腸道微生物及代謝產物對動物腸道免疫的研究進展
2021-03-01 11:34:32鐘奇祺張海波幸清鳳關瑋琨廖曉鵬黎力之郭冬生
中國畜牧雜志 2021年2期
關鍵詞:小鼠
鐘奇祺 ,張海波,幸清鳳,關瑋琨,廖曉鵬,黎力之*,郭冬生*
(1.宜春學院生命科學與資源環境學院,江西省高等學校硒農業工程技術研究中心,宜春市功能農業與生態環境重點實驗室,江西宜春 336000;2.宜春學院計劃財務處,江西宜春 336000;3.宜春學院繼續教育學院,江西宜春 336000)
哺乳動物腸道定植約1014個微生物,包括細菌、真菌、病毒等[1]。腸道微生物在營養物質的消化吸收過程中發揮重要作用,并促進機體免疫系統的建立[2]。缺少腸道微生物定植的宿主無法建立功能健全的免疫系統。研究表明,無菌小鼠的多個免疫細胞群均表現出缺陷,如先天淋巴細胞功能受損、產生免疫球蛋白A(IgA)的漿細胞存在功能障礙等[3-4]。健全免疫系統與腸道微生物間的關系已被證實,如分節絲狀菌(Segmented filamentous bacteria,SFB)誘導小鼠CD4+T 細胞激活輔助性T 細胞17(Th17 細胞)產生白細胞介素(IL)-17和IL-22,保護宿主免受細菌和真菌感染[5];脆弱擬桿菌(Bacteroides fragilis,BF)產生特有的抑制性鞘脂類物質對機體內源性抗原環境進行補充,維持宿主恒定型自然殺傷T 細胞的穩態平衡[6]。腸道微生物定植的同時,經自身轉化得到其代謝產物,如短鏈脂肪酸(SCFAs)、次級膽汁酸和離子多糖A(PSA)等。這些代謝產物直接通過黏膜層激活腸上皮細胞受體,如腸上皮細胞G 蛋白偶聯受體(GPRs)介導的信號通路被啟用后,刺激調節性T 細胞(Treg 細胞)轉錄程序,促進抗炎細胞因子合成釋放,提高機體免疫耐受能力。而這樣的代謝產物受體存在于不同的腸上皮細胞中,表明代謝產物廣泛作用于機體免疫過程[7]。因此,腸道微生物及其代謝產物對宿主免疫系統調控起至關重要的作用。本文主要綜述了腸道微生物及其代謝產物SCFAs、次級膽汁酸和PSA 對宿主腸道免疫的影響,以期為調控腸道免疫、治療動物疾病及保障機體健康提供思路和參考。
1 腸道微生物及代謝產物
1.1 腸道微生物的分布 在已鑒定的腸道菌群中,擬桿菌門和厚壁菌門的數量占腸道菌群總量90% 以上,其中專性厭氧菌數量約是需氧菌數量的1 000 倍[8]。由于不同腸段內環境不一樣,腸腔中微生物數量與組成均有差異。十二指腸中的微生物數量最少,主要原因是胃酸與膽汁排出后首先進入其中,導致內環境pH 較低,且過多的腸液和腸段快速蠕動也不利于細菌定植[9]。與十二指腸相比,空腸和回腸中的腸液更少,腸段蠕動速度減緩,氧化還原電位降低,促進厭氧菌定植,故空腸和回腸內的腸道微生物數量有所增加[10]。盲腸具有遠端閉塞不通、腸段蠕動速度緩慢和pH 呈弱堿性等特點,是細菌定植的主要場所,其中優勢菌群為雙歧桿菌(Bifidobacterium)、腸桿菌(Enterobacteriaceae)和腸球菌(Enterococcus)[9]。
1.2 腸道微生物產生的代謝產物 動物腸道無法直接消化吸收的飲食成分,經腸道微生物作用后,轉化為宿主代謝中的功能性補充物質[1]。微生物代謝產物分為3 類:第一類經腸道微生物直接對食物成分進行消化或發酵產生,如SCFAs[11]。SCFAs 由小腸厭氧微生物發酵結構性碳水化合物生成,具有促進腸道黏液分泌、提供結腸肌細胞能量等作用[12]。第二類由宿主產生后經腸道微生物修飾后的代謝產物,如次級膽汁酸[2]。次級膽汁酸由初級膽汁酸經腸內7-α/β脫羥基細菌解離、轉化而來,有助于動物構建腸道環境穩態[13]。第三類為腸道微生物自生合成的代謝產物,如PSA[14]。PSA 主要由BF 產生,是一種莢膜多糖,通過激活Toll 樣受體2(TLR2)信號通路誘導Treg 細胞產生具有抗炎作用的IL-10,抑制胃腸道炎癥的發生[15]。綜上所述,微生物可以通過其代謝產物調節宿主胃腸道免疫系統的正常生理功能(表1)。
2 腸道微生物對免疫細胞和免疫球蛋白的影響
2.1 腸道微生物對分泌型球蛋白A(sIgA)分泌的影響腸道微生物通過依賴T 細胞和非依賴T 細胞2 種方式調控sIgA 的產生(圖1)[22]。依賴T 細胞的sIgA 產生主要發生在派爾集合淋巴結中,其通過B 細胞與負載抗原的DCs 相互作用后產生sIgA[23]。捕獲抗原的DCs 刺激CD4+T 細胞增殖分化為濾泡輔助性T(Tfh)細胞,Tfh 細胞主要以CD40 配體和IL-21 作為信號,誘使B細胞表達胞嘧啶核苷脫氨酶(AID),促進B 細胞分化為IgA+B 細胞,進而分泌大量sIgA[24]。非依賴T 細胞的sIgA 產生主要在腸黏膜固有層和孤立淋巴濾泡中。腸道菌群可通過觸發腸上皮細胞中TLRs 直接刺激DCs和濾泡樹突狀細胞產生增殖誘導配體(APRLI)和B 細胞的活化因子(BAFF),誘導B 細胞中的AID 表達和IgA 類抗體轉換,提高B 細胞轉化為IgA+B 細胞的速率,顯著增強sIgA 合成能力[24]。

圖1 腸道微生物對sIgA 分泌的影響[22]

表1 腸道微生物產生的黏膜免疫調節代謝產物
腸黏膜固有層中IgA+漿細胞產生二聚體IgA(dIgA),dIgA 相互聚合形成多聚體IgA(pIgA),隨后與多聚免疫球蛋白受體(pIgR)結合轉運至腸上皮表面頂端,之后dIgA 與pIgR 中的分泌成分從結合物中分離,形成sIgA[25]。sIgA 接觸腸腔內微生物、膳食成分等物質后,在這些物質表面形成一層外膜,抑制腸腔致病菌直接接觸腸壁,使其無法黏附腸上皮細胞失去定植能力,從而維持機體腸道屏障的完整性,抵御其他病原微生物入侵[22,26]。
2.2 腸道微生物對T 細胞的影響 腸腔內特定的共生細菌可以促進T 細胞不同亞群的生成。抗原刺激使固有層中CD8+T 細胞向CD4+T 細胞增殖分化后,CD4+T 細胞又可特異性增殖分化為不同亞群,如Treg 細胞、Tfh細胞和Th17 細胞等。其中,Treg 細胞的增殖分化可以減輕因機體對致病菌的過度免疫反應而引起的黏膜損傷,如BF 誘導Treg 細胞增殖分化產生IL-10,保護腸道免受肝螺桿菌(Helicobacter hepaticus)入侵感染[27]。Th17 細胞具有介導炎癥反應的作用,其與Treg 細胞在功能上具有相互制約的作用。近年來大量研究表明,通過腸道微生物誘導產生的Treg 細胞與Th17 細胞可以維持腸道在免疫耐受與炎癥反應之間的平衡,有利于建立腸黏膜免疫穩態[28-31]。
2.2.1 腸道微生物對Treg 細胞增殖分化的影響 Treg細胞來源于胸腺和外周免疫器官,其增殖分化主要受CD25 和叉頭狀螺旋轉錄因子3(FoxP3)分子調控。腸道Treg 細胞存在于腸黏膜和淋巴組織中,抑制T 細胞過度反應,調節自身免疫。外周Treg 細胞(pTreg)聚集于結腸,常見表達形式為RORγt。無菌小鼠體內缺乏RORγt+pTreg 細胞,說明微生物可以誘導產生pTreg 細胞[32]。Atarashi 等[33]將正常人體17 種梭菌混合,定植于無菌小鼠體內,結果顯示pTreg 細胞水平顯著升高,促進IL-10 和細胞毒T 淋巴細胞相關抗原4 的合成釋放,共同抵御小鼠結腸炎。研究表明,BF 可以分泌外膜囊泡,通過非典型自噬途徑向免疫細胞傳遞免疫調節分子,誘導產生IL-10,表達FoxP3+細胞,促使CD4+T 細胞向Treg 細胞增殖分化,抑制結腸炎[34]。
2.2.2 腸道微生物對Th17 細胞增殖分化的影響 Th17細胞的應答時間早于其他T 細胞,且其可以在腸道內積聚,這表明Th17 細胞的發育由腸內機制調節[32]。與此相契合的是,使用抗生素處理后的小鼠或者無菌小鼠,腸內Th17 細胞數量均急劇下降。因此,腸道微生物在調控Th17 細胞的增殖分化中具有重要作用[29]。研究表明,腸道中SFB 具有鉤狀結構,鉤尖黏附小鼠腸上皮細胞形成內吞囊泡,可快速將SFB 的菌體蛋白從腸上皮細胞的頂端運送至基部,直接促使血清淀粉樣A 蛋白(SAA)產生,進而使黏膜固有層中DCs 產生IL-6 和IL-23,誘導Th17 細胞增殖分化[33,35]。同時,Mao 等[36]證明SFB 定植腸道后產生IL-23,刺激固有淋巴細胞(ILCs)與Th17 細胞共同分泌IL-22,間接上調SAA1和SAA2基因表達,并激活MAPK 通路釋放抗菌肽和β防御素,保證腸道屏障結構與功能的完整性,阻礙病原微生物入侵機體。除SFB 外,腸內白色念珠菌(Candida albicans)、鼠檸檬酸桿菌(Citrobacter ratus)及腸出血性大腸桿菌(Escherichia coli O157:H7)等致病菌均可黏附腸上皮促使Th17 細胞分化[37-38]。由此看來,腸道微生物可以通過對腸上皮細胞的黏附作用誘導T 細胞向Th17 細胞增殖分化,對機體炎癥迅速做出反應,加強宿主對病原微生物的防御作用(圖2)。
3 腸道微生物代謝產物與腸黏膜免疫
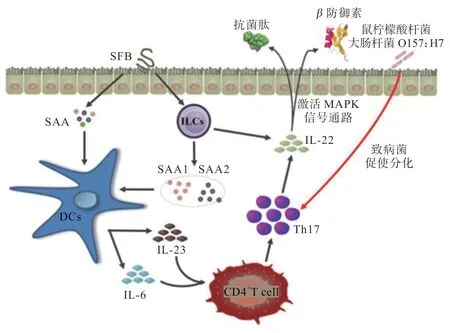
圖2 腸道微生物對Th17 細胞增殖分化的影響
3.1 SCFAs 與腸黏膜免疫 SCFAs 包括乙酸、丙酸、丁酸等,SCFAs 不僅是微生物本身的主要能量來源,還為腸上皮細胞提供能量。此外,SCFAs 還能抑制組蛋白去乙酰酶(HDACs)活性,促使腸上皮細胞緊密連接,參與調節機體免疫功能。SCFAs 通過抑制HDACs 活性影響Treg 細胞發育。HDAC9 的抑制作用使FoxP3 表達增加,促使Treg 細胞增殖分化,增強Treg 細胞介導的結腸炎抑制作用[39]。研究表明,SCFAs 與游離脂肪酸受體2(FFAR2)特異性結合后減弱腸道中性粒細胞的趨化作用,降低脂多糖刺激下中性粒細胞產生的促炎細胞因子水平[40]。Zhou 等[28]研究發現脂多糖刺激巨噬細胞后,丁酸能限制促炎細胞因子在結腸固有層中合成與分泌,降低腸道炎癥的發生幾率,同時結腸隱窩存在丁酸代謝,防止高濃度丁酸鹽對隱窩底部干細胞產生毒害效應,維護動物腸黏膜免疫細胞的更新能力。
SCFAs 通過完善腸上皮細胞屏障功能維持黏膜免疫。Wrzosek 等[41]研究發現,無菌小鼠接種產生SCFAs 的泰氏桿菌(Bacteroides thetaiotaomicron)和糞便桿菌(Faecalibacterium),能有效誘導杯狀細胞分化,促進黏液產生,有效阻止致病菌定植腸道,保障腸道健康。SCFAs 還能穩定腸上皮細胞連接穩定,維護腸道屏障完整。定植長雙歧桿菌(Bifidobacterium longum)產生高水平乙酸,增強腸上皮細胞間相互連接,提高腸道對病原體的防御能力[16]。
3.2 次級膽汁酸與腸黏膜免疫 膽汁酸由肝臟合成儲存于膽囊,動物采食后膽囊收縮,膽汁酸被釋放進入十二指腸,稱為初級膽汁酸[42]。腸道腔內乳酸桿菌、雙歧桿菌等利用膽鹽水解酶對初級膽汁酸進行早期解離后,再與具bai基因的梭菌發生7-α脫羥基反應生成次級膽汁酸,包括石膽酸(LCA)、脫氧膽酸(DCA)和熊脫氧膽酸(UDCA)等[18]。研究證明,患炎癥性腸病(IBD)的小鼠體內厚壁菌門數量減少,阻礙膽汁酸早期解離、轉化及脫硫過程,顯著降低次級膽汁酸轉化率,導致3-OH 硫化膽汁酸積聚腸內,加重炎癥反應[43]。Den Bossche 等[44]將UDCA 灌胃于葡聚糖硫酸鈉誘導IBD 小鼠后,使腸內厚壁菌門與擬桿菌門比例恢復正常,并有效降低小鼠腸道促炎細胞因子(IL-1β、IL-6)水平,加速潰瘍化上皮愈合,表明次級膽汁酸具備一定免疫調節作用。此外,研究表明,LCA 與DCA 有效抑制80%腸道白色念珠菌菌絲、芽管及生物膜的形成,阻止其生長繁殖,維持動物腸道健康[45]。Jason 等[19]試驗證明,LCA 和DCA 可增強腸內7-α脫羥基細菌所分泌抗生素的活性,解除外源抗生素治療艱難梭菌后產生的環狀二肽抑制其他菌群生長,阻止艱難梭菌異常增殖所導致的腸黏膜免疫極化現象出現。同時,次級膽汁酸作為膽汁酸家族一員,也具備激活法尼醇X 受體及G 蛋白偶聯膽汁酸受體5 通路的功能,發揮維持腸道穩態及黏膜屏障完整性的作用[18]。綜上所述,適宜濃度的次級膽汁酸對機體腸黏膜免疫建立具有重要意義。
3.3 PSA 與腸黏膜免疫 PSA 為囊狀兩性離子多糖,主要由BF 產生,常見于結腸黏液層,通過激活DCs和Treg 細胞合成分泌抗炎細胞因子,調控宿主免疫系統功能。一方面,PSA 依賴腸壁通透性進入固有層,被DCs 內化吸收后將抗原呈遞至CD4+T 細胞,導致其分泌IL-10,誘使CD45+細胞釋放IL-17A,抑制腸道T 細胞活性,尤其是Th17 細胞,繼而阻止炎癥反應發生,保護罹患結腸炎小鼠的生理健康[31]。另一方面,DCs 細胞表面的TLR2 也能被PSA 激活,促使T 細胞釋放IL-10[20-21]。此外,PSA 還能刺激CD4+T 細胞向FoxP3CD4+T 細胞或FoxP3-CD4+Tr1 細胞方向分化,與細胞表面TLR2 特異性結合,促進IL-10 的合成,緩解結腸炎[15]。綜上所述,PSA 通過直、間接方式作用于CD4+T 細胞,促進IL-10 分泌,抑制Th17 細胞過度免疫應答,輔助動物抵御腸道疾病(圖3)。
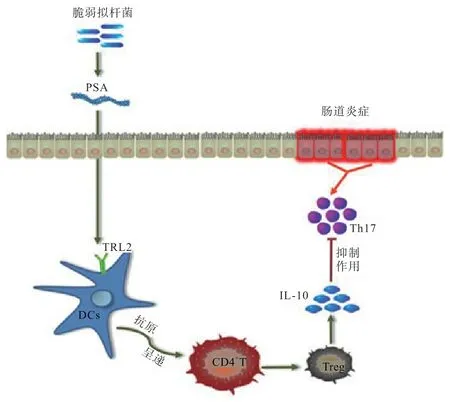
圖3 PSA 對腸黏膜免疫的影響
4 結 語
綜上所述,腸道微生物定植后調節sIgA 分泌及T細胞增殖,完善動物腸道免疫功能,同時產生大量代謝產物(SCFAs、次級膽汁酸和PSA 等),在維持腸道穩態、輔助動物抵御疾病方面具有重要作用。腸道微生物及其代謝產物對動物腸道免疫的影響已被大量研究和闡釋,但菌群種類及定植過程復雜,現有研究大多為孤立、單一角度的驗證,仍不足以解釋菌群與腸道免疫間復雜的動態關系。因此,腸道微生物及其代謝產物對宿主腸道免疫系統作用機制有待全面深入探索,相關研究也將為豐富機體腸內外疾病的防治理論提供參考依據。
猜你喜歡
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國男科學雜志(2016年4期)2016-12-01 05:20:09
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
