調控微小RNA-17-92表達對血吸蟲病大鼠肝纖維化的抑制作用▲
2021-02-26 08:57:30郭忠欣李蘭花李雪飛呂亞輝
廣西醫學 2021年24期
郭忠欣 李蘭花 康 慨 李雪飛 呂亞輝 朱 偉
(1 北京師范大學醫院公共衛生科,北京市 100875,電子郵箱:gcm580@163.com;2 濰坊醫學院公共衛生學院流行病教研室,山東省濰坊市 261053)
血吸蟲病是一種臨床較為常見的慢性感染性寄生蟲疾病,在數十個非洲、亞洲、拉丁美洲國家流行,而在我國血吸蟲病主要在長江流域及其以南地區流行[1-2],該病可對患者健康、生活質量造成嚴重威脅[3-4]。血吸蟲病的主要臨床表現為發熱、咳嗽、胸痛、血痰、腹瀉、肝區疼痛、腹痛、侵入部位皮炎等,疾病發展還可導致消化道出血、腹腔積液、肝纖維化等并發癥的發生[5-7]。臨床上治療血吸蟲病的常用手段為藥物治療,但是尚無特效藥物用于治療血吸蟲病所致的肝纖維化,因此尋找治療抑制血吸蟲病肝纖維化的新靶點成為目前臨床醫學的研究熱點之一。本研究建立血吸蟲病大鼠模型,并靶向調控微小RNA(mircoRNA,miRNA)-17-92表達,旨在探討調控miRNA-17-92表達對血吸蟲病大鼠肝纖維化的干預作用及相關機制。
1 材料與方法
1.1 實驗動物 選取40只SD健康雄性大鼠,由華蘭生物工程股份有限公司提供,動物許可證號:SYXK(豫)2018-0014。鼠齡8~11(9.5±1.2)個月,體重230~247(238.5±6.8)g。在相對濕度50%~55%、溫度(23.1±1.9)℃的環境中喂養1周,光照12 h/d。
1.2 主要試劑 Lipofectamine 2000試劑購自滁州仕諾達生物科技有限公司(批號:SND464);miRNA-17-92模擬物購自上海易匯生物科技有限公司(批號:CL-0005),miRNA-17-92抑制物購自北京達科為生物技術有限公司(批號:MA-0102);兔抗小鼠透明質酸抗體、兔抗小鼠Ⅲ型前膠原(typeⅢ procollagen,PCⅢ)抗體購自上海廣銳生物科技有限公司(批號:2013-11213、2013-1397),兔抗小鼠層粘連蛋白抗體購自上海瓦蘭生物科技有限公司(批號:E03551);大鼠抗小鼠白細胞介素(interleukin,IL)-2抗體、大鼠抗小鼠IL-22抗體購自上海科敏生物科技有限公司(批號:bs-4586R、bs-2623R);兔抗小鼠活性氧簇抗體、兔抗小鼠8-羥基脫氧鳥苷(8-hydroxy-2′-deoxyguanosine,8-OHdG)抗體購自上海梵態生物科技有限公司(批號:FT-T21239P、FT-P36089R);RNA提取試劑盒、反轉錄試劑盒、miRNA-17-92 RT-PCR試劑盒均購自QIAGEN公司(貨號:217184、218161、218193);小鼠抗兔miRNA-17-92、轉化生長因子β受體Ⅲ(transforming growth factor β receptor type Ⅲ,TGFβRⅢ)抗體購自亞科因(武漢)生物技術有限公司(批號:A25022-1、ABP57480);大鼠抗小鼠Smad2、Smad3抗體購自武漢菲恩生物科技有限公司(批號:FN-ER0645、FNab07994);大鼠抗小鼠磷脂酰肌醇-3-激酶(phosphatidylinositol-3-kinase,PI3K)、蛋白激酶B(protein kinase B,Akt)、哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)抗體購自武漢菲恩生物科技有限公司(批號:FN-ER1910、ER1857、ER1520);山羊抗兔IgG購自上海晶風生物科技有限公司(批號:TF-0295G-SA)。
1.3 建模、分組和干預方法 采用隨機數字表法選取10只大鼠作為正常組,不做處理。其余30只大鼠建立血吸蟲病模型:將大鼠腹部小塊區域被毛去除,采用生理鹽水濕潤裸露皮膚,顯微鏡下計數60條血吸蟲尾蚴(購自江蘇省血吸蟲病防治中心),貼蓋在大鼠裸露皮膚處,持續10 min,以皮膚可見多處點狀充血為建模成功。20 d后將30只血吸蟲病大鼠模型隨機分為血吸蟲病組、上調組、下調組各10只。利用Lipofectamine 2000試劑將miRNA-17-92模擬物、miRNA-17-92抑制物稀釋為5 mmol/L的濃度,然后通過尾部靜脈分別注射入上調組、下調組大鼠體內,正常組、血吸蟲病組大鼠經尾部靜脈注射等量生理鹽水,各組的注射劑量均為1 mL/次,間隔12 h注射一次,共2次。注射結束后24 h檢測相關指標。
1.4 指標檢測
1.4.1 獲取標本:對各組大鼠進行麻醉(25%烏拉坦,0.5 mL/100 g,腹腔注射)處理后,頸椎脫臼法處死,取各組大鼠腹部靜脈血3 mL,室溫下靜置2 h,4℃下2 000 r/min離心15 min后分離上清液,在-80℃環境中保存待檢,并留取肝臟組織用于后續實驗。
1.4.2 觀察肝臟組織病理學:取大鼠肝臟組織固定在4%甲醛中,完全浸泡,于24 h后行常規石蠟包埋及連續切片。將切片烤干后進行脫蠟處理,之后順序置入不同濃度乙醇中各水化3 min。使用蘇木精染色15 min后采用TBST清洗3次(5 min/次),使用鹽酸乙醇溶液分化處理30 s,充分清洗后使用1%伊紅染色,使用不同濃度乙醇進行脫水處理后進行脫蠟處理,封片后使用顯微鏡(深視光谷儀器有限公司,型號:SGO-KK209)進行觀察。
1.4.3 檢測血清中肝功能、肝纖維化指標水平:使用全自動生化分析儀(深圳市新產業生物醫學工程股份有限責任公司,型號:VDVIA3500)檢測血清中肝功能指標,γ-谷氨酰轉移酶(gamma-glutamyltransferase,GGT)、ALT、AST水平;采用放射免疫分析法,使用相應試劑檢測血清中肝纖維化指標,透明質酸、PCⅢ、層粘連蛋白水平。嚴格按照試劑說明書步驟進行操作。
1.4.4 檢測血清中氧化應激、炎癥反應指標水平:將血清標本、稀釋液以及檢測卡平衡至24℃,對檢測卡進行編號后置于平臺之上,配制標準液并按照1 ∶10比例對待測標本進行稀釋。酶標板中添加100 μL血清標本或標準液,混合均勻后在恒溫箱(37℃)中靜置0.5 h,采用TBST緩沖液洗板3次(5 min/次),每孔中添加50 μL一抗以及50 μL蒸餾水,混勻后37℃環境中靜置0.5 h。采用TBST緩沖液洗板3次(5 min/次)后每孔中添加100 μL酶標抗體,37℃環境中靜置10 min,采用TBST緩沖液洗板3次(5 min/次)后每孔中添加100 μL底物液,37℃、陰暗環境中靜置15 min后每孔中添加100 μL終止液混勻,使用酶標儀(賽默飛公司,型號:Varioskan LUX)檢測波長為450 nm處的吸光值。
1.4.5 實時PCR法檢測肝組織miRNA-17-92的相對表達水平:提取肝臟組織總RNA,檢測RNA純度、含量,反轉錄處理后獲得cDNA。使用Primer5.0軟件設計引物。miRNA-17-92上游引物序列為5′-GTGCAGGGTCCGAGGT-3′,下游引物序列為5′-GTGTTCAAAGTCTTACCGTGC-3′;內參U6上游引物序列為5′-GGGGACATCCGATAAAATTGG-3′,下游引物序列為5′-GATTTGTGCGTGTCATCATCCTTG-3′。PCR儀購自杭州朗基科學儀器有限公司(型號:T30D)。配置反應體系,包括0.4 μL上下游引物、1 μL cDNA模板、10 μL SYBR Green,加蒸餾水至20 μL。反應條件為97℃預熱8 min、95℃變性5 s、60℃退火31 s,共進行40個循環。采用2-△△Ct方法計算出miRNA-17-92的相對表達量。
1.4.6 蛋白免疫印跡法檢測TGF-β/Smads通路、PI3K/Akt通路的蛋白相對表達水平:將各組大鼠肝臟組織剪碎后加入裂解液裂解30 min,然后3 000 r/min離心處理10 min(離心半徑3 cm),取上清液,提取50 μg蛋白,煮沸5 min使蛋白變性后進行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳,然后依據蛋白分子大小把凝膠切開并轉移至聚偏二氟乙烯膜上,采用Western封閉液室溫封閉90 min;按比例稀釋一抗(待測蛋白抗體按1 ∶1 000,內參抗體按1 ∶2 000)后4℃孵育過夜,使用Western洗滌液洗滌5次,10 min /次;加入稀釋山羊抗兔IgG(1 ∶5 000),室溫水平搖床孵育60 min,Western洗滌液洗滌5次,10 min /次。經過增強型化學發光試劑顯色和曝光后,使用Image J圖像分析系統檢測條帶的灰度值,以待測蛋白與內參條帶灰度值的比值作為目的蛋白的相對表達量。
1.5 統計學分析 使用SPSS 26.0軟件進行統計學分析。計量資料以(x±s)表示,多組比較采用方差分析,進一步兩兩比較采用LSD-t檢驗。以P<0.05為差異有統計學意義。
2 結 果
2.1 4組大鼠的肝組織病理學變化 正常組大鼠肝組織細胞排列整齊且緊密,結構較為清晰,大小均一;血吸蟲病組大鼠肝組織細胞排列雜亂且疏松,出現部分細胞壞死、結構損壞、纖維結締組織增生、炎性細胞浸潤狀況;下調組大鼠肝組織細胞排列雜亂且疏松,出現明顯細胞壞死、結構損壞、纖維結締組織增生、炎性細胞浸潤狀況;上調組大鼠肝組織細胞排列較為整齊,結構較為清晰,細胞壞死、結構損壞、纖維結締組織增生、炎性細胞浸潤狀況均較血吸蟲病組明顯減輕。見圖1。
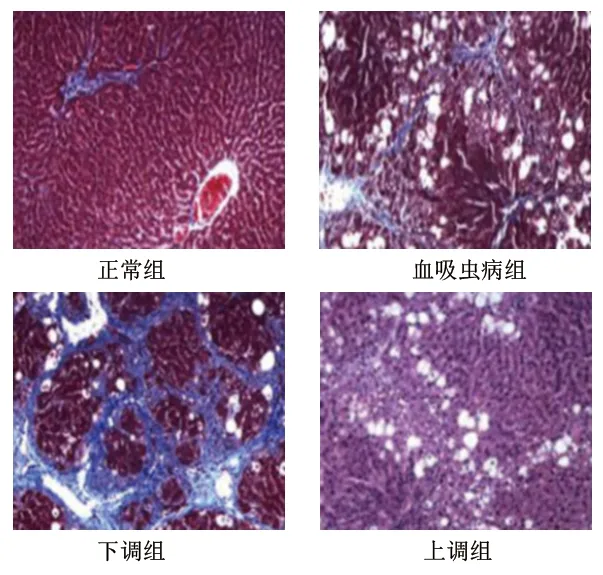
圖1 4組大鼠的肝組織病理學變化(蘇木精-伊紅染色,×100)
2.2 4組大鼠肝功能指標水平的比較 與正常組比較,血吸蟲病組、下調組、上調組大鼠的血清GGT、ALT、AST水平均升高;與血吸蟲病組比較,下調組大鼠的血清GGT、ALT、AST水平均上升,上調組大鼠的血清GGT、ALT、AST水平均降低;與下調組相比,上調組大鼠的血清GGT、ALT、AST水平均降低(均P<0.05)。見表1。
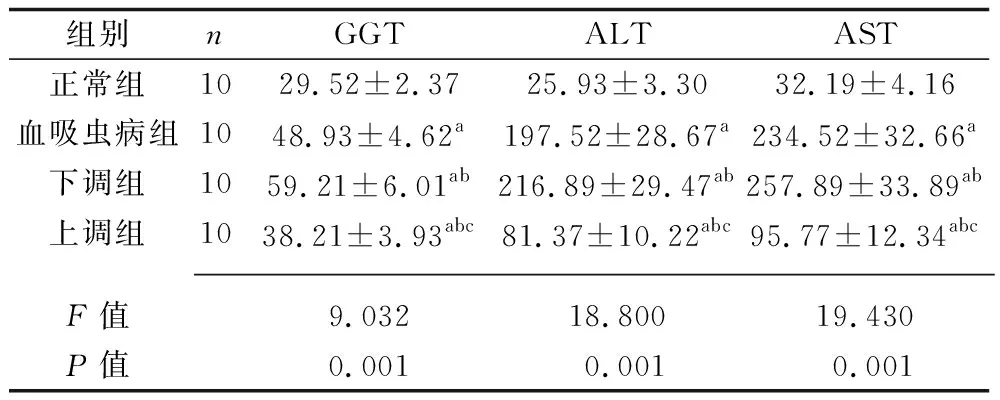
表1 4組大鼠肝功能指標水平的比較(x±s,U/L)
2.3 4組大鼠肝纖維化指標的比較 與正常組比較,血吸蟲病組、下調組、上調組大鼠的血清透明質酸、PCⅢ、層粘連蛋白水平均升高;與血吸蟲病組比較,下調組的血清透明質酸、PCⅢ、層粘連蛋白水平升高,上調組大鼠的血清透明質酸、PCⅢ、層粘連蛋白水平均降低;與下調組大鼠比較,上調組大鼠的血清透明質酸、PCⅢ、層粘連蛋白水平均降低(均P<0.05)。見表2。
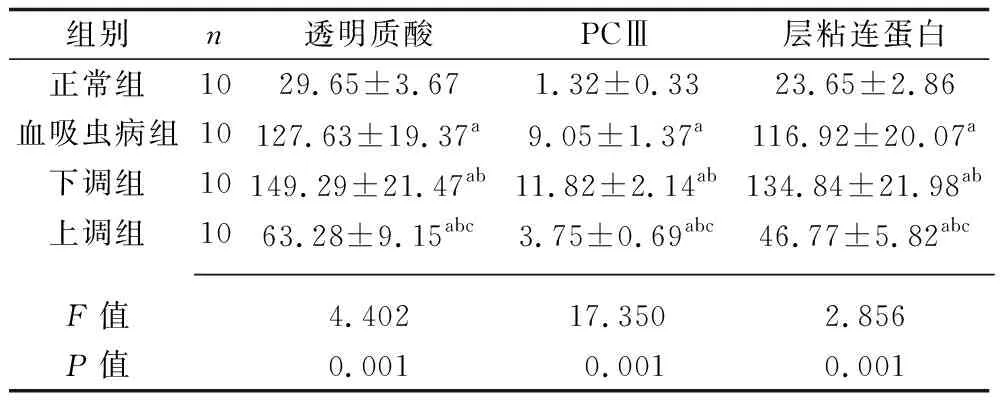
表2 4組大鼠肝纖維化指標水平的比較(x±s,μg/L)
2.4 4組大鼠氧化應激和炎癥指標的比較 與正常組比較,血吸蟲病組、下調組、上調組大鼠的血清活性氧簇、8-OHdG、IL-2、IL-22水平均升高;與血吸蟲病組比較,下調組大鼠的血清活性氧簇、8-OHdG、IL-2、IL-22水平升高,上調組大鼠的血清活性氧簇、8-OHdG、IL-2、IL-22水平均降低;與下調組大鼠比較,上調組大鼠的血清活性氧簇、8-OHdG、IL-2、IL-22水平均降低(均P<0.05)。見表3。
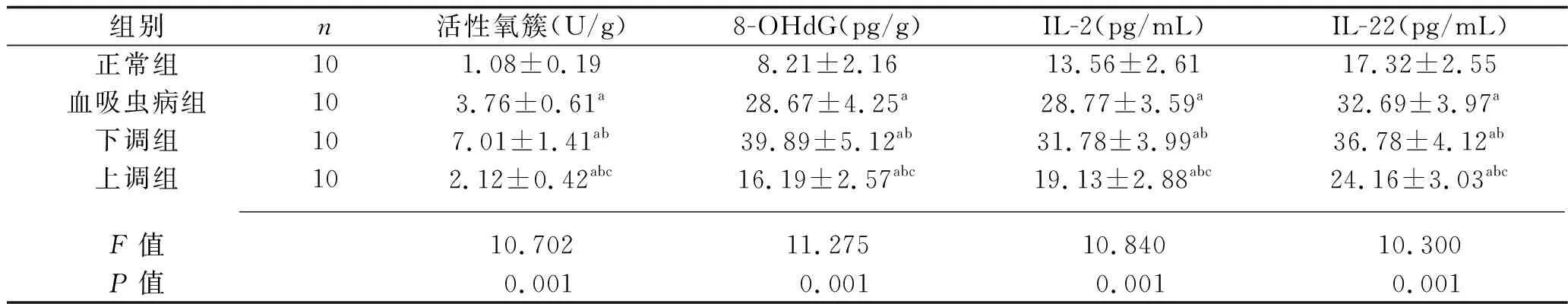
表3 4組大鼠氧化應激和炎癥指標的比較(x±s)
2.5 4組大鼠的miRNA-17-92及TGF-β/Smad通路蛋白相對表達量的比較 與正常組比較,血吸蟲病組、下調組、上調組大鼠miRNA-17-92的相對表達量較低,TGFβRⅢ、Smad2、Smad3蛋白的相對表達量較高;與血吸蟲病組比較,下調組大鼠miRNA-17-92的相對表達量較低,TGFβRⅢ、Smad2、Smad3蛋白的相對表達量較高,上調組大鼠miRNA-17-92的相對表達量較高,TGFβRⅢ、Smad2、Smad3蛋白的相對表達量較低;與下調組大鼠比較,上調組大鼠miRNA-17-92的相對表達量較高,TGFβRⅢ、Smad2、Smad3蛋白的相對表達量較低(均P<0.05)。見表4。
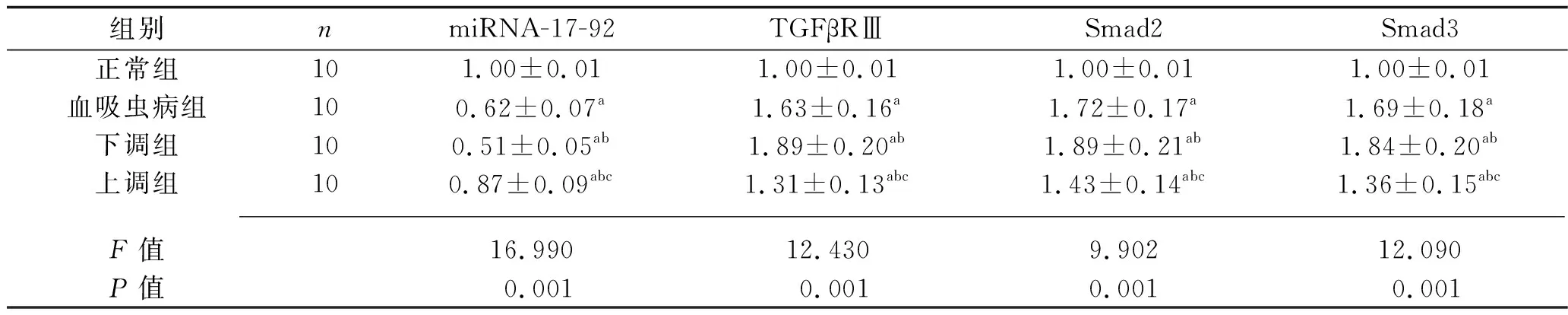
表4 4組大鼠miRNA-17-92及TGF-β/Smad通路蛋白相對表達量的比較(x±s)
2.6 4組大鼠PI3K/Akt通路蛋白相對表達量的比較 與正常組比較,血吸蟲病組、下調組、上調組大鼠PI3K、Akt、mTOR蛋白的相對表達量較高;與血吸蟲病組比較,下調組PI3K、Akt、mTOR蛋白的相對表達量較高,上調組大鼠PI3K、Akt、mTOR蛋白的相對表達量較低;與下調組比較,上調組大鼠PI3K、Akt、mTOR蛋白的相對表達量較低(均P<0.05)。見表5。
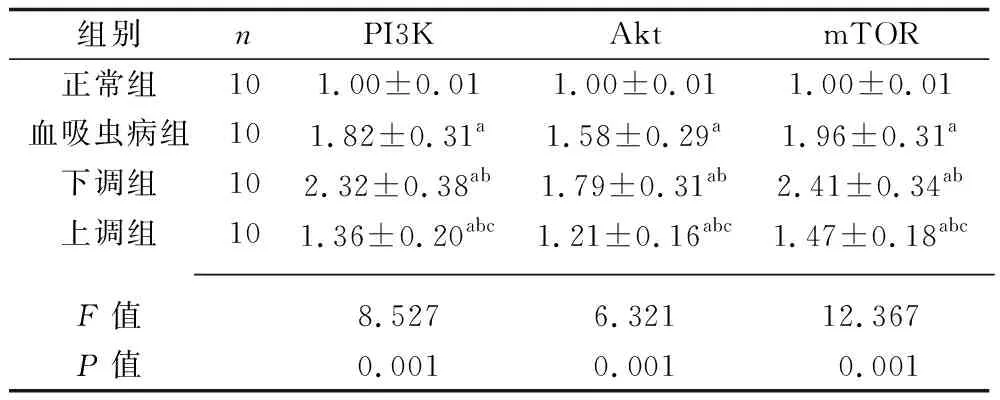
表5 4組大鼠PI3K/Akt通路蛋白相對表達量的比較(x±s)
3 討 論
作為一種慢性感染性免疫系統疾病,血吸蟲病廣泛分布于全球數十個國家和地區[8-9]。有研究顯示,隨著血吸蟲病的發展,沉積于機體肝臟組織中的蟲卵會導致肝損傷、肝纖維化,可嚴重影響患者的預后,因此尋找一種安全有效的治療手段具有重要意義[10-11]。目前臨床上尚無一種特效的治療血吸蟲病所致肝纖維化的藥物,越來越多的專家學者開始致力于這方面的研究。
血吸蟲病的發生、發展可引起肝功能損傷甚至肝硬化等,減輕肝功能損傷的嚴重程度對血吸蟲病治療、預后改善具有重要意義[12]。miRNA廣泛存在于大多數生物中,具有組織特異性、穩定性、保守性等特點,與機體組織細胞增殖、遷移、凋亡等生物學行為密切相關[13-14]。miRNA-17-92作為miRNA家族的重要成員,其表達水平的變化與血管疾病、機體正常生長發育、免疫調節密切相關[15]。有研究顯示,miRNA-17-92缺失對肝臟再生具有抑制作用,進而引發了肝功能損傷[16]。但目前鮮有miRNA-17-92在血吸蟲病方面的研究。本研究結果顯示,血吸蟲病大鼠的血清GGT、ALT、AST水平上升,而上調miRNA-17-92表達后血吸蟲病大鼠的血清GGT、ALT、AST水平下降,說明血吸蟲病大鼠出現一定程度的肝功能損傷,而上調miRNA-17-92表達能夠緩解大鼠的肝功能損傷嚴重程度,改善血吸蟲病大鼠的肝功能指標。
肝組織持續的炎癥反應會造成纖維結締組織增生并大量沉積,進而促進肝組織纖維化的發生、發展[17]。有研究顯示,蟲卵可溶性抗原會對血吸蟲患者機體進行持續刺進,進而引發了免疫應答,導致組織損傷、蟲卵肉芽腫等慢性炎癥的發生,最終引發肝纖維化,嚴重者可導致死亡,這對患者的預后造成了嚴重影響[18]。本研究結果顯示,血吸蟲病大鼠的血清透明質酸、PCⅢ、層粘連蛋白水平升高,肝組織細胞排列雜亂且疏松,出現部分細胞壞死、結構損壞、纖維結締組織增生、炎性細胞浸潤狀況;而上調miRNA-17-92表達的血吸蟲病大鼠的血清透明質酸、PCⅢ、層粘連蛋白水平下降,且肝組織細胞排列較為整齊,結構較為清晰,細胞壞死、結構損壞、纖維結締組織增生、炎性細胞浸潤狀況較血吸蟲病組均明顯減輕,這說明血吸蟲病大鼠出現肝纖維化現象,而上調miRNA-17-92表達能夠減輕其肝纖維化程度。研究表明,TGF-β/Smad信號通路在機體組織纖維化發展過程中起到重要作用[19]。本研究結果顯示,上調miRNA-17-92表達的血吸蟲病大鼠TGFβRⅢ、Smad2、Smad3蛋白表達下調,說明上調miRNA-17-92表達能夠靶向調控TGF-β/Smad通路蛋白表達,這可能是其抑制肝纖維化發展、發揮肝臟保護作用的可能機制之一。
肝損傷、肝纖維化的發生和發展伴隨著肝組織的氧化應激反應及炎癥反應[20]。活性氧簇具有一定氧化性,肝損傷發展過程中活性氧簇大量生成[21],致使氧化產物8-OHdG大量生成。IL-2、IL-22是常用的評價機體炎癥反應的指標,二者水平變化與機體炎癥反應密切相關[22-23]。本研究結果顯示,血吸蟲病大鼠的血清活性氧簇、8-OHdG、IL-2、IL-22水平上升,而上調miRNA-17-92表達的血吸蟲病大鼠的血清活性氧簇、8-OHdG、IL-2、IL-22水平下調,這說明上調miRNA-17-92表達能夠有效地減輕血吸蟲病大鼠肝組織氧化應激損傷和炎癥反應,從而減輕肝纖維化。
肝臟組織細胞凋亡是慢性肝損傷、肝纖維化發展過程中的主要特征[24-25]。PI3K/Akt信號通路相關蛋白P13K、Akt、mTOR蛋白表達變化與細胞凋亡能力密切相關,靶向調控PI3K/Akt信號通路蛋白表達,能夠干預細胞增殖、凋亡[26]。本研究結果顯示,上調miRNA-17-92表達的血吸蟲病大鼠PI3K、Akt、mTOR的蛋白相對表達水平下調,因此我們推測上調miRNA-17-92表達可能通過靶向調控PI3K/Akt通路蛋白表達,抑制肝臟組織細胞凋亡,從而起到抑制肝纖維化發展的作用。
綜上所述,上調miRNA-17-92表達能夠改善血吸蟲病大鼠模型的肝功能和肝纖維化程度,其作用機制可能是調控miRNA-17-92能夠減輕肝組織氧化應激反應和炎癥反應,并靶向調控TGF-β/Smad通路、PI3K/Akt通路蛋白從而抑制肝纖維化及肝組織細胞凋亡,這或許可為血吸蟲病所致肝纖維化的臨床治療提供一定的理論依據。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年12期)2019-05-21 02:55:32
學苑創造·A版(2015年11期)2016-01-14 09:03:27
現代檢驗醫學雜志(2014年4期)2014-02-02 02:44:59
