不同倍性芍藥雜交親和性及子代倍性1)
2021-02-10 11:17:04朱煒陳啟航董志君孫慧儀孟子卓陳曦陶婧婷于曉南
東北林業大學學報 2021年12期
朱煒 陳啟航 董志君 孫慧儀 孟子卓 陳曦 陶婧婷 于曉南
(北京林業大學,北京,100083)
中國是野生芍藥的分布中心,共有7種2亞種,約占芍藥組(Sect.Paeonia)植物的1/3[1]。近年來,我國研究者對芍藥組植物的重視程度逐漸增加,但目前國內對于芍藥組植物的新品種選育工作還不足,豐富的種質資源未得到充分的利用[2]。截至2020年10月,芍藥屬植物國際登錄權威美國芍藥牡丹協會共登錄了芍藥品種5 200多個,其中中國登錄的芍藥品種僅25個,說明我們國家的芍藥品種的選育水平與歐美國家還存在較大的差距。
定向雜交,尤其是遠緣雜交,是園林植物種質創新的重要途徑[3],亦是芍藥屬植物育種的最主要方式[2]。芍藥組野生種存在二倍體和四倍體的倍性分化,經過雜交選育獲得的芍藥品種倍性豐富,存在二倍體、三倍體和四倍體[4-5]。多倍體植株通常具有莖稈粗壯、花色艷麗、花瓣增多、葉片肥厚、抗性強等特點。不同倍性植物雜交是多倍體和非整倍體種質創造的一種重要手段。然而,不同倍性植物雜交存在一定的親和性障礙。目前,在蘭花[6-7]、萱草[8-9]、紫薇[10]、石竹[11-12]、百合[13-14]等觀賞植物中對不同倍性雜交親和性及子代倍性分離已有一些研究報道。芍藥屬植物雜交親和性研究主要集中在同倍性的牡丹亞組間雜交[15-16]、組間雜交[17-18]和芍藥組內雜交[19-20],對于不同倍性芍藥品種的雜交親和性及后代倍性的分離規律鮮見報道[21]。
本研究以16個芍藥品種為親本,設計不同倍性組合的雜交試驗,研究不同倍性芍藥品種間雜交的親和性,并對部分子代的染色體數目進行鑒定,以期為芍藥定向育種、多倍體種質創新提供理論依據。
1 材料與方法
選取16個芍藥品種(表1),統一種植在國家花卉工程技術研究中心北京市小湯山基地,常規田間管理。選擇生長勢基本一致、健壯、無病蟲害植株作為試驗材料。親本材料的倍性信息參照前人研究結果[4,18,22-23]。
1.1 雜交試驗
根據品種的生物學特性,共設計不同倍性雜交組合42組。雜交試驗于2017年4月底至5月底進行,雜交授粉方法參照劉建鑫等[18]。
1.2 雜交親和性
待芍藥果莢變為蟹黃色時采收,獲得種子,計算結實率。通過水選法篩選得到有胚種子,計算有胚率。對有胚種子沙藏處理,待胚根突破種皮時大田播種,翌年5月計算出苗率。最終,計算雜交組合的親和性指數,以此作為雜交親和性的指標。其中:結實率=(結實朵數/授粉朵數)×100%,有胚率=(有胚種子數/種子總數)×100%,出苗率=(出苗數/有胚種子數)×100%,單果種子數=種子總數/授粉朵數,親和性指數=單果種子數×有胚率×出苗率。
1.3 雜交后代倍性鑒定
采用根尖壓片法對獲得的部分雜交后代進行倍性鑒定。倍性鑒定材料選取和處理參照朱煒等[23]的方法。染色體條數鑒定參照李懋學等[24]的標準。
1.4 數據處理
采用WPS Office軟件對數據進行統計分析,采用Adobe Photoshop CS6對圖片進行分析。
2 結果與分析
2.1 以二倍體為母本的雜交
以二倍體芍藥品種為母本,共設計雜交組合15組,雜交739朵花,獲得后代212株(表2)。從表2中可以看出,同一類倍性雜交組合中不同親本組合結實率、單果種子數、有胚率及出苗率均存在較大差異。在二倍體×二倍體中,結實率和單果種子數最高的組合均為203,分別為90.00%和18.1;有胚率和出苗率最高的組合為202,分別為99.31%和38.89%。在二倍體×四倍體中,結實率和單果種子數最高的組合均為215,分別為97.50%和10.6;有胚率最高的組合為212,有胚率為29.74%;出苗率最高的組合為209,出苗率為66.67%。二倍體品種LT作為母本,與二倍體、四倍體品種雜交,結實率均最高,是較好的育種母本材料。
2.2 以三倍體為母本的雜交
以三倍體品種為母本,共設計雜交組合12組,雜交433朵花,獲得后代3株(表3)。從表3可以看出,以三倍體品種為母本,結實率、單果種子數均普遍較低,即使獲得種子,大部分組合的有胚率或出苗率也為0,最終僅獲得3株后代。其中,三倍體×二倍體中結實率最高的組合為304,結實率20.00%,單果種子數為0.4;三倍體×四倍體中結實率最高的組合為305,結實率為38.81%,單果種子數為0.6。組合304和305的母本均為CH,說明三倍體品種CH可以作為母本獲得種子并得到后代,是一個創造新種質的好材料。
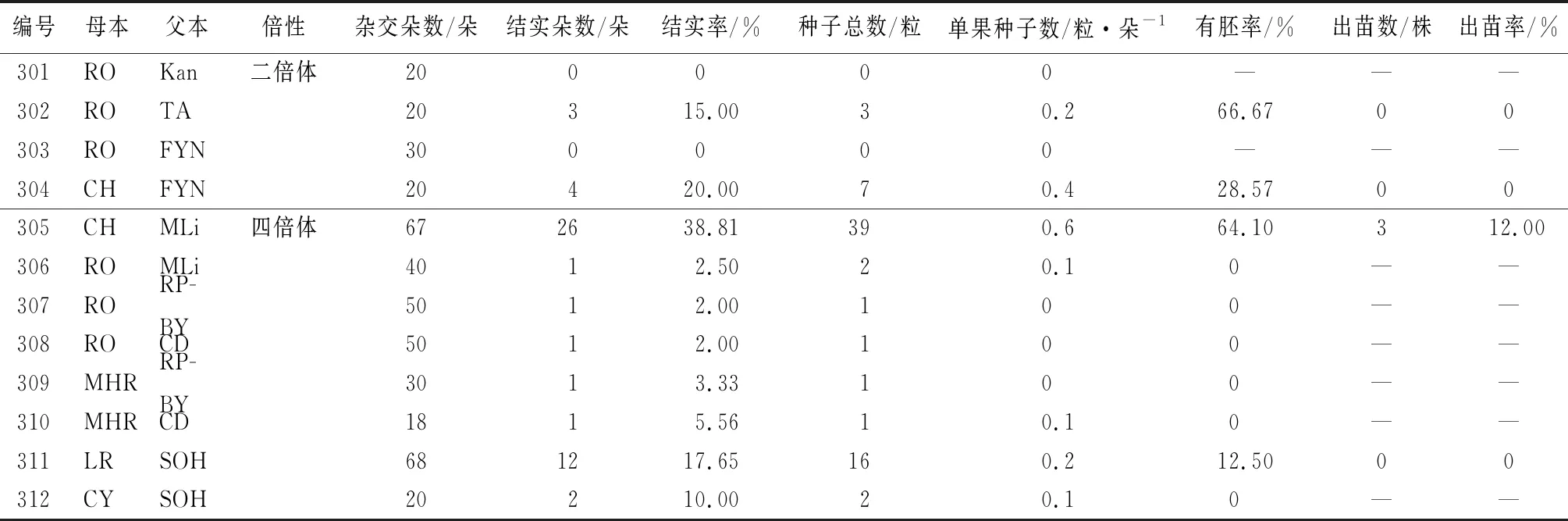
表3 以三倍體芍藥為母本的雜交結果
2.3 以四倍體為母本的雜交
以四倍體品種為母本,共設計雜交組合17組,獲得雜交苗共173株(表4)。從表4可以看出,同一類倍性雜交組合中不同親本組合結實率、單果種子數、有胚率和出苗率均存在較大差異。四倍體×二倍體中組合402的結實率、單果種子數、有胚率和出苗率均為最高,分別為95.00%、4.9、18.37%和5.56%;組合405和406結實率最低,均為0。四倍體×四倍體中組合410結實率和單果種子數均最高,分別為100.00%和7.3;組合413有胚率最高,為100.00%;組合407出苗率最高,為35.28%。四倍體品種SOH、GP作為母本不論與二倍體還是四倍體雜交,結實率均為0,說明在多倍體雜交育種中SOH、GP均不適宜作為母本。
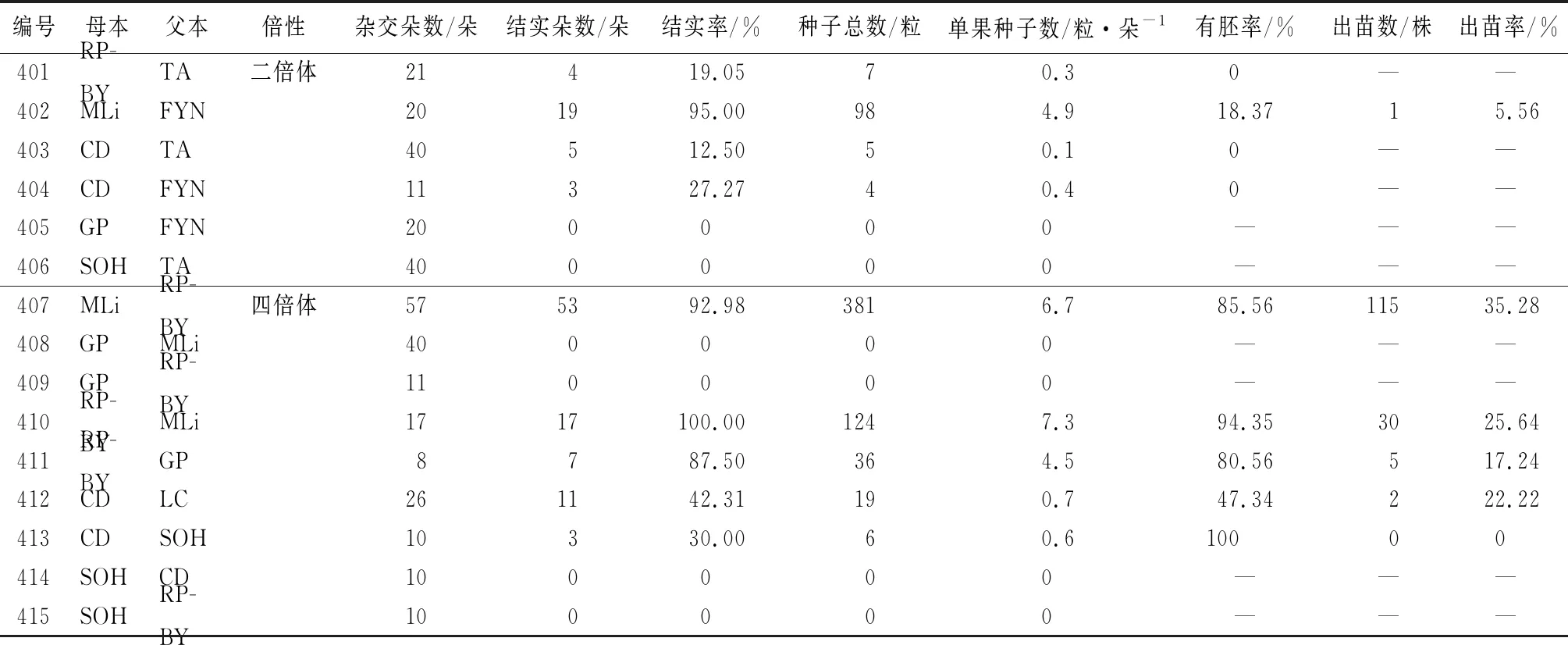
表4 以四倍體芍藥為母本的雜交結果
2.4 不同倍性品種雜交親和性比較
從表5可以發現,在以二倍體為母本的組合中,二倍體×二倍體組合的平均單果種子數、平均有胚率和平均出苗率均顯著高于二倍體×四倍體組合,其中平均有胚率的差異最為顯著。在以三倍體為母本的組合中,三倍體×二倍體組合的平均單果種子數與三倍體×四倍體組合接近,平均有胚率顯著高于后者,但前者的出苗率為零,未獲得后代,說明在育種中以三倍體為母本時,應該選擇四倍體作為父本。在以四倍體為母本時,四倍體×四倍體組合的平均單果種子數、平均有胚率和平均出苗率均顯著高于四倍體×二倍體組合。綜合3項指標,計算各類雜交組合親和性指數,6類雜交組合的親和性由大到小為:二倍體×二倍體、四倍體×四倍體、二倍體×四倍體、四倍體×二倍體、三倍體×四倍體、三倍體×二倍體。
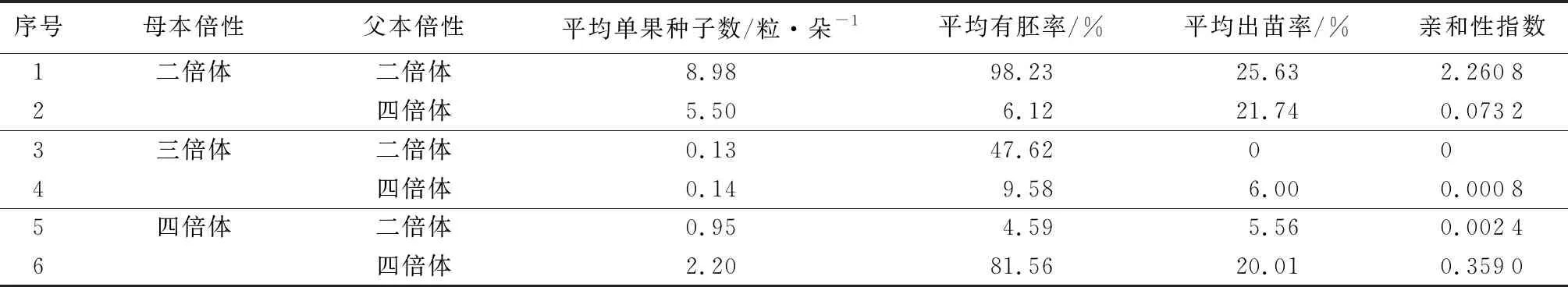
表5 不同父本、母本倍性品種雜交后親和性
2.5 雜交后代染色體倍性鑒定
通過根尖壓片法鑒定獲得的部分子代倍性(表6)。二倍體×二倍體的后代染色體數均為10條,為二倍體(圖1a);二倍體×四倍體的后代染色體數目表現多樣,有10,15和20條3種情況,共鑒定出30株二倍體(圖1b),11株三倍體(圖1cd),3株四倍體(圖1e);三倍體×四倍體的后代均為四倍體(圖1f),未檢測到非整倍體;四倍體×二倍體的唯一的1株后代鑒定為非整倍體(圖1g),染色體數目為2n=3x+1=16;四倍體×四倍體的后代均為四倍體(圖1h),未發現其他倍性結果。
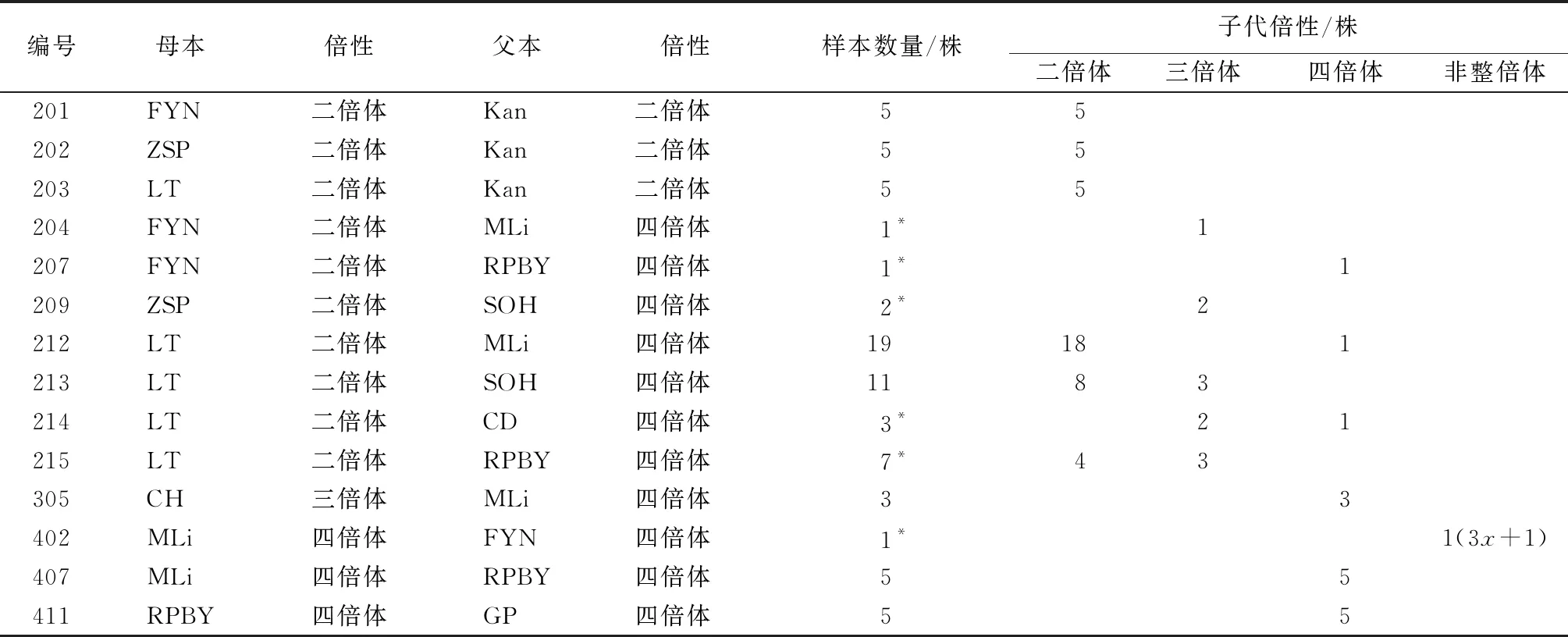
表6 部分子代倍性
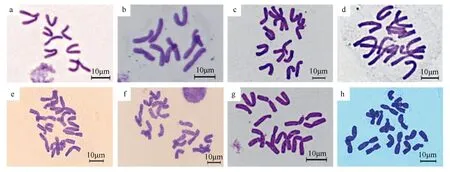
a.組合203子代2n=2x=10;b.組合213子代2n=2x=10;c、d.組合213子代2n=3x=15;e.組合214子代2n=4x=20;f.組合305子代2n=4x=20;g.組合402子代2n=3x+1=16;h.組合407子代2n=4x=20。
3 結論與討論
染色體倍性對植物雜交親和性具有直接的影響[25]。本研究中,不同倍性芍藥親和性由大到小為:二倍體×二倍體、四倍體×四倍體、二倍體×四倍體、四倍體×二倍體、三倍體×四倍體、三倍體×二倍體。相同倍性品種間雜交具有較高的親和性,其中二倍體×二倍體的親和性高于四倍體×四倍體,這可能是四倍體大孢子母細胞減數分裂過程中染色體更容易發生異常行為所致。二倍體×四倍體單果種子數要高于四倍體×四倍體,但是二倍體×四倍體的有胚率非常低,所以二倍體×四倍體的雜交親和性要低于四倍體×四倍體。不同倍性雜交親和性研究在其他物種中早有報道,但因材料的不同雜交親和性結果也存在一些差異。本研究中二倍體×四倍體的單果種子數、有胚率和出苗率均顯著高于四倍體×二倍體,說明在培育三倍體芍藥時應該以二倍體做母本,這與郭印山等[26]對2n和4n葡萄雜交組合的親和性研究的結果一致。王滑等[10]以紫薇為材料研究不同倍性雜交的親和性,結果顯示二倍體與四倍體的雜交中,四倍體作為母本的親和性更高,與本研究結果不一致。在以三倍體為母本的雜交試驗中,三倍體×四倍體親和性高于三倍體×二倍體,與袁國良[27]以三倍體LAA百合做母本,與二倍體和四倍體亞洲百合雜交結果一致。
Johnston et al.[28]提出的胚乳平衡數(EBN)假說認為在雜交中當胚乳母本與父本基因組比率偏離2∶1時,胚乳將發育異常,導致種子敗育。本研究二倍體×四倍體和四倍體×二倍體組合雜交種子中,胚乳的母本與父本基因組比值分別為1∶1和4∶1,故種子敗育嚴重。而四倍體×二倍體親和性要低于二倍體×四倍體,可能是由于芍藥四倍體品種為異源多倍體,形成的胚珠數量有限且育性較低,最終導致二倍體×四倍體獲得的種子和后代植株比四倍體×二倍體要多。
Stebbins[29]基于形態性狀的比較和減數分裂的觀察,認為芍藥組內大多數四倍體類群可能是異源四倍體。而雜種芍藥品種群中的多倍體均由兩個及以上種源組成,故屬于異源多倍體品種。前人研究表明,三倍體可產生可育的雌配子,作為母本與合適的父本雜交獲得后代[30-31]。本研究中篩選出一個作為母本育性較好的三倍體品種‘Chalice’,其育性機理有待進一步研究。四倍體植株是獲得多倍體種質的重要親本材料。本研究發現4個四倍體品種‘Old Faithful’、‘May Lilac’、‘Cream Delight’和‘Roy Pehrson Best Yellow’不論作為父本或母本都具有較好的育性,是芍藥多倍體育種的重要種質資源。研究還發現2個四倍體品種‘Garden Peace’和‘Scarlet O’Hara’僅適合作為父本,作為母本不結實,可能是無法形成正常的雌配子,其雌配子敗育的原因有待進一步研究。
本研究中,相同倍性的雜交組合二倍體×二倍體和四倍體×四倍體后代染色體均保持與父母本相同的倍性,說明在芍藥中染色體數目遺傳相對較穩定,這與周建金等[6]對不同倍性蝴蝶蘭雜交后代的染色體倍性鑒定結果不一致。本研究二倍體×四倍體組合產生了二倍體、三倍體和四倍體,與周建金等[6]研究結果一致,二倍體的產生可能是四倍體父本減數分裂異常產生了具有活力的雄配子,四倍體的形成可能是二倍體母本產生了未減數雌配子。袁國良[27]對百合三倍體×四倍體的雜交子代進行鑒定,發現均為非整倍體。而本研究中三倍體×四倍體產生的后代均為四倍體,未發現非整倍體,可能是三倍體母本產生的有活力的二倍體雌配子更容易與四倍體產生的二倍體雄配子結合。芍藥屬植物染色體屬于大型染色體,數目較少,基數x=5,這說明其單條染色體控制著眾多的遺傳信息,增減一條均可能導致配子死亡,故在芍藥屬植物中尚未見有非整倍體的報道。但在本研究的四倍體×二倍體組合中,對僅獲得的1株苗進行鑒定,結果顯示為非整倍體(2n=3x+1=16),可能是四倍體母本產生了有活力的二倍體+1雌配子。
