可變剪接的表觀遺傳學(xué)調(diào)控機(jī)制及其在脂肪代謝中的作用研究進(jìn)展
2021-03-01 11:34:34馬海明張躍博
中國畜牧雜志 2021年2期
騫 鑫,馬海明,何 俊,徐 康,張躍博*
(1.湖南農(nóng)業(yè)大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院,湖南長沙 410128;2.中國科學(xué)院亞熱帶農(nóng)業(yè)生態(tài)研究所,湖南長沙 410125)
mRNA 的轉(zhuǎn)錄后加工是基因表達(dá)必需一個(gè)基本的生物學(xué)過程,在高等真核生物中蛋白質(zhì)多樣性很大程度上是由Pre-mRNA 可變剪接引起的,約90% 的人類基因經(jīng)歷此過程,基因通過可變剪接等表達(dá)調(diào)控機(jī)制控制著細(xì)胞的增殖、分化、凋亡等生物學(xué)進(jìn)程,剪接的異常會(huì)引起蛋白質(zhì)的功能異常甚至導(dǎo)致疾病的發(fā)生[1-2],所涉及的詳細(xì)機(jī)制則需要進(jìn)一步的研究。表觀遺傳修飾是在不改變DNA 序列的基礎(chǔ)上參與基因組的調(diào)控,即可以直接作用在DNA 或RNA 上,也可以作用在與DNA結(jié)合的蛋白上,對(duì)表觀遺傳學(xué)修飾的研究將大大提高對(duì)基因表達(dá)調(diào)控的理解[3-4],越來越多的研究證明DNA 甲基化、RNA 編輯以及非編碼RNA 等表觀遺傳學(xué)修飾在可變剪接的啟動(dòng)以及剪接位點(diǎn)的識(shí)別中起到重要作用,這提示著表觀遺傳學(xué)在pre-mRNA剪接中的重要意義[5]。脂質(zhì)是細(xì)胞內(nèi)重要的組成成分和信號(hào)分子,參與細(xì)胞各生命活動(dòng),機(jī)體在生理或病理?xiàng)l件下能改變細(xì)胞內(nèi)脂質(zhì)組成,針對(duì)脂質(zhì)代謝的研究對(duì)治療疾病或者改良動(dòng)物經(jīng)濟(jì)性狀都具有重要意義。近年來已經(jīng)有大量證據(jù)證明可變剪接在脂質(zhì)代謝過程發(fā)揮著重要作用。本文主要介紹了表觀遺傳學(xué)對(duì)于可變剪接的調(diào)控機(jī)制以及可變剪接在脂肪代謝中相關(guān)功能的研究進(jìn)展。
1 可變剪接的表觀遺傳學(xué)調(diào)控
在Pre-mRNA 剪接過程中,剪接體在剪接因子的輔助下識(shí)別并結(jié)合到mRNA 前體位于外顯子/內(nèi)含子邊界的特定序列,將內(nèi)含子切除和外顯子連接產(chǎn)生成熟的mRNA[6]。在此過程中,表觀遺傳學(xué)修飾起著重要作用,這涉及到DNA 甲基化(DNA Methylation)、RNA 編輯、組蛋白修飾等多種表觀遺傳學(xué)進(jìn)程,以下主要介紹表觀遺傳學(xué)修飾對(duì)于可變剪接的調(diào)控機(jī)制。
1.1 DNA 甲基化與可變剪接 DNA 甲基化是在DNA甲基化酶(DNA methyltransferases,Dnmt)的作用下,以S-腺苷甲硫氨酸作為甲基供體,使DNA 序列上特定的堿基通過共價(jià)鍵結(jié)合的方式獲得1 個(gè)甲基團(tuán)的化學(xué)修飾過程[7]。DNA 甲基化不僅可通過調(diào)控RNA 聚合酶II(pol II)在Pre-mRNA 上的移動(dòng)速率來影響剪接信號(hào)的識(shí)別,還能通過影響組蛋白修飾調(diào)節(jié)剪接調(diào)節(jié)因子的招募,從而實(shí)現(xiàn)對(duì)Pre-mRNA 剪接的調(diào)節(jié)。
CTCF 是一種可以與DNA 序列產(chǎn)生特異性結(jié)合的DNA 結(jié)合蛋白,其與DNA 序列的特異性結(jié)合可調(diào)節(jié)pol II 在pre-mRNA 序列上的移動(dòng)速率。DNA 甲基化可以抑制CTCF 與DNA 序列上外顯子的特異性結(jié)合,影響剪接過程中外顯子的剪接[8]。MeCP2 可以通過與甲基化DNA 的特異性結(jié)合抑制其下游靶基因的轉(zhuǎn)錄。當(dāng)DNA 發(fā)生甲基化時(shí)會(huì)促使MeCP2 結(jié)合在DNA 序列上招募組蛋白去乙酰酶,使剪接位點(diǎn)上游組蛋白發(fā)生去乙酰化,導(dǎo)致pol II 在Pre-mRNA 序列上延伸速率減緩,緩慢延伸的pol II 較正常生理情況下的pol II 更能夠識(shí)別序列上弱的剪接信號(hào),使得剪接時(shí)外顯子得以保留[9]。
在Pre-mRNA 的剪接過程中,HP1 可以協(xié)助招募U2 snRNP 到H3K9 甲基化組蛋白上,進(jìn)而影響靶序列轉(zhuǎn)錄調(diào)節(jié)因子的招募[10]。為證明DNA 甲基化對(duì)于剪接的調(diào)控與HP1 相關(guān),Yearim 等[10]對(duì)受甲基化影響的外顯子剪接區(qū)域和受HP1 介導(dǎo)的組蛋白修飾影響的外顯子剪接區(qū)域進(jìn)行定位比對(duì),結(jié)果顯示受甲基化影響的靶基因剪接區(qū)域與受HP1 介導(dǎo)的組蛋白修飾影響的外顯子剪接區(qū)域基本相同,證明了DNA 甲基化對(duì)靶基因的剪接調(diào)控與HP1 之間存在著一定關(guān)系。
1.2 N6-甲基腺嘌呤與可變剪接 N6-甲基腺嘌呤(N6-methyladenosine,m6A)修飾是高等生物mRNA 上最為普遍的修飾,約占所有RNA 修飾的60%以上[11]。m6A過程是由METTL13、METTL14、WTAP、RBM15、KIAA-1429 等甲基化酶催化完成的,這些酶在哺乳動(dòng)物中被稱為“Writers”,m6A 甲基化過程是可逆的,整個(gè)過程由FTO、ALKBH5 等去甲基化酶催化,這些酶被統(tǒng)稱為“Erasers”,而m6A 對(duì)可變剪接產(chǎn)生調(diào)控作用是通過YTHDF1、eIF3 等名為“Readers”的甲基化閱讀蛋白來實(shí)現(xiàn)。
METTL13、METTL14 和WTAP 是具有代表性的m6A 甲基化酶,其功能是催化Pre-mRNA 上腺苷酸發(fā)生m6A 修飾,三者通過形成WMM 復(fù)合物行使對(duì)基因剪接的調(diào)控功能。Ping 等[12]對(duì)缺乏WMM 復(fù)合物的細(xì)胞中的基因進(jìn)行功能富集分析,發(fā)現(xiàn)富集在RNA 加工功能上的差異表達(dá)基因顯著減少,說明m6A 甲基化酶通過可變剪接在RNA 的多樣性上起到至關(guān)重要的作用。
FTO 作為標(biāo)志性的去甲基化酶,其表達(dá)量和m6A甲基化水平呈負(fù)相關(guān)。Zhao 等[13]研究發(fā)現(xiàn),在3T3-L1 細(xì)胞中m6A 修飾富集于5' 或者3' 剪接位點(diǎn)附近的外顯子區(qū)域內(nèi),與參與mRNA 剪接調(diào)控的一種RNA 結(jié)合蛋白結(jié)合區(qū)域重疊,當(dāng)FTO 表達(dá)量過低時(shí),m6A 水平升高可以促進(jìn)剪接因子SRSF2 的RNA 結(jié)合能力,最終導(dǎo)致剪接過程中外顯子保留。在脂肪的生成過程中,具有催化活性的FTO 對(duì)于脂肪細(xì)胞的分化以及脂肪的生成具有促進(jìn)作用,這揭示了依賴FTO 的m6A 去甲基化作為RNA 加工的一種新的調(diào)控機(jī)制,在脂肪形成的調(diào)控中發(fā)揮著關(guān)鍵作用。
YTHDF1 作為一種閱讀蛋白,其主要功能是識(shí)別發(fā)生m6A 修飾的堿基,從而激活下游的調(diào)控通路。YHTDF1 可以識(shí)別編碼序列和3'-UTR 區(qū)上的m6A 修飾并招募剪接調(diào)節(jié)因子SRSF 與之結(jié)合,進(jìn)而對(duì)下游的剪接進(jìn)行調(diào)控,其中SRSF3 和SRSF10 與YHTDF1 的結(jié)合區(qū)域基本相同,說明這2 種剪接調(diào)節(jié)因子與YHTDF1 的結(jié)合具有競爭性,但SRSF3 比SRSF10 具有更高的優(yōu)先級(jí)。在正常生理?xiàng)l件下YHTDF1 識(shí)別m6A 并且優(yōu)先招募SRSF3 并與之結(jié)合,促進(jìn)外顯子保留[14]。
1.3 RNA 編輯與可變剪接 RNA 編輯即在初級(jí)轉(zhuǎn)錄本中發(fā)生堿基的加入、丟失或轉(zhuǎn)換現(xiàn)象。在某些情況下,通過RNA 編輯在非剪接位點(diǎn)附近形成典型的剪接受體末端會(huì)導(dǎo)致剪接提前發(fā)生。腺苷脫氨酶2(Adenosine Deaminase RNA Specific 2,ADAR2)的Pre-mRNA 中腺苷到肌苷的核苷酸修飾已被證明影響下游的可變剪接事件[16],Shenasa 等[17]通過將大鼠ADAR2基因AA 編輯為AI,由此產(chǎn)生的AI 二核苷酸被識(shí)別為3'剪接位點(diǎn),導(dǎo)致剪接激活,新的3'剪接位點(diǎn)的出現(xiàn)增加了47 個(gè)核苷酸到ADAR2mRNA,最終導(dǎo)致閱讀框架改變,而該基因發(fā)生RNA 編輯所產(chǎn)生的異常轉(zhuǎn)錄本亦能反向調(diào)控該基因RNA 前體的RNA 編輯,揭示了RNA 編輯是調(diào)節(jié)ADAR2不同轉(zhuǎn)錄本表達(dá)水平的一種調(diào)節(jié)機(jī)制[18]。此外,如富含絲氨酸/精氨酸(Serine/Arginine-Richprotien,SR)蛋白和核不均一核糖蛋白(Heterogeneous Nuclear Ribonucleoprotein,hnRNPs)等反式作用因子可以通過與順式作用元件序列上的剪接調(diào)控元件(Splicing Regulatory Elements,SREs)結(jié)合來調(diào)節(jié)內(nèi)含子保留水平。Solomon 等[19]研究發(fā)現(xiàn),在ADAR編輯位點(diǎn)上A到I 的編輯可以制造或破壞順式作用元件上的SREs,從而影響外顯子上剪接因子的結(jié)合,這是RNA 編輯對(duì)于可變剪接的另一種調(diào)控機(jī)制。
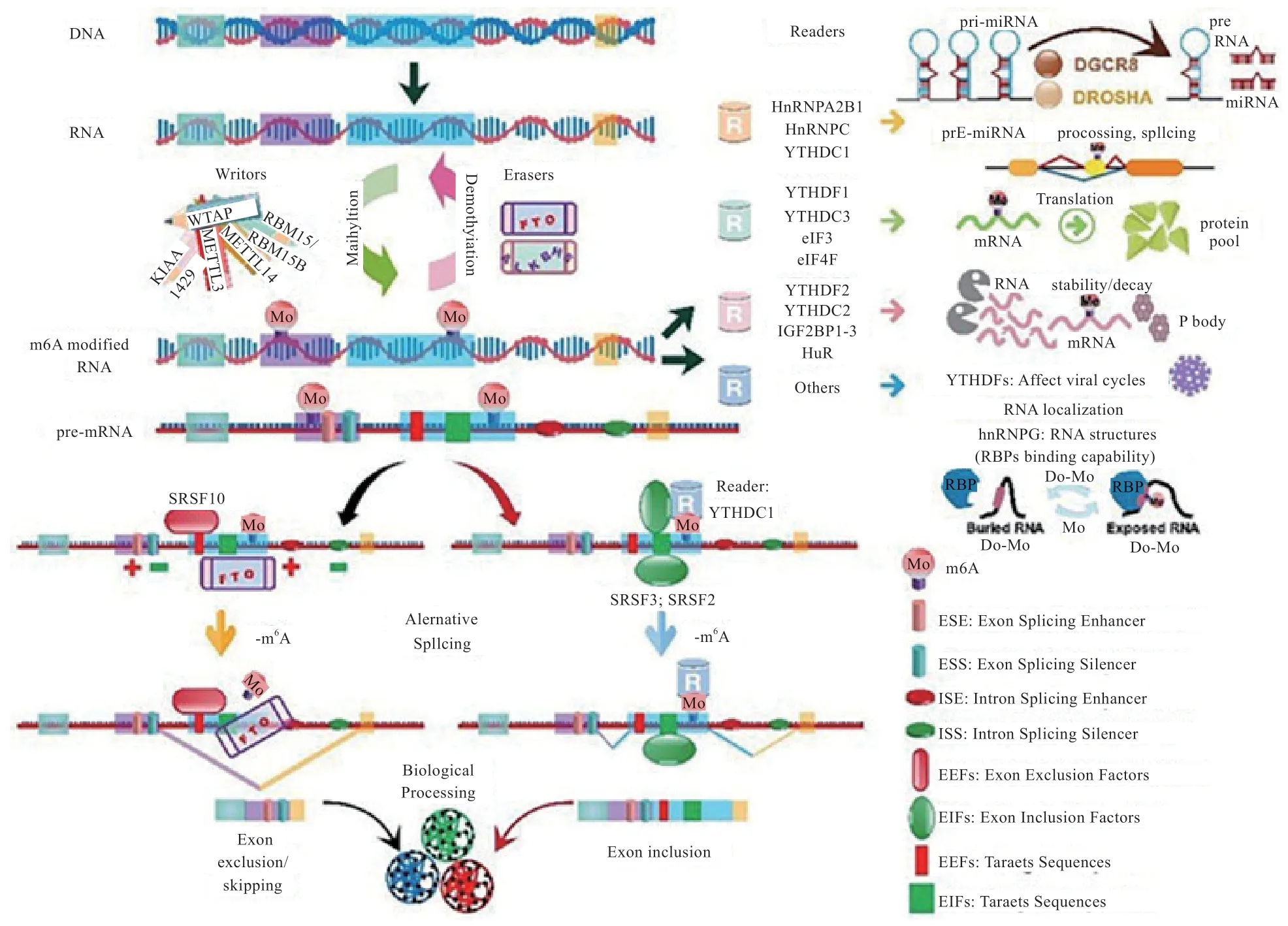
圖1 m6A 對(duì)可變剪接的調(diào)控機(jī)制[15]
1.4 組蛋白修飾與可變剪接 組蛋白是存在于真核生物染色質(zhì)中與DNA 結(jié)合的蛋白質(zhì),組蛋白的N 末端尾部和球狀結(jié)構(gòu)域含有許多可被修飾的氨基酸,可通過發(fā)生一系列共價(jià)修飾改變?nèi)旧|(zhì)構(gòu)型或者招募轉(zhuǎn)錄調(diào)節(jié)因子調(diào)控Pre-mRNA 剪接[20]。組蛋白去乙酰化酶抑制劑TSA 對(duì)細(xì)胞的處理可誘導(dǎo)神經(jīng)細(xì)胞黏附分子(Neural Cell Adhesion Molecule,NCAM)外顯子18 的外顯子跳躍[21],生理?xiàng)l件下人神經(jīng)細(xì)胞的去極化增加了NCAM外顯子18 周圍的H3K9 乙酰化和H3K36 甲基化,致使轉(zhuǎn)錄速率加快,誘導(dǎo)發(fā)生外顯子跳躍事件[22],與利用組蛋白去乙酰化酶抑制劑TSA 對(duì)細(xì)胞處理后的NCAM具有相同效果,提示著細(xì)胞去極化可能通過局部組蛋白的乙酰化和甲基化調(diào)控外顯子的剪接。成纖維細(xì)胞生長因子受體2(Fibroblast Growth Factor Receptor 2,F(xiàn)GFR2)通過可變剪接產(chǎn)生2 種具有高度的組織特異性的剪接變體FGFR2-IIIb 和FGFR2-IIIc。在間充質(zhì)細(xì)胞中,F(xiàn)GFR剪接位點(diǎn)附近富集的H3K36me3 與H3K4me1 修飾通過對(duì)招募剪接因子的影響Pre-mRNA的剪接最終產(chǎn)生FGFR2-IIIc,上皮細(xì)胞中FGFR剪接位點(diǎn)附近主要發(fā)生H3K27me3 和H3K4me3 修飾,其剪接形式則主要是外顯子IIIb 保留[23-24]。Luco 等[23]通過過表達(dá)或下調(diào)組蛋白甲基轉(zhuǎn)移酶調(diào)控H3K36me3 或H3K4me3 水平可以操控FGFR2的剪接方式,揭示了通過對(duì)組蛋白修飾調(diào)控可影響外顯子的剪接。腺病毒E1A相關(guān)蛋白BS69 是一種具有多個(gè)功能結(jié)構(gòu)域的細(xì)胞蛋白,包括PHD、Bromo、PWWP 以及MYND 結(jié)構(gòu)域[25],該蛋白上的PWWP 結(jié)構(gòu)域可以特異性識(shí)別H3K36me3以及H3.3K36me3 并與之結(jié)合,作為剪接因子調(diào)控pol II的延伸速率最終影響可變剪接[26]。此外,BS69 還可以通過識(shí)別H3.3K36me3 并招募調(diào)控U5 snRNP 生成的剪接因子EFTUD2 調(diào)控Pre-mRNA 的剪接[27]。
1.5 非編碼RNA 與可變剪接 非編碼RNA(Non-coding RNA,ncRNA)是從基因組上轉(zhuǎn)錄而來不具備翻譯功能的RNA[28]。按照其生物學(xué)功能,可以將ncRNA 分為持家ncRNA 以及調(diào)控ncRNA,其中調(diào)控ncRNA 又包括短調(diào)控ncRNA(如siRNA、miRNA)、長調(diào)控ncRNA(如lncRN)等[29],ncRNA 不僅可以從RNA水平上影響基因表達(dá),還可以通過影響組蛋白修飾等間接影響剪接行使對(duì)基因的表達(dá)調(diào)控功能。
1.5.1 sncRNA 與可變剪接 miRNA 是一類小于22nt 的sncRNA,參與調(diào)節(jié)所有細(xì)胞信號(hào)通路,其表達(dá)水平的改變與組織的生長發(fā)育和疾病有關(guān)[30-31],miRNA 主要位于編碼基因的內(nèi)含子中,通過堿基配對(duì)與細(xì)胞質(zhì)中靶mRNA轉(zhuǎn)錄本的3'-UTR 區(qū)域結(jié)合,參與基因剪接的負(fù)調(diào)控[32]。miRNA-99b 存在于精子頂體相關(guān)6(Sperm Acrosome Associated 6,SPACA6)的內(nèi)含子中,LINC01129 是由該基因從反義方向轉(zhuǎn)錄所形成的一種轉(zhuǎn)錄本,miRNA-99b 在SPACA6 的反義方向轉(zhuǎn)錄中起到一定作用,miRNA-99b 與SPACA6第1 個(gè)內(nèi)含子的5'剪接位點(diǎn)完全互補(bǔ)。這種互補(bǔ)性表明miRNA-99b 可通過與U1 snRNPs、U6 snRNPs 競爭性結(jié)合抑制LINC01129 初級(jí)轉(zhuǎn)錄本的表達(dá),從而在SPACA6和LINC01129 之間的表達(dá)平衡中發(fā)揮作用[33]。另外,miRNA-211[34]與DNA 序列的識(shí)別并結(jié)合被證明可以促進(jìn)內(nèi)含子的保留,其中機(jī)制與限制性內(nèi)切酶活性有關(guān)。
RNA 干擾(RNA interference,RNAi)是一種基因沉默機(jī)制,siRNA 是一種與互補(bǔ)序列結(jié)合來抑制基因表達(dá)的長22~24 個(gè)堿基的sncRNA[35]。siRNA 介導(dǎo)的基因沉默分為胞質(zhì)(Posttranscriptional Gene Silencing,PTGS)和胞核(Chromatin-Dependent Gene silencing,CDGS)2 種,其中CDGS 中siRNA 在轉(zhuǎn)錄和共轉(zhuǎn)錄基因調(diào)控中的作用與可變剪接關(guān)系密切[36]。Alló 等[37]研究發(fā)現(xiàn),siRNA 可以與Pre-mRNA 內(nèi)含子與外顯子邊界的剪接位點(diǎn)結(jié)合,調(diào)控外顯子的剪接,在siRNA 結(jié)合位點(diǎn)附近發(fā)現(xiàn)富集有異染色質(zhì)標(biāo)記H3K9me2 和H3K27me3,Pre-mRNA 序列可以通過識(shí)別這些異染色質(zhì)標(biāo)記招募異染色質(zhì)相關(guān)蛋白,從而降低pol II 的合成,這亦是一種剪接相關(guān)的動(dòng)力學(xué)機(jī)制。
1.5.2 長鏈非編碼RNA 與可變剪接 長鏈非編碼RNA(Long non-coding RNA,lncRNA)的長度大于200 nt,但無明顯蛋白質(zhì)編碼功能[38]。根據(jù)lncRNA 的宿主基因在基因組中的位置,lncRNA 可分為基因間lncRNA、天然反義lncRNA 及內(nèi)含子lncRNA[39]。lncRNA 對(duì)可變剪接的調(diào)控主要有3 種方式:與特定剪接因子相互作用調(diào)節(jié)剪接、與Pre-mRNA 形成RNA-RNA 雙鏈調(diào)控剪接、通過染色質(zhì)修飾調(diào)控剪接[40]。
lncRNA 與SR 蛋白以及hnRNPs 等剪接因子相結(jié)合是lncRNA 調(diào)控可變剪接的一條重要途徑[15]。lncRNA通過調(diào)節(jié)SR 蛋白上SR 結(jié)構(gòu)域的磷酸化水平影響SR 蛋白與Pre-mRNA 的結(jié)合,最終導(dǎo)致剪接過程中剪接位點(diǎn)的改變。過氧化物酶體增殖物激活受體γ(Peroxisome Proliferator-Activated Receptorγ,PPARγ)的可變剪接能夠產(chǎn)生2 種轉(zhuǎn)錄本PPARγ1 和PPARγ2,其所編碼的蛋白在脂肪形成過程中起到驅(qū)動(dòng)作用[41]。研究發(fā)現(xiàn)PPARγ的剪接過程受到SR 蛋白SRp40 磷酸化水平的正向調(diào)節(jié)。在脂肪細(xì)胞分化過程中,lncRNA NEAT1 可以與Clk 激酶相互作用調(diào)節(jié)SRp40 的磷酸化水平,影響PPARγ的剪接從而對(duì)脂肪細(xì)胞的分化產(chǎn)生調(diào)控[42]。

圖2 NEAT1 對(duì)PPARγ 剪接影響機(jī)制
lncRNA 通過與Pre-mRNA 形成RNA-RNA 雙鏈調(diào)控內(nèi)含子保留的研究發(fā)現(xiàn),在E 盒結(jié)合鋅指蛋白2(Zinc Finger E-box Binding Homeobox 2,Zeb2)上,該基因是編碼上皮—間質(zhì)轉(zhuǎn)化(Epithelial-Mesenchymal Transition,EMT)的E—鈣黏蛋白的轉(zhuǎn)錄抑制因子[43],Zeb2對(duì)于上皮—間質(zhì)轉(zhuǎn)化受到上皮細(xì)胞中snail1 轉(zhuǎn)錄因子的調(diào)控。Zeb2基因5'—UTR 上的長片段內(nèi)含子存在的核糖體進(jìn)入位點(diǎn)(Internal Ribosome Entry Sites,IRES)對(duì)于蛋白表達(dá)是必需的,通常情況下該基因通過剪接將含有IRES 的內(nèi)含子剪掉從而阻止Zeb2蛋白翻譯,snail1通過反義方向轉(zhuǎn)錄產(chǎn)生的lncRNA 可以與5'-UTR 上的5'剪接位點(diǎn)結(jié)合導(dǎo)致該區(qū)域內(nèi)含子保留,促進(jìn)Zeb2翻譯并最終激活EMT[43]。
人類FGFR2基因反義方向轉(zhuǎn)錄形成的lncRNA 可以通過誘導(dǎo)染色質(zhì)修飾劑對(duì)FGFR2剪接位點(diǎn)附近的染色質(zhì)環(huán)境進(jìn)行修飾來調(diào)控FGFR2的剪接。在FGFR2剪接過程中,該lncRNA 通過與Pre-mRNA 形成RNARNA 異源雙鏈核酸分子招募染色質(zhì)修飾劑KDM2a 對(duì)染色質(zhì)進(jìn)行修飾,所產(chǎn)生的染色質(zhì)環(huán)境可以阻止抑制性剪接因子與Pre-mRNA 序列的結(jié)合,有利于FGFR2外顯子IIIb 的保留[44]。
2 可變剪接在脂肪代謝中的功能
脂肪組織是體內(nèi)重要的內(nèi)分泌組織,脂肪細(xì)胞不僅儲(chǔ)存脂質(zhì),也可以釋放一系列脂肪因子調(diào)節(jié)能量消耗[45],哺乳動(dòng)物脂肪組織根據(jù)形態(tài)、功能以及發(fā)育起源的不同可分為白色脂肪組織(White Adipose Tissue,WAT)和棕色脂肪組織(Brown Adipose Tissue,BAT)[46]。WATs主要功能是把多余脂肪存儲(chǔ)在體內(nèi),過多攝入WATs 便會(huì)產(chǎn)生肥胖;而BATs 負(fù)責(zé)分解引發(fā)肥胖的白色脂肪組織。基因的剪接對(duì)脂肪細(xì)胞的分化以及脂肪代謝具有明顯的調(diào)控作用。該部分主要綜述可變剪接在脂肪代謝調(diào)控中的功能相關(guān)研究進(jìn)展。
2.1 可變剪接與脂肪合成代謝 矮小相關(guān)轉(zhuǎn)錄因子1(Runt-Related Transcription Factor 1,Translocated to 1,RUNX1T1)是參與胚胎干細(xì)胞造血分化的髓系易位基因家族成員[47],可以作為脂肪發(fā)生相關(guān)的轉(zhuǎn)錄因子。RUNX1T1在外顯子6 附近位點(diǎn)不同的剪接形式產(chǎn)生2 種剪接變體[48-49],F(xiàn)TO通過調(diào)節(jié)剪接位點(diǎn)周圍的m6A 水平來控制脂肪生成調(diào)節(jié)因子RUNX1T1的外顯子剪接,從而調(diào)節(jié)分化[48-50]。在小鼠前脂肪細(xì)胞中,外顯子6 缺失轉(zhuǎn)錄本過表達(dá)抑制脂肪細(xì)胞分化,外顯子6 包含轉(zhuǎn)錄本的表達(dá)對(duì)脂肪細(xì)胞分化和脂肪生成具有促進(jìn)作用[13,51]。
絲裂原活化蛋白4 激酶4(Mitogen Activated Protein Kinase Kinase 4,MAP4K4)是一種絲氨酸/蘇氨酸激酶,屬于MAPK 信號(hào)通路上游激活因子,參與細(xì)胞的多種生物學(xué)活動(dòng)[52]。該基因在人體大部分組織中均有表達(dá),其中大腦和睪丸中表達(dá)量更高[53],但其全部功能還尚未完全清楚。最近的一項(xiàng)研究表明,MAP4K4表達(dá)量的增加減弱了胰島素介導(dǎo)的信號(hào)轉(zhuǎn)導(dǎo)參與BATs 的發(fā)育,并與其剪接異常有關(guān)[54]。Peng 等[54]研究發(fā)現(xiàn),在脂肪細(xì)胞分化的不同階段,MAP4K4 的剪接形式均存在差異,其中機(jī)制還需要進(jìn)一步探索。
PPARγ屬于激素配體依賴性核受體家族,該基因控制脂肪酸結(jié)合蛋白4(Fatty Acid Binding Protein4,F(xiàn)ABP4)、脂蛋白脂肪酶以及脂肪細(xì)胞因子等在脂肪細(xì)胞中發(fā)揮關(guān)鍵作用的效應(yīng)蛋白的表達(dá)。PPARγ在調(diào)節(jié)脂肪生成早期階段的轉(zhuǎn)錄事件中起關(guān)鍵作用,對(duì)于實(shí)現(xiàn)成熟脂肪細(xì)胞的功能也是必不可少的[41],PPARγ基因利用不同剪接機(jī)制產(chǎn)生PPARγ1 和PPARγ2,這2 種轉(zhuǎn)錄本由不同的5'-UTR 和6 個(gè)編碼外顯子組成,其中PPARγ1在肝臟、心臟、骨骼肌和白色脂肪組織中廣泛表達(dá),而PPARγ2 在N 末端包含28 個(gè)額外的氨基酸,主要存在于脂肪組織中[55]。體外的功能試驗(yàn)表明,不管是PPARγ1還是PPARγ2 對(duì)于脂肪生成都是必須的,PPARγ2 在棕色脂肪生成中是一種更有效的轉(zhuǎn)錄激活因子。
2.2 可變剪接與脂肪分解代謝 脂肪分解是在一系列脂肪分解酶的作用下對(duì)細(xì)胞內(nèi)脂質(zhì)進(jìn)行水解的過程,該過程對(duì)于體內(nèi)的能量供給以及維持體內(nèi)脂肪儲(chǔ)存具有重要意義。目前對(duì)于可變剪接在脂肪分解代謝中的功能研究還相對(duì)較少,但是已經(jīng)有證據(jù)證明基因可以通過可變剪接對(duì)脂肪分解產(chǎn)生一定的調(diào)控作用。激素敏感脂肪酶(Hormone-Sensitive Triglayceride Lipase,HSL)是細(xì)胞內(nèi)脂質(zhì)水解的關(guān)鍵限速酶,其表達(dá)變化能在一定程度上反映脂肪的分解能力[64]。hHSL-S 是人脂肪細(xì)胞HSL剪接時(shí)外顯子6 跳躍形成的新轉(zhuǎn)錄本。Laurell 等[65]在COS 細(xì)胞中過表達(dá)hHSL-S 發(fā)現(xiàn)該細(xì)胞缺乏脂肪水解酶活性,說明HSL 的剪接異常會(huì)影響細(xì)胞脂肪水解酶活性從而抑制脂肪分解。肉堿脂酰轉(zhuǎn)移酶1(Carnitine Palmitoyl Transferase I,CPT1)是肝臟組織細(xì)胞中長鏈脂肪酸β-氧化的關(guān)鍵調(diào)節(jié)酶和限速酶,該基因具有CPT1A、CPT1B、CPT1C 等多種轉(zhuǎn)錄本[66]。在乳腺癌細(xì)胞中,CPT1A 的表達(dá)加速了脂肪酸β氧化為細(xì)胞提供能量,促進(jìn)乳腺癌細(xì)胞增殖[67]。PD-1 可以通過抑制糖酵解并促進(jìn)脂肪分解和脂肪酸氧化來誘導(dǎo)T 細(xì)胞代謝,其對(duì)脂肪酸氧化的調(diào)節(jié)是通過增加CPT1A 的表達(dá)來實(shí)現(xiàn)的[68]。在果蠅體細(xì)胞中存在的CPT1某種轉(zhuǎn)錄本導(dǎo)致該細(xì)胞脂酶活性降低,這對(duì)于脂質(zhì)分解具有減緩作用[69],這些研究表明了可變剪接在脂肪分解中的重要調(diào)控作用。
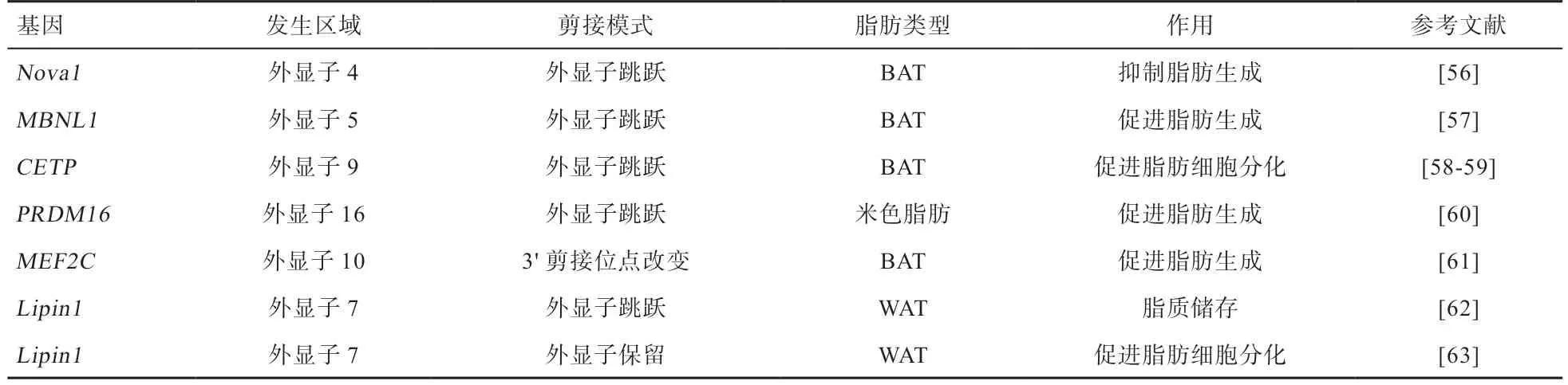
表1 脂肪合成相關(guān)可變剪接事件
3 總結(jié)與展望
近年來,已經(jīng)有大量證據(jù)證明表觀遺傳學(xué)修飾與Pre-mRNA 的剪接之間存在互作,并且對(duì)于表觀遺傳學(xué)與Pre-mRNA 之間的動(dòng)態(tài)關(guān)系正在取得重大進(jìn)展,這為全面了解復(fù)雜的基因表達(dá)調(diào)控網(wǎng)絡(luò)提供了新的見解。脂肪的代謝不僅與人類健康息息相關(guān),而且對(duì)于畜禽的經(jīng)濟(jì)性狀具有重要意義,深入了解脂肪的代謝調(diào)控機(jī)制對(duì)于生命科學(xué)研究的推進(jìn)也具有一定意義。本文主要總結(jié)了可變剪接在脂肪代謝調(diào)控中的功能相關(guān)研究進(jìn)展,然而目前的研究大都停留在研究脂肪代謝調(diào)控相關(guān)的基因,對(duì)于更深入地了解相關(guān)調(diào)控因子與剪接缺乏進(jìn)一步研究。
隨著組學(xué)時(shí)代的不斷發(fā)展,轉(zhuǎn)錄組測(cè)序技術(shù)已經(jīng)發(fā)展成熟并且成為了生命科學(xué)領(lǐng)域研究的重要工具。RNA-seq 能夠以效益高、時(shí)間短的方式產(chǎn)生更多的基因組序列,結(jié)合相關(guān)的生信分析手段可以實(shí)現(xiàn)在全基因組水平分析各種細(xì)胞不同生理?xiàng)l件下的可變剪接事件,為進(jìn)一步探索脂肪代謝相關(guān)剪接及其調(diào)控因子提供了便利,將有助于人類肥胖相關(guān)疾病的治療以及經(jīng)濟(jì)動(dòng)物的肉質(zhì)性狀改良。
