東北地區李屬(Prunes L.)植物導管分子形態結構研究
2021-02-26 07:49:54鄒子瑜谷利偉張大維
植物研究 2021年1期
關鍵詞:植物
鄒子瑜 谷利偉 張大維*
(1. 哈爾濱師范大學,哈爾濱 150025;2. 黑龍江八一農墾大學,大慶 163319)
李屬(Prunes L.)隸屬于薔薇科(Rosaceae)李亞科(Prunoideae)。該屬植物全球約有150 余種,分布在除澳洲外各地,我國有50余種,全國均有分布;其中東北地區產10種,栽培15種[1]。李屬植物具有較高的經濟價值,桃(P.persica L.)、杏(P.vul?garis Lam.)、櫻 桃(P.pseudocerasus(Lindl.)G.Don)等都是生活中常見的水果。稠李(P.racemosa(Lam.)Gilib.)、榆葉梅(P.triloba(Lindl.)Ricker)、櫻花(P.yedoensis(Matsum.)Yu et Li)等具有較高的觀賞價值。李屬同時具有一定的學術價值:近年對李亞科各屬的分類標準一直存在分歧:一種是“大屬”分類法,1926 年由德國科學家A.Rehder提出,將核果類植物合并成李屬,并將李屬分為五個亞屬:李亞屬、扁桃亞屬、櫻亞屬、稠李亞屬與桂櫻亞屬,這種分類方法國內外采用者很多[2],本文也采用這種分類方法。另一種是由我國學者俞德浚提出的,將李亞科分為9 個小屬。其中桃屬(Amygdalus L.)最為進化,稠李屬(Padus Mill.)與桂櫻屬(Laurocerasus Tourn. ex Duh.)相對最原始[3]。早期對李屬植物的形態學研究以木材解剖學為主:認為李亞科和蘋果亞科是薔薇科中較為進化的亞科,且李亞科獨立分支較早。并與蘋果亞科親緣較遠[4~7]。隨著近年來分子生物學的發展,對李屬植物的分類研究也深入到了分子層面。Ma R.C.在對梨屬與李屬S-RNases cDNA 序列的研究中發現,扁桃仁(P.communis L.)與甜櫻桃(P.avi?um(L.)Moench.)的S-RNases 基因沒有特異性差異。這也可以成為李亞科大屬分類的分子生物學佐證[8]。由于李亞科分類學仍存在分歧,而導管分子既存在一定微觀分類學意義,又是反應生境狀態的良好指標。因此,對薔薇科植物導管分子的研究成為近年來的研究熱點。我國學者尹秀玲曾對薔薇科9 屬12 種代表植物的導管分子進行觀察,并找出了薔薇科導管分子共性特征與屬間差異[9]。郭學民等人發現李屬砧木莖段到接穗莖段出現了導管寬度不匹配,說明即使為同屬植物,不同種植物的導管直徑在同生境下也存在著差異[10]。胡蓉在比較日本晚櫻與櫻桃導管分子的研究中發現,日本晚櫻較櫻桃更為進化,但他們個各性狀在系統演化中不同步[11]。近年來對其他科屬植物導管分子的研究以導管分子生理結構和不同植物導管分子形態結構差異為主。高建平、王彥涵等通過導管分子形態特征研究與年降水量比較,得出其與導管分子長度與直徑的比值相關[12]。張大維等在研究樺木科(Betulaceae)與槭樹科(Ac?eraceae)槭屬(Acer Linn.)植物導管分子的過程中發現同科同屬植物在結構形態上有一定共性,并發現導管分子的形態特征可能與環境因素有關[13~14]。
然而現階段國內外對李屬植物導管的分子的研究基本都停留于種間結構對比,并且同種植物沒有在不同生境下進行結構與形態的分析。因此,本文選取了東北地區10 種當地自然生長的李屬植物,并在不同生境多點采樣。利用掃描電子顯微鏡進行觀察,在研究導管分子結構的同時分析了生境因素對導管的影響,提供了微形態學證據。
1 材料和方法
1.1 材料
實驗材料均采自中國東北地區自然環境生長的李屬植物枝條(見表1)。選取一年生枝條的節間。每種植物分別采集自3個不同地理位置的生境下。
1.2 實驗方法
采用植物導管樹脂鑄型法對導管分子內壁形態結構特征進行觀察和測量。1 年生枝條自然干燥后,將周皮處理干凈,切成10 mm×5 mm大小,浸泡于苯乙烯中放于真空抽氣泵中抽出存留在輸導組織中的空氣,再浸泡4~6 周。滲透過苯乙烯的實驗材料于70℃溫箱中進行高溫聚合,直到完全凝固為止。取出凝固好的材料,浸泡在以下2種溶液中使有機材料分解:①30%的過氧化氫和99.5%的冰醋酸1∶1的混合液。在60℃下浸泡1~2 h。②浸泡在98%的濃硫酸中10 min。更換溶液時需用蒸餾水沖洗后再浸泡在另一溶液中。兩步驟輪流操作3~5 次,直至復型材料充分分散。暴露出的復型材料在70%的酒精中保存以待使用。樣品材料經過完全處理后,稀釋在70%的酒精中。
1.3 觀察與測量
在解剖鏡下用牙簽、解剖針等較尖的用具將處理好的導管離析材料粘貼于掃描電鏡的樣品臺上。再置于離子濺射儀中進行噴金導電處理,并在HITACHI S-4800 型掃描電子顯微鏡下(加速電壓20 kV)觀察。隨機在掃描電鏡的樣品臺上選擇30個導管分子管長、尾長、寬度及端壁長度進行測量。根據端壁長度與導管寬度計算傾斜角正弦值,根據正弦值換算成角度(即角度數值越大面積越小)。
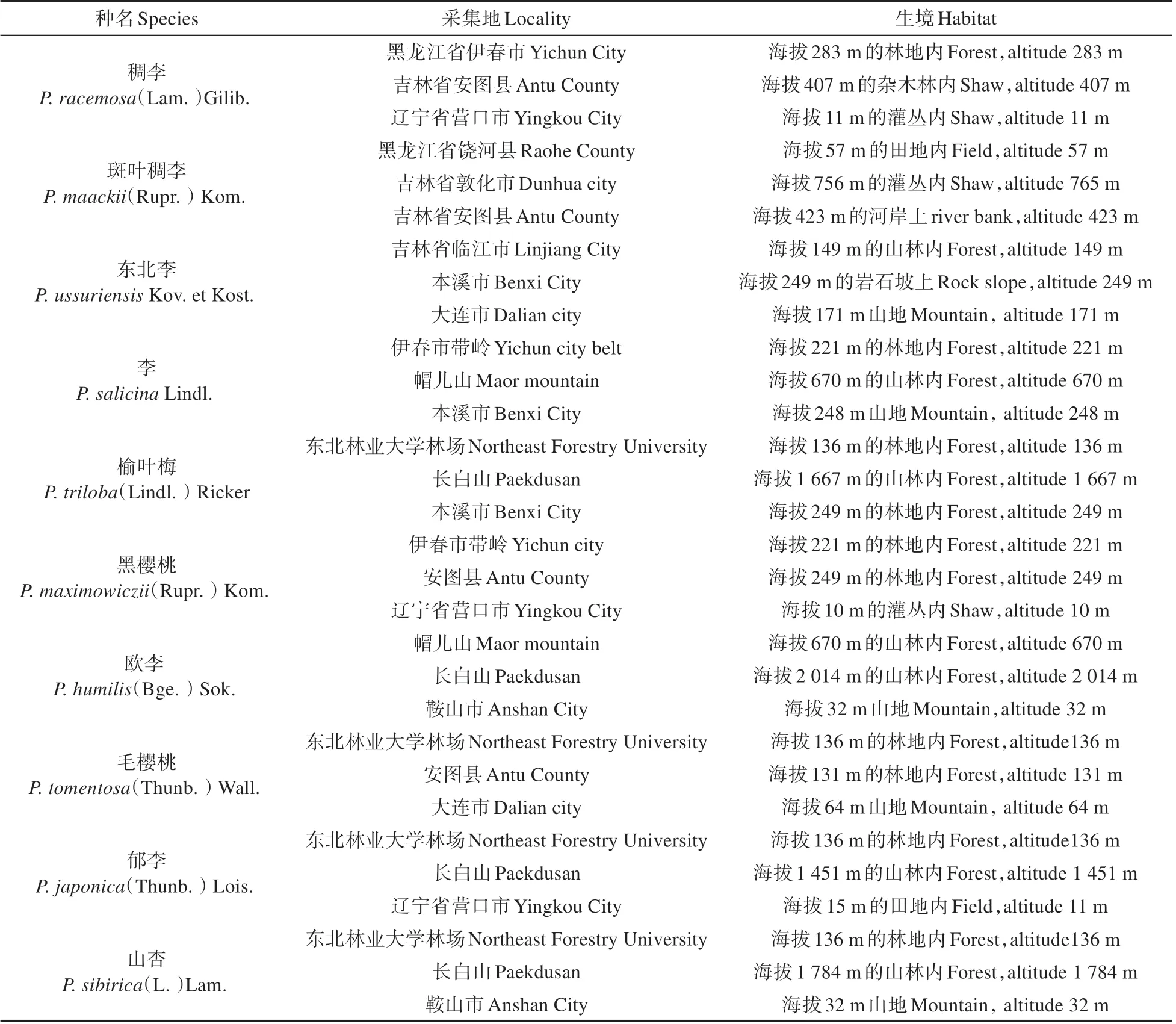
表1 實驗材料信息Table 1 Experimental material information
1.4 數據分析方法
本文利用spss 軟件對每組數據進行差異性分析與標準差分析。
2 觀察結果
不同生境下的李屬植物導管分子的管腔長度、寬度及端壁斜度角有較大的差別(見圖1)。端壁傾斜角為端壁與直徑的夾角度數。角度數值越小,說明端壁越傾斜,端壁面積越大。李屬植物導管分子的類型有網紋、孔紋及螺紋3種形態;紋孔排列方式出現2 種分別是:對列、互列同時存在和僅存在互列式。李屬植物導管分子均存在由管分子次生壁內側具有螺旋狀隆起的螺紋加厚(見表2)。由于同種植物不同生境下導管類型與結構特征并沒有變化,故不區分采集地,僅以種名分類列表。
2.1 導管類型與穿孔板類型
研究中李屬植物導管分子的導管形態類型共有3 種,網紋導管與孔紋導管普遍存在。僅山杏、歐李、東北李3種李屬植物的導管分子中沒有發現螺紋導管(見圖版Ⅰ:1)。李屬植物中普遍存在較為進化的孔紋導管。單穿孔板在該屬孔紋導管中普遍存在,僅在櫻亞屬的黑櫻桃、毛櫻桃與郁李中發現梯狀穿孔板。
2.2 紋孔的排列方式
李屬植物導管分子紋孔排列方式為互列式的情況在所觀察的植物中普遍存在,其中山杏、稠李、東北李以及毛櫻桃為對列式、互列式同時存在(見圖版Ⅰ:5~7)。其余6 種僅存在對列式(見圖版Ⅰ:8~9)。
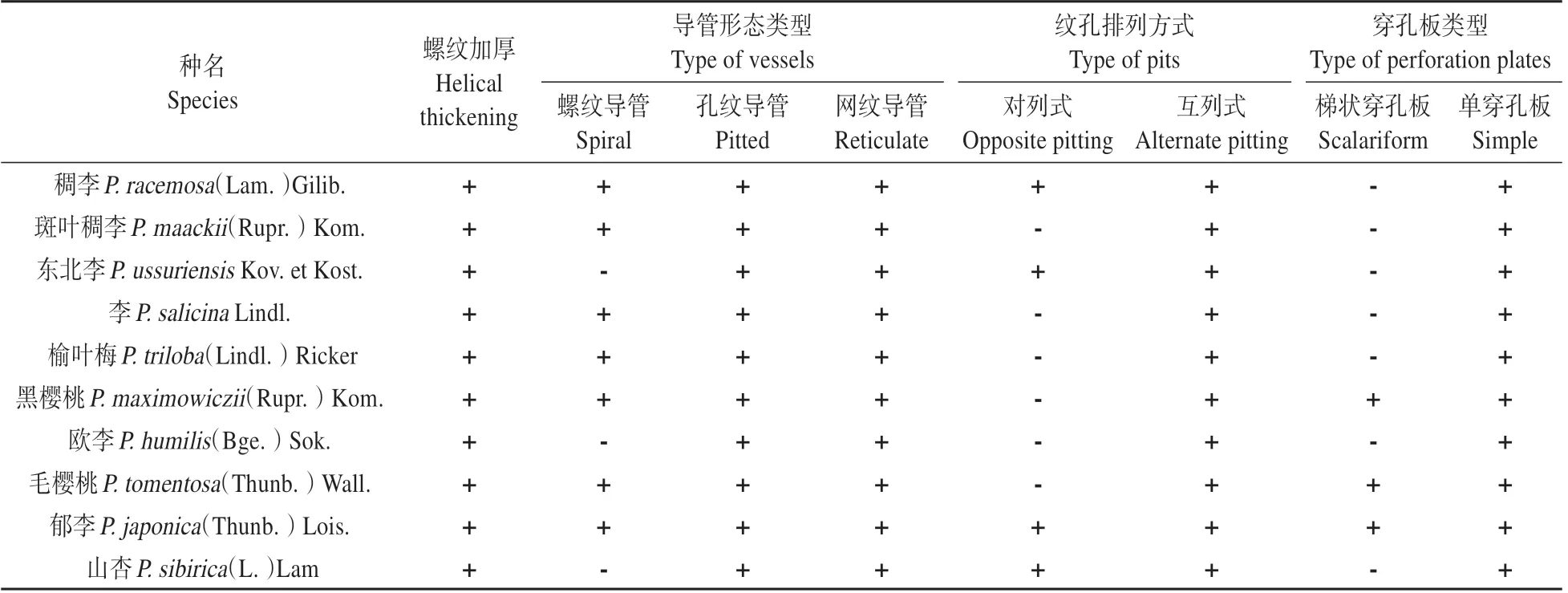
表2 東北地區植物導管分子形態特征Table 2 morphological characteristics of vessel elements from Northeast of China
2.3 導管管壁的螺紋加厚
所觀察的李屬植物導管分子的內表面均呈螺紋加厚(見圖版Ⅰ:2)。螺紋型加厚會增加導管內壁的表面積,這可以提高導管對水分的附著力,從而提高水分運輸效率,但不同生境下李屬植物均存在螺紋加厚(見圖版Ⅰ:4),且尾端也具有螺紋加厚與紋孔對(見圖版Ⅰ:3)。
2.4 李屬植物導管分子長度、端壁斜度角、寬度及尾端長度
所觀察的10 種李屬植物分別選擇3 個不同的地區取樣,共計30組樣本。從數據分析可知,導管長度、端壁傾斜角與海拔和緯度正相關。并且在對榆葉梅、歐李、郁李跟山杏4 種植物的觀察中發現,生長在長白山1 500 m 左右海拔的植物,緯度低于黑龍江地區,但導管長度與端壁角度與黑龍江地區的植物無顯著差異,甚至高于黑龍江地區。導管寬度與海拔緯度呈負相關,即海拔、緯度越高導管寬度(即直徑)越小。在對東北李的觀察中發現,在海拔相對一致,緯度相對靠近的情況下,管長、寬度與端壁傾斜角度均無顯著差異。在所觀察不同生境下的李屬植物中,尾端長度在種內相對穩定,無顯著差異,但種間仍存在差異。
3 討論
植物導管分子形態結構觀察結果表明,同種植物在不同生境下形態(長度、寬度、端壁面積)上存在差異。但管壁螺紋加厚、尾端長度、導管類型,穿孔板類型以及紋孔式并沒有出現變化。其中孔紋、網紋導管、螺紋加厚、互列紋孔式與單穿孔板是李屬植物普遍存在的結構特征。梯狀穿孔板僅在櫻亞屬植物中被觀察到。但相對原始的螺紋導管存在在3種不同亞屬的植物中。因此,我們認為導管分子的各結構的進化程度有所不同。對尾端長度的觀察發現,同種植物尾端在不同生境下長度無變化,這說明尾端長度僅與進化程度相關。螺紋加厚在李屬中普遍存在。內壁螺紋加厚是一種次生壁內側產生的規則性凸出,它可以增強管壁對水的附著力,從而提高植物對水分的利用效率[15~16]。由于所觀察的李屬植物無論生境如何均存在螺紋加厚,所以推測這一形態應該是李屬植物長期適應旱生環境而演化出來的,但這仍需要進一步研究。在對長度、寬度與端壁面積的測量中發現,李屬植物在不同生境下導管分子會對環境產生一定的適應性變化。緯度與海拔越高,長度越長、寬度越小、端壁面積越小。這種變化是個體植物對環境適應的反饋,并不是適應環境而演化的結果。因此,我們認為李屬植物導管分子的微形態結構具有十分重要的意義,既可以研究李屬的系統學地位,又可以為李亞科的分類方法提供佐證。同時在對導管分子的環境適應性方面提供重要的參考依據。
導管的出現,是植物為適應陸地生境的一個重要標志。目前,國內外學者發現導管分子的形態結構具有較重要的系統發育學地位,導管分子中穿孔板的形態特征、導管類型及尾端長度是顯示類群演化地位的重要微觀指標[17~18]。梯狀穿孔板作為較原始的穿孔板類型,常出現在較為原始的植物類群中,如毛茛科(Ranunculaeeae)等[19]。但在薔薇科中較為進化的李屬中仍有少數種存在梯狀穿孔板(黑櫻桃、毛櫻桃與郁李)。且這些種都隸屬于李屬的櫻亞屬,但櫻亞屬的植物也存在較為進化的單穿孔板。因此,我們認為櫻亞屬的導管是東北地區李屬中相對進化,但最為原始。由于導管分子管壁發育的先后和導管分子次生壁增厚的位置、方式不同,從而形成了不同類型的導管。螺紋、環紋導管常出現在植物發育的早期,而網紋與孔紋導管則出現在較為成熟的植物發育時期。并且國內外研究也表明導管類型也是分類學與系統發育學的重要指征[5,9,19]:環紋導管與螺紋導管較原始;網紋、孔紋導管則相對進化;梯紋導管則是較為原始類型的導管分子向孔紋導管進化的過渡類型。并且在所觀察的植物中發現:不同生境下的同種植物導管類型相同,這與前人的研究相符合[20]。所觀察的李屬植物均存在孔紋導管與網紋導管;絕大多數李屬植物均存在螺紋導管(僅東北李、歐李與山杏中沒有)。這說明李屬植物的導管類型以最進化的孔紋導管為主。多數種仍保留著較為原始、輸水功能較弱的螺紋導管。歐李中既不存在螺紋導管,又沒有發現梯狀穿孔板。因此,我們認為歐李在李屬中相對進化。本研究測量的尾端長度也支持這一結果。從導管類型與穿孔板類型來看李亞屬與杏亞屬相對較為進化,這也符合前人的結論。但他們仍存在相對過渡類型的紋孔式,即對、互列同時存在的紋孔排列方式。孔紋導管的紋孔式演化方式是由對列式向互列式方向進行的[21]。李屬植物孔紋導管的演化已經處于較為進化的階段,但尚存少許互列式紋孔同時說明著仍處于較為初級的進化階段。綜合來看,櫻亞屬的郁李中觀察到了所有的原始結構特征,這顯示著郁李是李屬最原始的種。因此,我們認為李屬植物的導管分子處于相對進化但較為初級的階段。即已經普遍存在單穿孔板,孔紋導管等進化的標志,但仍存留如螺紋導管、梯狀穿孔板等相對原始的結構。并且在相對進化的桃亞屬中仍存在螺紋導管。但由于東北地區本地產的桃亞屬植物相對較少,無法進行亞屬內的比較,有待進一步研究。在之前的研究中,對導管分子尾端的研究基本停留在長度的相對性上:即尾端相對較長的導管分子較為原始,尾端較短的相對進化[13~14,22]。本研究則對李屬植物導管分子的尾端長度進行了測量,認為李屬植物導管分子尾端長度在24~36 μm,屬于相對短的尾端長度。并且發現在不同生境下同種植物尾端差異不顯著,長度較為穩定。這說明尾端長度可能僅與物種相關,與生境無關。由于櫻亞屬在李屬中相對原始,但觀察結果中櫻亞屬的毛櫻桃尾端最長、同亞屬的歐李、郁李卻最短。推測櫻亞屬植物的導管分子不同特征可能在進化程度上不一致。但由于李屬植物導管分子尾端長度整體相對穩定,因此本研究也支持將尾端長度作為衡量植物系統學地位佐證。但這也需要進一步測量薔薇科其他各屬植物后再進行比較分析。并且我們在觀察中發現,導管分子尾端也有功能性結構(螺紋加厚與紋孔對),這也推測李屬植物的尾端并不是無功能的盲端結構,而是可以參與橫向水分運輸的功能性結構。在環境適應性方面,導管為植物各個器官運輸水分與無機鹽,而植物導管其輸水能力和導管分子的形態結構是密切相關的。端壁斜度角度大小直接影響管腔端壁截面的面積,端壁截面面積越大,運輸水分阻力越大,輸水效率越低。導管分子越長,單位長度內端壁數量越少,在增加管腔壓力(即提高根系吸水能力)的同時,減少了端壁對水的阻力。導管直徑越大,輸水量越大,但越容易出現栓塞與空穴化[23~24]。但環境對長度與寬度的影響則有很多理解:Steven J.發現由寒帶到亞熱帶植物導管分子的紋孔發生率明顯上升。V.E.Ben?kova 在對西伯利亞的薔薇科植物的木材解剖研究中發現,生長在亞北極地帶的植物擁有較高的導管頻率與較細的導管分子。劉樹煥等則認為導管分子較長,端壁面積較小是導管分子適應高海拔等干旱條件的結果[22,25~26]。從導管分子的長度、寬度和端壁斜度角度數據分析結果顯示,李屬植物在高緯度高海拔地區為了適應生境提高了導管長度(即減少了單位長度穿孔板數量),減小了管腔直徑(即降低了栓塞的發生率),從而提高了輸水能力(即提高了水分利用率)。生長在吉林與遼寧的東北李,由于海拔與緯度變化幅度不大,所以在導管長度、寬度等形態上并沒有顯著差異,這也推測生境的緯度與海拔變化相對大時,導管分子形態會出現變化。但對比生長在黑龍江與遼寧的李時發現:在緯度相對較高的地區,到緯度高于一定程度時小幅度的緯度增加導管分子結構也可能出現差異。東北地區除長白山外,海拔均低于700 m,屬于地平地帶。比較適合同種植物在不同緯度下的生境比較。由于長白山均海拔1 500 m 以上,屬于亞高山帶。所以生長在長白山亞高山帶的榆葉梅、歐李、郁李和山杏與生境在更高緯度的同種植物相比較,其導管長度相同甚至更長,寬度幾乎相同,端壁面積相同甚至更小。因此,我們認為在高緯度、高海拔的生境下,植物為了更適應生境,導管分子形態會根據生境做出反饋:為提高輸水效率,增加了導管長度進而減少了穿孔板數量。為降低空穴化發生率,降低了導管寬度,從而減少了導管栓塞的發生率。但成熟導管分子的寬度似乎存在最小極限,以至于高海拔情況下寬度并沒有更大幅度的變小,但這仍需要進一步研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
