基酸的脫氨脫羧反應以及形成揮發性支鏈脂肪酸的過程中都發揮重要的作用[20]。酵母發酵飼料提高了埃氏巨球型菌數量,可提高營養物質消化率,也可預防瘤胃酸中毒,維持瘤胃微生態環境的穩定。此外,酵母細胞壁的特殊空間結構可以吸附黃曲霉毒素、玉米赤霉烯酮等霉菌毒素。酵母細胞壁中的多糖類物質能競爭性地阻止病原菌在腸壁上的定植,可有效減少金黃色葡萄球菌、沙門氏菌、大腸桿菌等病原菌數量,有利于瘤胃細菌的繁殖并改善瘤胃微生態環境[21-25]。微生物發酵飼料對牛鏈球菌影響的報道并不一致[8,26],造成差異的原因可能與發酵底物、發酵菌種以及試驗動物的品種有關。本文中酵母發酵飼料增加了牛鏈球菌數量。反芻獸甲烷短桿菌是瘤胃中最主要的產甲烷菌。蘋果酸或延胡索酸等有機酸物質可通過氫代謝途徑產生丙酸和共軛亞油酸,消耗氫離子來抑制甲烷的生成[27]。有研究表明飼糧中添加乳酸菌或活性干酵母可降低產甲烷短桿菌屬的數量,降低了甲烷排放量約40%[28-29]。本研究中的酵母發酵飼料也可顯著降低反芻獸甲烷短桿菌數量,減輕畜牧生產中甲烷對外界環境的污染。
3.3 16S rRNA 基因高通量測序結果的分析 諸多學者利用高通量測序技術發現,生物飼料可提高瘤胃菌群的多樣性與豐度[30]。本研究結果與之相悖,可能與飼料的高消化率與高效的代謝途徑相關。部分菌群因為具有高效簡便的代謝通路,可在營養物質消化過程中占據主導地位,在短時間內積累大量的代謝產物。高效的微生物菌群并不會出現冗雜的結構層次,能快速地滿足機體能量需求即可[31]。Bagheri[32]認為玄參提取物和莫能菌素可使菌群多樣性降低。本研究中酵母發酵飼料降低了瘤胃微生物菌群的多樣性,使瘤胃細菌更有效率地降解營養物質,這可能是其能夠顯著提高綿羊生長性能和免疫功能的原因[5]。在門水平上,擬桿菌門和厚壁菌門是反芻動物瘤胃中的優勢菌門[33]。其次,根據物種、日糧、年齡和生長環境等條件的不同,纖維桿菌門和密螺旋體門會出現不同的優勢占比。擬桿菌門在非纖維物質的降解中發揮重要作用,而厚壁菌門、螺旋菌門和纖維桿菌門能夠降解纖維素、半纖維素、果膠等,對植物類纖維物質轉化為揮發性脂肪酸(VFA)產生重要影響[34]。有研究表明隨著酵母培養物(Yeast Culture,YC)添加水平的升高,擬桿菌門相對豐度逐漸增加,厚壁菌門的相對豐度逐漸降低,螺旋菌門和纖維桿菌門的數量也會出現相應的變化,但不顯著[34]。本試驗結果也出現了類似的變化趨勢。在屬水平上,本試驗中的普雷沃氏菌屬_1 的相對豐度最高,與前人研究結果[35]一致。普雷沃氏菌屬_1 在各組間的差異不顯著,可能因為對照組與試驗組的飼糧粗蛋白質和能量水平相近,沒有對普雷沃氏菌屬的相對豐度產生顯著影響。研究發現理研菌科_RC9 菌屬含量隨著日糧纖維含量的降低,表達量降低[36]。理研菌科具有降解結構性碳水化合物的作用[37],酵母發酵飼料可增加理研菌科含量以提高纖維素的降解。克里斯滕森菌屬于厚壁菌門,在維持胃腸道結構、功能和動物機體免疫調節方面發揮重要作用[38]。有研究認為克里斯滕森菌的相對豐度隨著YC 添加水平的升高而降低,且與瘤胃pH 和NH3-N 含量呈顯著正相關,說明克里斯滕森菌屬與VFA 和NH3-N 的代謝相關[34]。本課題組前期試驗表明,酵母發酵飼料可增加瘤胃中VFA,降低NH3-N 的濃度[9,39]。所以克里斯滕森菌屬_R-7 含量增加受到了酵母發酵飼料的正向影響,進而改善了瘤胃的微環境。
16S rRNA 高通量測序結果最低到屬水平,需要結合實時熒光定量PCR 的結果細化到種水平。在qPCR檢測中,隸屬于擬桿菌門的溶纖維丁酸弧菌、黃色瘤胃球菌、棲瘤胃普雷沃氏菌、和隸屬于纖維桿菌門的產琥珀酸絲狀桿菌的變化趨勢與高通量測序結果保持一致。高通量測序結果顯示厚壁菌門在試驗組中含量較低,而qPCR 檢測中埃氏巨型球菌在對照組中含量較低,二者結果不一致。在屬水平上,高通量測序結果中纖維桿菌屬和瘤胃球菌屬在對照組中的含量略高于試驗組,與qPCR 檢測中產琥珀酸絲狀桿菌和黃色瘤胃球菌的結果不一致,可能原因為qPCR 引物的特異性較高,而高通量測序中使用的是通用引物。此外,反芻獸甲烷短桿菌屬于古細菌中的廣古菌門,不屬于細菌。高通量測序中的細菌V3~V4 區通用引物未能涵蓋檢測,需要qPCR檢測來補充說明。
近年來利用分子生物學方法研究反芻動物瘤胃調控和微生物菌群之間互作關系雖有些許成果,但還需全面地深入了解和揭示。有國外的學者應用計算硬件及配套的控制理論,設計了高性能微生物群落數據分析方法GPU-Meta-Storms,可實現在微秒的時間內對微生物群落的環境樣本元基因組數據進行比較,未來可能應用于監控瘤胃微生物的實時動態變化。另外,隨著完整基因組序列數據庫不斷擴大,再加上環境特征數據的補充完善,可進一步評估酵母發酵飼料對瘤胃微生物多樣性及其功能的影響[40]。
4 結 論
本研究結果表明,酵母發酵飼料可促進纖維降解菌、蛋白降解菌、淀粉降解菌、乳酸利用菌的生長繁殖,降低產甲烷菌的數量和瘤胃細菌的多樣性,改善瘤胃菌群結構。
