外源噴施植物生長調節劑對套作大豆碳氮代謝和花莢脫落的影響
2021-02-05 14:26:12雍太文楊文鈺
作物學報 2021年4期
羅 凱 謝 琛 汪 錦 王 甜 何 舜 雍太文,* 楊文鈺
外源噴施植物生長調節劑對套作大豆碳氮代謝和花莢脫落的影響
羅 凱1謝 琛1汪 錦1王 甜1何 舜2雍太文1,*楊文鈺1
1四川農業大學農學院/ 農業農村部西南作物生理生態與耕作重點實驗室/ 四川省作物帶狀復合種植工程技術研究中心, 四川溫江 611130;2成都市種子管理站/成都市農產品質量安全中心, 四川青羊 610072
玉米–大豆帶狀套作模式下, 玉米蔭蔽會抑制大豆苗期生長、減少花莢數、降低產量, 探究外源植物生長調節劑對大豆開花結莢和產量的調控效應對套作大豆生產具有重要意義。采用單因素隨機區組設計研究套作大豆初花期葉面噴施6-芐基腺嘌呤(6-Benzylaminopurine, 6-BA)、2-N,N-二乙氨基乙基己酸酯(diethyl aminoethyl hexanoate, DTA-6)、烯效唑(S3307)對花莢脫落、葉片碳氮代謝及產量形成的影響。結果表明, 外源調節劑會增強盛莢期和鼓粒期葉片蔗糖合成酶(sucrose synthetase, SS)、蔗糖磷酸合成酶(sucrose phosphate synthase, SPS)及轉化酶(invertase, Inv)的酶活性; 提高始粒期莖、葉、莢果的可溶性糖含量, 促進后期莖、葉中可溶性糖向莢果的轉移。外源調節劑會增加始莢期葉片碳、氮含量, 降低葉片C/N比值; 增加始粒期葉片碳含量, 減少氮含量, 提高葉片C/N比值。外源調節劑會增加大豆開花結莢數, 降低落莢數、落莢率及花莢脫落率, 以DTA-6效果較好, 其處理下2年大豆單株有效莢數和產量顯著高于CK, 較CK分別增加25.4%、41.3%和32.9%、37.6%。套作大豆初花期葉面噴施6-BA、DTA-6、S3307提高葉片SPS、SS和Inv酶活性, 協調大豆各器官碳氮代謝動態平衡, 促進大豆開花結莢、減少大豆落莢, 顯著提高大豆的單株有效莢數與產量, 其中DTA-6的增產效果最好。
玉米–大豆帶狀套作; 花莢脫落; 植物生長調節劑; 碳氮代謝; 產量
在大豆生產中, 環境因素和種植模式影響大豆花莢脫落率, 決定大豆產量構成因素和產量的形成[1-2]; 過高的花莢脫落率降低大豆單株有效莢數和單株粒數, 是限制大豆發揮產量潛力的主要因素[3]。研究表明, 大豆花莢的形成與脫落受植物內源激素信號水平變化和同化物供應有效性的協同調控[4-5]; 碳、氮代謝是植物體內重要的代謝途徑, 調控不同時期的養分供應水平, 決定作物生長發育, 影響大豆花莢形成與脫落[6]。栽培措施的優化和化控技術的應用能促進植株生長、提高作物產量, 是發掘大豆產量潛力的重要途徑[7-9]。
調節劑能影響作物在不同生育時期的生長發育, 協調不同器官間的養分分配狀況, 提高對環境的適應性[10-12]。烯效唑(S3307)是一種能抑制赤霉素生物合成的高效植物生長延緩劑[8]; 閆艷紅等[13]發現, 葉面噴施S3307能通過改善大豆葉片碳氮代謝水平, 增加大豆的單株有效莢數與百粒重。2-N,N-二乙氨基乙基乙酸酯(diethyl aminoethyl hexanoate, DTA-6)作為一種新型的植物生長調節劑, 可有效提高作物的品質和產量, 已廣泛應用于玉米、大豆、花生等作物[14-15]。6-芐基腺嘌呤(6-benzylaminopurine, 6-BA)通過促進細胞分裂素的生物合成調節植物細胞的增殖和分化[16], 具有延緩葉片衰老和保綠等作用[17]。
玉米–大豆帶狀套作模式是我國西南地區的主推模式, 能提高復種指數和土地利用率, 實現對自然資源的充分利用[18]。套作大豆因苗期受玉米蔭蔽影響處于生長劣勢, 營養生長期間干物質積累不足, 營養生長與生殖生長之間的平衡被打破[19]; 養分供應失衡加劇花、莢器官間養分競爭, 促使花敗育與莢脫落, 減少開花結莢數與單株有效莢數, 降低大豆產量[20-22]。為充分挖掘套作大豆的產量潛力, 本研究以玉米–大豆帶狀套作模式為對象, 通過初花期葉面噴施6BA、DTA-6、S3307, 研究其對套作大豆葉片碳氮代謝、花莢脫落和產量形成的影響, 旨在為完善植物生長調節劑在套作大豆中的調控技術應用提供理論支撐。
1 材料與方法
1.1 試驗材料
選用緊湊型玉米品種‘登海605’和耐蔭型大豆品種‘南豆25’為試驗材料, 分別由山東登海種業股份有限公司和四川省南充市農業科學院提供。6-芐基腺嘌呤(6-BA, 含量≥98%)和2-N,N-二乙氨基乙基己酸酯(DTA-6, 含量≥98%)為促進型調節劑, 購自生工生物工程(上海)股份有限公司; 烯效唑(S3307, 5%可濕性粉劑)為延緩型調節劑, 購自四川國光農化股份有限公司。
1.2 試驗設計
試驗于2018年和2019年分別在四川省現代糧食產業仁壽示范基地(30°02'N, 104°15'E)和四川省崇州現代農業研發基地(30°56'N, 103°64'E)進行。采用單因素隨機區組設計, 以清水為對照(CK), 在套作大豆初花期葉面噴施20 mg L-16-BA、60 mg L-1DTA-6、50 mg L-1S3307, 用水量為450 kg hm-2。種植方式采用玉米–大豆寬窄行種植, 玉米窄行行距40 cm, 寬行行距160 cm, 寬行內種植2行大豆, 大豆帶內行距40 cm, 玉米、大豆間距60 cm, 帶寬2 m, 帶長6 m。分別于2018年4月8日和2019年4月15日播種玉米, 株距17 cm, 密度58,500株hm-2。分別于2018年6月18日和2019年6月19日播種大豆, 株距8.5 cm, 密度為117,000株hm-2。每個小區內種植3帶, 小區面積36 m2。大豆底肥施用P2O563 kg hm-2, K2O 52.5 kg hm-2。玉米底肥施N 120 kg hm-2, P2O5105 kg hm-2, K2O 112.5 kg hm-2; 大喇叭口期追施N 120 kg hm-2, 施肥方式為行間開溝施肥。在整個生育期間, 適時除草和防治病蟲。
1.3 測定項目與方法
1.3.1 大豆植株可溶性糖、總碳和總氮含量的測定
于大豆初花期(R1)、始莢期(R3)、始粒期(R5)、成熟期(R8)各小區隨機選取長勢一致植株3株, 按莖、葉、柄、莢分別裝袋, 與105℃下殺青30 min后, 在80℃下烘干至恒重, 粉碎后過100目篩后存放于干燥器中。R8期測定的葉片采用倒置網袋法從初熟期(R7)開始收集。參照硫酸-苯酚定糖法[23]測定R1、R5、R8期大豆葉片可溶性糖含量。使用Elementar vario MICRO cube元素分析儀(Elementar公司, 德國)測定R3、R5期大豆各器官中總碳和總氮含量, 碳氮比即總碳含量/總氮含量[24]。
1.3.2 大豆葉片中糖代謝相關酶活性測定 于大豆盛花期(R2)、盛莢期(R4)、鼓粒期(R6)各小區隨機選擇長勢一致3株大豆, 取其倒三葉中間葉, 清洗干凈去除葉脈, 用液氮處理后, 放在-80℃超低溫中保存。參照Chopra等[25]的方法測定蔗糖合成酶(sucrose synthetase, SS)、蔗糖磷酸合成酶(sucrose phosphate synthase, SPS)、轉化酶(invertase, Inv)活性。
1.3.3 大豆花莢脫落調查 于大豆R1期前, 每小區選取4株長勢一致植株, 在地上鋪設尼龍網,以備準確調查花莢脫落數目。自R1期后, 每7 d記錄1次落花數和落莢數, 在R8期考察單株成莢數。
單株結莢數(個株-1)=單株成莢數+單株落莢數
單株開花數(朵株-1)=單株結莢數+單株落花數
單株落花率(%)=單株落花數/單株開花數×100%
單株落莢率(%)=單株落莢數/單株結莢數×100%
花莢脫落率(%)=(單株落花數+單株落莢數)/單株開花數×100%
1.3.4 產量相關參數測定 2018年和2019年間, 于大豆R8期, 各小區隨機選取15株植株, 調查大豆單株有效莢數、單株粒數及百粒重; 選取長6 m的未取樣大豆帶測產, 在脫粒并曬干至籽粒含水量約為13.5%時, 測定籽粒產量。
1.4 數據分析
本研究所列結果為3次重復測定值的平均值, 使用Microsoft Excel 2016處理和分析數據, 采用統計分析軟件SPSSv.22軟件對數據進行方差分析和差異顯著性測驗(ANOVA, LSD, 顯著性水平為a=0.05)。利用Origin作圖。圖表中數據為平均值±標準誤。
2 結果與分析
2.1 外源植物生長調節對大豆碳代謝的影響
2.1.1 套作大豆莖、葉、莢果中可溶性糖含量 大豆莖稈、葉片可溶性糖含量隨著生育時期呈先增后減趨勢, 莢果可溶性糖含量呈連續增加趨勢(表1)。R5期, 調節劑處理下大豆莖稈、葉片可溶性糖含量顯著高于CK; 分別在6-BA和S3307處理下最高, 較CK分別增加28.8%和19.2%; 調節劑處理下莢果可溶性糖含量較CK呈增加趨勢。R8期, 調節劑處理下莖稈、葉片中的可溶性糖含量顯著低于CK, 分別在DTA-6和6-BA處理下最低, 較CK分別降低29.3%和20.2%; 調節劑處理下莢果可溶性糖含量顯著高于CK, 在DTA-6處理下最高, 較CK增加19.7%。
2.1.2 套作大豆葉片蔗糖代謝相關酶活性 隨生育期變化, 大豆葉片中SS、SPS、Inv酶活性的變化呈先上升后下降的趨勢, 并在R4期達到峰值(圖1)。R2期, DTA-6處理下大豆葉片SS、SPS、Inv酶活性顯著高于CK, 較CK分別增加22.9%、54.3%和20.4%, 與S3307差異不顯著。R4期, 調節劑處理下SS和SPS酶活性顯著高于CK, 分別在6-BA和S3307處理下最高, 較CK分別顯著增加25.0%和33.0%; 調節劑處理增加Inv酶活性, 在S3307處理下最高, 較CK增加55.7%, 各調節劑處理間差異不顯著。R6期, 調節劑處理下SS和SPS酶活性顯著高于CK, 分別在DTA-6、6-BA處理下最高, 較CK分別增加33.3%和41.1%; 調節劑處理增加Inv酶活性, 在6-BA處理下達到顯著水平, 較CK增加41.1%, 與DTA-6差異不顯著。
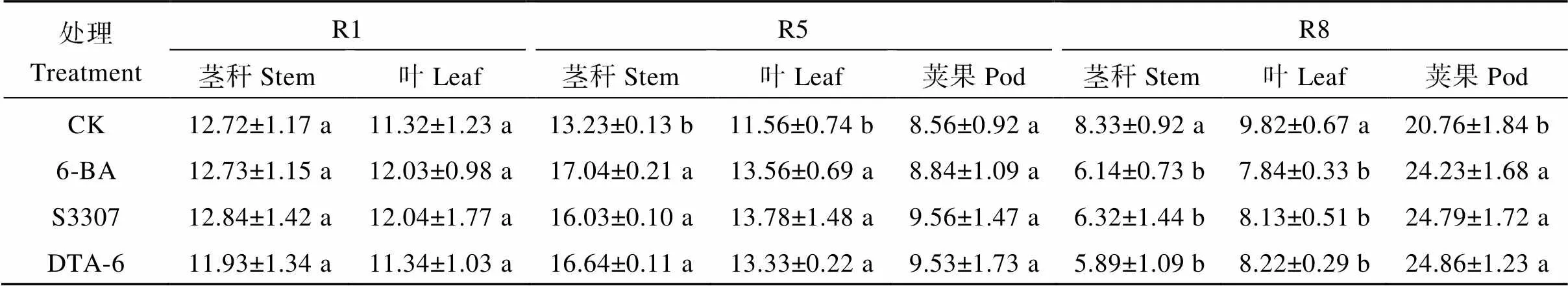
表1 外源噴施植物生長調節劑對套作大豆可溶性糖含量的影響
CK: 清水對照; 6-BA: 6-芐基腺嘌呤; DTA-6: 2-N,N-二乙氨基乙基己酸酯; S3307: 烯效唑。R1: 初花期; R5: 始粒期; R8: 成熟期。不同小寫字母表示處理在0.05水平下差異顯著。
CK: water control; 6BA: 6-benzylaminopurine; DTA-6: diethyl aminoethyl hexanoate; S3307: uniconazole. R1: beginning flower stage; R5: beginning seed stage; R8: mature stage. Different lowercase letters indicate significant differences among different regulator treatments at the 0.05 probability level.
2.2 外源植物生長調節對大豆植株各器官碳、氮含量, 碳/氮比值的影響
2.2.1 各器官總碳含量 從R3到R5期, 各器官中碳含量逐漸增加(表2)。R3期, 調節劑處理增加大豆莖稈、莢中的碳含量, 在S3307達到顯著水平, 較CK分別增加3.7%和3.5%; 調節劑處理下葉片碳含量較CK呈增加趨勢。R5期, 調節劑處理下大豆葉片氮素含量顯著高于CK, 在DTA-6處理下最高, 較CK增加1.3%; 調節劑處理下大豆莖稈、莢皮碳含量較CK呈增加趨勢。
2.2.2 各器官總氮含量 大豆莖稈、葉片、莢皮中的氮素含量隨生育時期逐漸下降, 籽粒中氮素含量逐漸增加(表3)。R3期, 6-BA和DTA-6處理下葉片氮含量顯著高于CK, 較CK分別增加7.8%和8.0%, 與S3307差異不顯著; 調節劑處理后的大豆莖稈、莢皮中的氮含量較CK呈下降趨勢。R5期, 6-BA和DTA-6處理下大豆莖稈氮含量顯著高于CK, 較CK分別增加25.8%和24.8%; S3307處理下大豆葉片氮含量顯著低于CK, 較CK降低23.2%, 各調節劑處理間差異不顯著; 調節劑處理下大豆莢皮氮含量顯著高于CK, 在S3307最高, 較CK增加28.0%。
2.2.3 大豆各器官碳/氮比值 調節劑處理增加R3期大豆莖稈和莢果中的C/N比值, 降低大豆葉片中的C/N比值; 在R5期呈現出相反變化趨勢(表4)。R3期, S3307處理下大豆莖稈C/N比值顯著高于CK, 較CK增加7.8%, 與6-BA和DTA-6處理差異不顯著; 6-BA和DTA-6處理下大豆葉片中C/N比值顯著低于CK, 較CK分別降低7.0%和6.8%, 與S3307差異不顯著; 調節劑處理下大豆莢果C/N比值較CK呈增加趨勢。R5期, 6-BA和DTA-6處理下大豆莖稈C/N比值顯著低于CK和S3307, 較CK分別降低27.7%和21.0%; S3307處理下大豆葉片C/N比值顯著高于CK, 較CK增加31.4%, 與6-BA和DTA-6處理差異不顯著; 調節劑處理下大豆莢皮C/N比值顯著低于CK, 在6-BA最小, 較CK降低20.8%; 調節劑處理下大豆籽粒C/N比值較CK呈降低趨勢。
不同小寫字母表示處理在0.05水平下差異顯著。R2: 盛花期; R4: 盛莢期; R6: 鼓粒期。處理同表1。
Different lowercase letters indicate significant differences among different regulator treatments at the 0.05 probability level. R2: blooming flower stage; R4: blooming pod stage; R6: full seed stage. Treatments are the same as those given in Table 1.
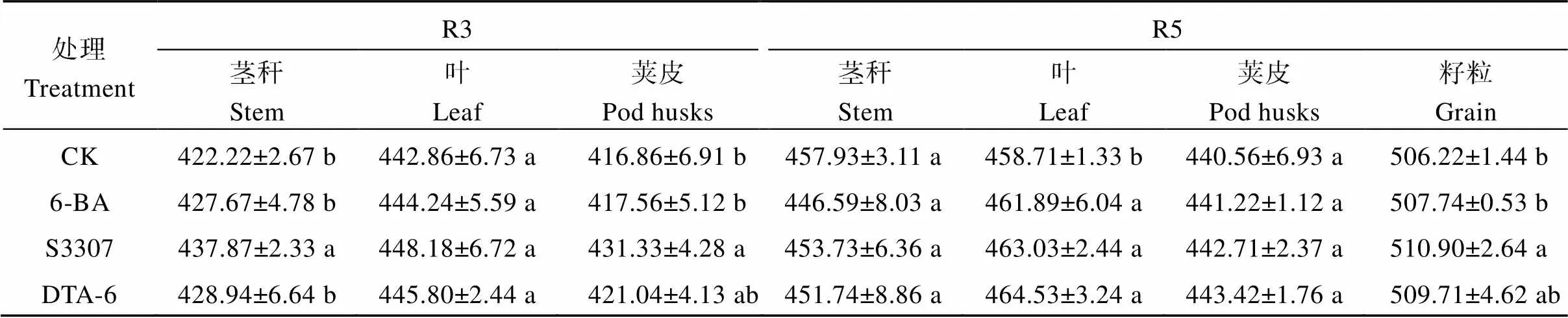
表2 外源噴施植物生長調節劑對套作大豆莖、葉、莢皮和籽粒的碳素含量的影響
不同小寫字母表示處理在0.05水平下差異顯著。R3: 始莢期; R5: 始粒期。處理同表1。
Different lowercase letters indicate significant differences among different regulator treatments at the 0.05 probability level. R3: beginning pod stage; R5: beginning seed stage. Treatments are the same as those given in Table 1.
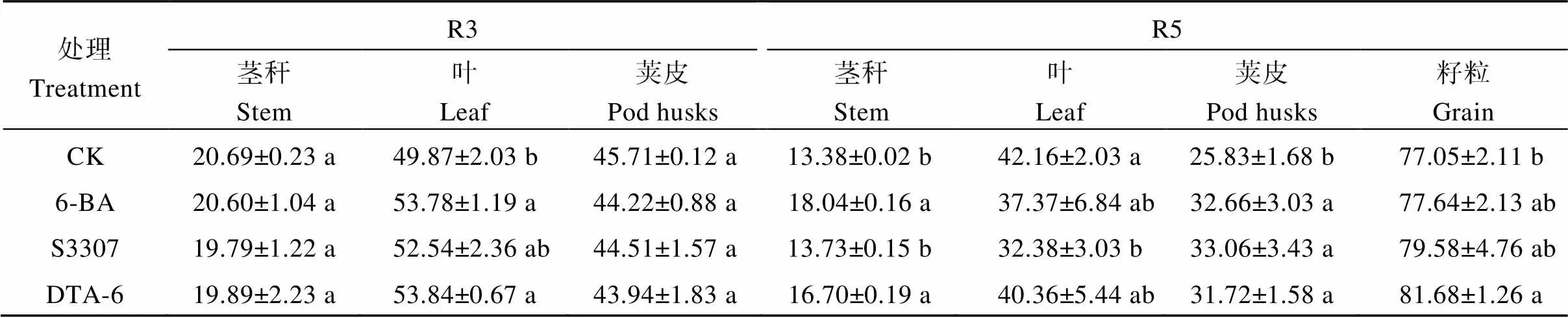
表3 外源噴施植物生長調節劑對大豆莖、葉、莢皮和籽粒的氮素含量的影響
不同小寫字母表示處理在0.05水平下差異顯著。R3: 始莢期; R5: 始粒期。處理同表1。
Different lowercase letters indicate significant differences among different regulator treatments at the 0.05 probability level. R3: beginning pod stage; R5: beginning seed stage. Treatments are the same as those given in Table 1.
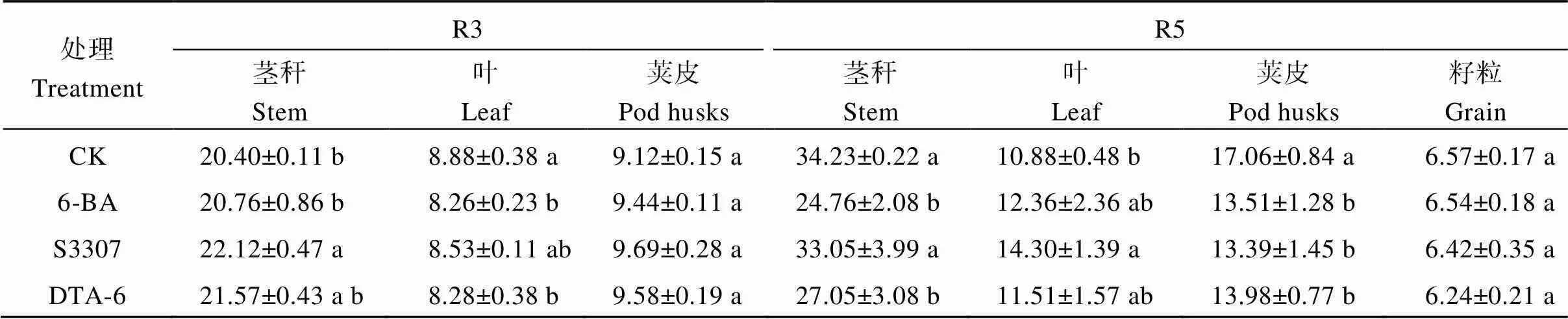
表4 外源噴施植物生長調節劑對大豆莖、葉和莢果的C/N的影響
不同小寫字母表示處理在0.05水平下差異顯著。R3: 始莢期; R5: 始粒期。處理同表1。
Different lowercase letters indicate significant differences among different regulator treatments at the 0.05 probability level. R3: beginning pod stage; R5: beginning seed stage. Treatments are the same as those given in Table 1.
2.3 外源植物生長調節劑對大豆花莢脫落的影響
外源調節劑處理會提高大豆的開花數和結莢數,降低花莢脫落率, 以DTA-6效果最佳, 且不同年份大豆開花數、結莢數、落花數之間存在差異(表5)。DTA-6處理下大豆開花數、結莢數顯著高于CK, 2年較CK分別增加10.4%、15.0%和5.2%、8.1%, 與6-BA、S3307處理差異不顯著。調節劑處理下大豆落花數、落莢數(6-BA除外)和落花率較CK呈下降趨勢。DTA-6處理下大豆落莢率、花莢脫落率顯著低于CK, 2年較CK分別降低15.0%、7.1%和29.4%、10.8%, 與S3307處理差異不顯著。
2.4 外源植物生長調節劑對大豆產量及產量構成的影響
調節劑處理對大豆單株有效莢數、單株粒數、產量有極顯著影響, 對大豆百粒重有顯著影響, 不同年份下大豆單株有效莢數、產量有差異(表6)。DTA-6處理下2年大豆單株有效莢數、單株粒數顯著高于CK, 較CK分別增加25.4%、35.6%和32.9%、33.2%, 顯著高于6-BA處理。調節劑處理下2年大豆百粒重顯著高于CK, 在S3307處理下最大, 較CK分別增加3.6%和2.1%。調節劑處理下2年大豆產量顯著高于CK, 在DTA-6處理下最大, 較CK分別增加41.3%和37.6%, 顯著高于6-BA和S3307處理。
3 討論
3.1 植物生長調節劑對大豆花莢脫落的影響
在本研究中, 調節劑處理對大豆單株莢數、單株粒數、產量的影響達到極顯著水平, 調節劑處理下大豆單株有效莢數、單株粒數、百粒重、產量顯著高于CK。玉米–大豆帶狀套作復合種植模式中, 受玉米蔭蔽影響, 大豆群體處于光能截獲劣勢, 光合作用減弱, 葉片碳代謝受到抑制, 作物整體代謝能力較弱[27], 大豆開花數結莢數減少, 且易出現莢而不實現象[28]。前人研究表明, 大豆單株有效莢數、單株粒數與大豆單株產量呈極顯著正相關關系[8], 花莢脫落率與大豆單株莢數、單株粒數、單株產量呈顯著負相關關系[2]。在開花期到始粒期期間, 大豆開花結莢數及其脫落率對大豆單株有效莢數的形成起決定性作用[26]。2年數據表明, 調節劑處理顯著增加套作大豆開花數、結莢數, 降低大豆落莢數、落莢率、花莢脫落率, 顯著提高大豆單株有效莢數與產量, 這與馮乃杰等[29]的研究結果相符合。
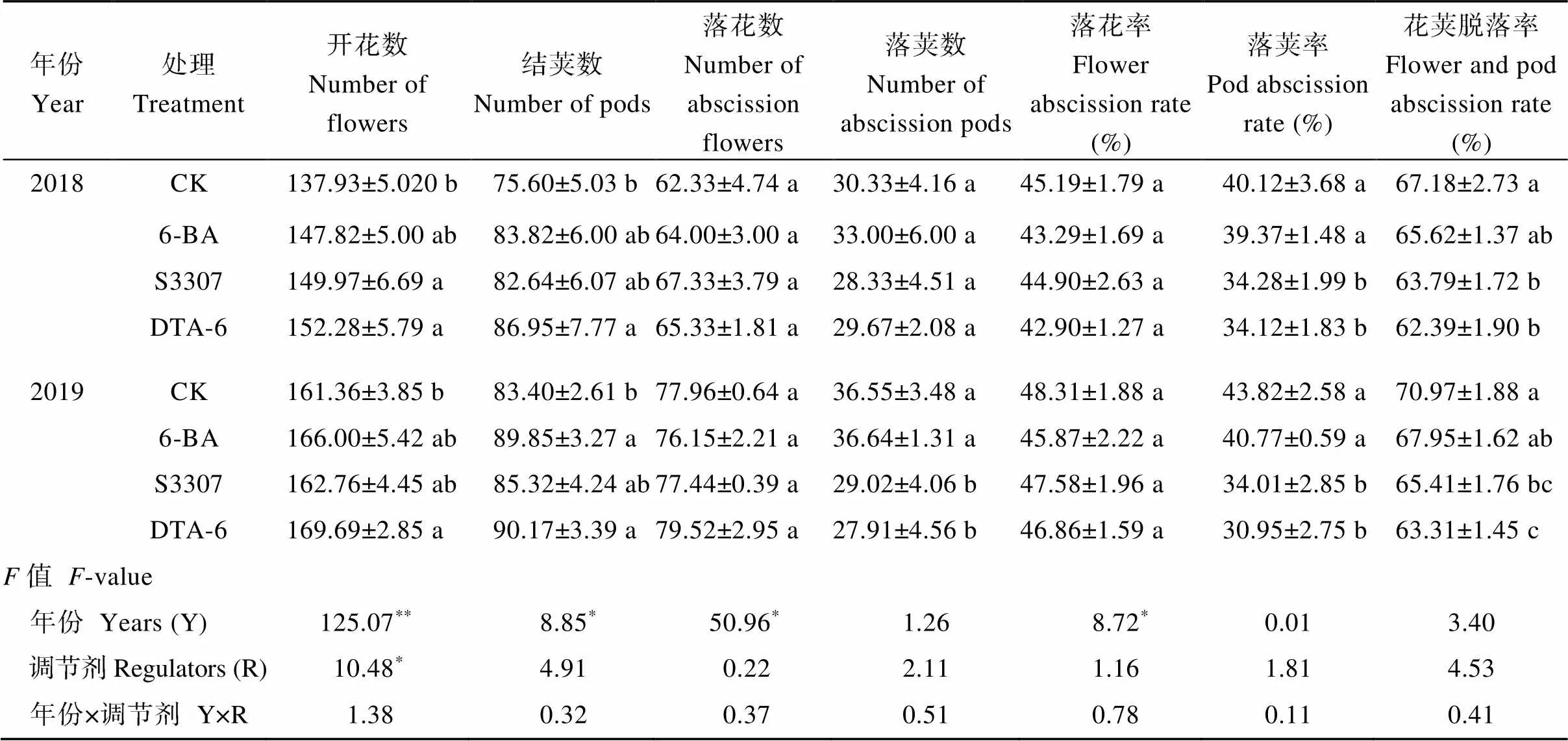
表5 外源噴施植物生長調節劑對套作大豆花莢脫落數及脫落率的影響
處理同表1。不同小寫字母表示處理在0.05水平下差異顯著。*表示在0.05水平顯著,**表示在0.01水平顯著。
Treatments are the same as those given in Table 1. Different lowercase letters indicate significant differences among different regulator treatments at the 0.05 probability level.*represents significant difference at the 0.05 probability level;**represents significant difference at the 0.01 probability level.
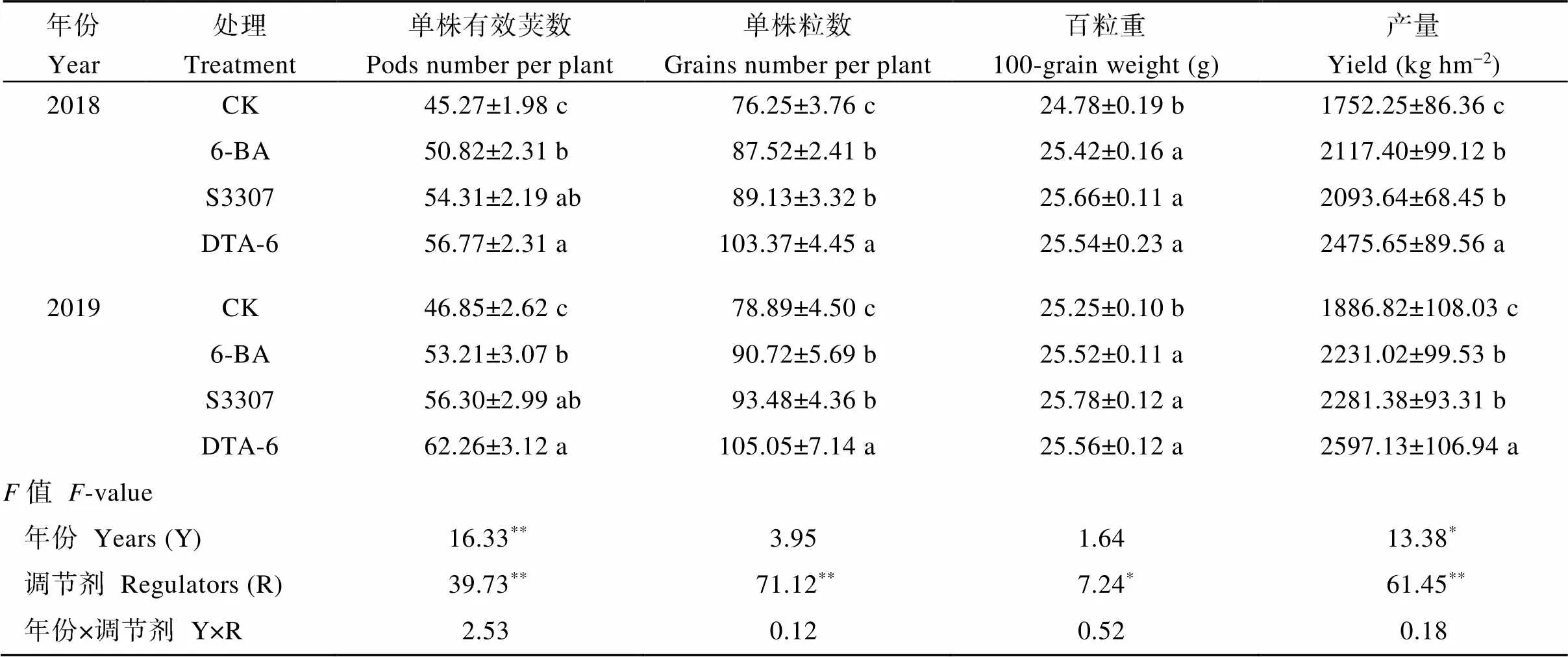
表6 外源噴施植物生長調節劑對大豆產量與產量構成的影響
處理同表1。不同小寫字母表示處理在0.05水平下差異顯著。*表示在0.05水平顯著,**表示在0.01水平顯著。
Treatments are the same as those given in Table 1. Different lowercase letters indicate significant differences among different regulator treatments at the 0.05 probability level.*represents significant difference at the 0.05 probability level;**represents significant difference at the 0.01 probability level.
3.2 植物生長調節劑對大豆糖代謝的影響
同化物供應的有效性, 決定了不同器官間的養分分配狀況; 合理的養分分配有利于減少不同器官間的養分競爭, 對生殖器官的形成和發育起著重要的調控作用[15]。可溶性糖是典型的碳水化合物, 其含量水平能反映作物體內作為有效態營養物的碳水化合物和能量水平[30]。本試驗結果表明, 調節劑處理提高R5期大豆葉片內可溶性糖的含量, 增強葉片內碳水化合物的供應; 促進后期莖、葉中碳水化合物向籽粒的轉移, 這與閆艷紅等[13]的研究結論相一致。糖的合成、水解與轉移, 需要多種酶的共同參與; 蔗糖合成酶能夠催化蔗糖的合成; 蔗糖磷酸合成酶調控光合產物向蔗糖和淀粉分配; 轉化酶能將蔗糖水解為同等量的果糖和葡萄糖, 是參與蔗糖水解的關鍵酶[31]。葉片內蔗糖合成酶和蔗糖磷酸合成酶活性的增強, 有利于提高葉片可溶性糖、蔗糖和淀粉含量[25]。葉片內Inv酶活性提高有利于促進葉片內不同碳水化合物之間的相互轉化, 增強碳水化合物的轉運能力。本研究中, 調節劑處理提高各時期大豆葉片SS、SPS、Inv酶活性, 提高葉片內碳水化合的合成和向籽粒運輸的能力, 為花莢的形成提供能量供應和物質基礎, 這與趙黎明等[32]和宋春燕等[33]的研究結果相一致。
3.3 碳氮協調與大豆花莢脫落
前人研究表明, 大豆開花結莢期間的養分比例失調, 器官間的養分競爭是誘導花莢脫落的重要原因[34]。營養生長時期積累足夠的干物質, 是協調器官間養分比例、維持花莢正常發育的必要條件。蔣利[35]研究發現, 在玉米–大豆帶狀套作模式下, 花莢脫落率高的大豆品種, 盛花期到盛莢期仍有大量的干物質分配到莖稈和葉片的生長上, 增強了莖葉與花莢之間的養分競爭。本研究結果表明, R3期, 調節劑處理增加葉片和莢果碳、氮含量, 葉片C/N比值降低, 莢果C/N比值上升, 說明調節劑處理在維持葉片代謝強度的同時, 提高莢果代謝強度, 促進莢的形成, 降低莢的脫落。劉春娟等[36-37]研究發現, 葉面噴施DTA-6和S3307能延緩葉片衰老, 促進生育后期大豆葉片活性, 提高莢中碳、氮代謝強度, 促進葉片中蔗糖、果糖和淀粉向莢果的轉移, 使更多的碳水化合物用于莢的形成, 進而提高大豆產量, 與本研究結果相一致。
4 結論
外源噴施6-BA、DTA-6、S3307增強葉片SS、SPS、Inv酶的活性, 提高始粒期大豆莖、葉、莢中的可溶性糖含量, 增強后期籽粒中可溶性糖的積累, 優化葉片內碳水化合物的合成及轉運; 調節劑處理下R3期葉片C/N比值降低, R5期葉片C/N比值增加, 維持不同時期碳氮代謝的動態平衡; 調節劑處理促進大豆開花結莢, 降低落莢率及花莢脫落率, 顯著提高套作大豆的單株有效莢數與產量, 其中以DTA-6處理的增產效果最佳。
[1] Chen P, Song C, Liu X M, Zhou L, Yang H, Zhang X N, Zhou Y, Du Q, Pang T, Fu Z D, Wang X C, Liu W G, Yang F, Shu K, Du J B, Liu J, Yang W Y, Yong T W. Yield advantage and nitrogen fate in an additive maize–soybean relay intercropping system., 2019, 657: 987–999.
[2] 蔣利, 雍太文, 張群, 肖靜, 楊歡, 楊文鈺. 種植模式和施氮水平對大豆花莢脫落及產量的影響. 大豆科學, 2015, 34: 843–849. Jiang L, Yong T W, Zhang Q, Xiao J, Yang H, Yang W Y. Effect of different planting patterns and N application rates on abscission of flower and pod of soybean and yield., 2015, 34: 43–849 (in Chinese with English abstract).
[3] Liu B, Qu D N. Effects of shading on spatial distribution of flower and flower abscission in field-grown three soybeans in Northern China., 2015, 27: 629–635.
[4] 崔洪秋, 馮乃杰, 孫福東, 劉濤, 李建英, 杜吉到, 韓毅強, 鄭殿峰. DTA-6對大豆花莢脫落纖維素酶和基因表達的調控. 作物學報, 2016, 42: 51–57. Cui H Q, Feng N J, Sun F D, Liu T, Li J Y, Du J D, Han Y Q, Zheng D F. Regulation of DTA-6 by abscission cellulase andgene expression in flowers and pods of soybean., 2016, 42: 51–57 (in Chinese with English abstract).
[5] Duarte L, Clemente J, Caixeta I, Senoski M, Aquino L. Dry matter and nutrient accumulation curve in cabbage crop., 2019, 32: 679–689.
[6] Zhang J H, He N P, Liu C G, Xu L, Chen Z, Li Y, Wang R M, Yu G R, Sun W, Xiao C W, Reich P. Variation and evolution of C:N ratio among different organs enable plants to adapt to N-limited environments., 2020, 26: 2534–2543.
[7] 胡志輝, 汪艷杰, 陳禪友. 噴施細胞分裂素對豇豆花莢脫落率及花莢酶活性的影響. 植物科學學報, 2016, 34: 439–445. Hu Z H, Wang Y J, Chen C Y. Effects of spraying cytokinin on abscission rate and enzymic activity of flowers and pods of cowpea., 2016, 34: 439–445 (in Chinese with English abstract).
[8] 萬燕, 閆艷紅, 楊文鈺. 不同氮肥水平下葉面噴施烯效唑對套作大豆生長和氮代謝的影響. 浙江大學學報(農業與生命科學版), 2012, 38: 185–196. Wan Y, Yan Y H, Yang W Y. Effects of foliar spraying uniconazole on growth and nitrogen metabolism of relay strip intercropping soybean under different nitrogen levels.(Agric Life Sci), 2012, 38: 185–196 (in Chinese with English abstract).
[9] 鄭殿峰, 宋春艷. 植物生長調節劑對大豆氮代謝相關生理指標以及產量和品質的影響. 大豆科學, 2011, 30: 109–112. Zheng D F, Song C Y. Effects of plant growth regulators on nitrogen metabolism related indicators and yield in soybean., 2011, 30: 109–112 (in Chinese with English abstract).
[10] Zhang M C, Duan L S, Tian X L, He Z P, Li J M, Wang B M, Li Z H. Uniconazole-induced tolerance of soybean to water deficit stress in relation to changes in photosynthesis, hormones and antioxidant system., 2007, 164: 709–717.
[11] Wen D X, Li Y, He L F, Zhang C Q. Transcriptome analysis reveals the mechanism by which spraying diethyl aminoethyl hexanoate after anthesis regulates wheat grain filling., 2019, 19: 327.
[12] Kim J H, Shon J Y, Lee C K, Yang W, Yoon Y W, Yang W H, Kim Y G, Lee B W. Relationship between grain filling duration and leaf senescence of temperate rice under high temperature., 2011, 122: 207–213.
[13] 閆艷紅, 萬燕, 楊文鈺, 王小春, 雍太文, 劉衛國, 張新全. 葉面噴施烯效唑對套作大豆花后碳氮代謝及產量的影響. 大豆科學, 2015, 34: 75–81. Yan Y H, Wan Y, Yang W Y, Wang X C, Yong T W, Liu W G, Zhang X Q. Effect of spraying uniconazole on carbon and nitrogen metabolism and yield of relay strip intercropping soybean., 2015, 34: 75–81 (in Chinese with English abstract).
[14] Zhou W G, Chen F, Zhao S H, Yang C Q, Meng Y J, Shuai H W, Luo X F, Dai Y J, Yin H, Du J B, Liu J, Fan G Q, Liu W G, Yang W Y, Shu K. DA-6 promotes germination and seedling establishment from aged soybean seeds by mediating fatty acid metabolism and glycometabolism., 2018, 70: 101–114.
[15] Liu C J, Feng N J, Zheng D F, Cui H Q, Sun F D, Gong X W. Uniconazole and diethyl aminoethyl hexanoate increase soybean pod setting and yield by regulating sucrose and starch content., 2019, 99: 748–758.
[16] Roitsch T, Ehne? R. Regulation of source/sink relations by cytokinins., 2000, 32: 359–367.
[17] Choi J, Hwang I. Cytokinin: perception, signal transduction, and role in plant growth and development., 2007, 50: 98–108.
[18] Du J B, Han T F, Gai J Y, Yong T W, Sun X, Wang X C, Yang F, Liu J, Shu K, Liu W G, Yang W Y. Maize–soybean strip intercropping: achieved a balance between high productivity and sustainability., 2018, 17: 747–754.
[19] Yang W T, Miao J Q, Wang X W, Xu J C, Lu M J, Li Z X. Corn-soybean intercropping and nitrogen rates affected crop nitrogen and carbon uptake and C:N ratio in upland red soil., 2018, 41: 1890–1902.
[20] Yang F, Huang S, Gao R C, Liu W G, Yong T W, Wang X C, Wu X L, Yang W Y. Growth of soybean seedlings in relay strip intercropping systems in relation to light quantity and red: far-red ratio., 2014, 155: 245–253.
[21] Liu X, Rahman T, Song C, Su B Y, Yang F, Yong T W, Wu Y S, Zhang C Y, Yang W Y. Changes in light environment, morpho-logy, growth and yield of soybean in maize–soybean intercropping systems., 2017, 200: 38–46.
[22] Chen P, Du Q, Liu X M, Zhou L, Hussain S, Lei L, Song C, Wang X C, Liu W G, Yang F, Shu K, Liu J, Du J B, Yang W Y, Yong T W. Effects of reduced nitrogen inputs on crop yield and nitrogen use efficiency in a long-term maize–soybean relay strip intercropping system., 2017, 12: e0184503.
[23] 徐光域, 顏軍, 郭曉強, 劉嵬, 李曉光, 茍小軍. 硫酸-苯酚定糖法的改進與初步應用. 食品科學, 2005, 26(8): 342–346. Xu G Y, Yan J, Guo X Q, Liu W, Li X G, Gou X J. The betterment and apply of phenol-sulphate acid method., 2005, 26(8): 342–346 (in Chinese with English abstract).
[24] 鄧小燕, 王小春, 楊文鈺, 張群. “麥/玉/豆”模式下氮肥運籌對玉米碳氮代謝的影響. 草業學報, 2012, 21(4): 52–61. Deng X Y, Wang X C, Yang W Y, Zhang Q. Effects of nitrogen strategies on carbon and nitrogen metabolism of maize in wheat/maize/soybean relay intercropping system., 2012, 21(4): 52–61 (in Chinese with English abstract).
[25] Chopra J, Kaur N, Gupta A K. Ontogenic changes in enzymes of carbon metabolism in relation to carbohydrate status in developing mungbean reproductive structures., 2000, 53: 539–548.
[26] Dyer D J, Carlson D R, Cotterman C D, Sikorski J A, Ditson S L. Soybean pod set enhancement with synthetic cytokinin analogs., 1987, 84: 240–243.
[27] Cui L, Yang W Y, Huang N, Liu J, Wang Y L, Wang X H, Liu Y, Yan S. Effects of maize plant types on dry matter accumulation characteristics and yield of soybean in maize–soybean intercropping systems., 2015, 26: 2414–2420.
[28] Yan Y H, Wan Y, Liu W G, Wang X C, Yong T W, Yang W Y, Zhao L L. Influence of seed treatment with uniconazole powder on soybean growth, photosynthesis, dry matter accumulation after flowering and yield in relay strip intercropping system., 2015, 18: 295–301.
[29] 馮乃杰, 宋莉萍, 劉金輝, 鄭殿峰. 不同時期PGRS對大豆花莢脫落率及纖維素酶活性的影響. 中國油料作物學報, 2011, 33: 253–258. Feng N J, Song L P, Liu J H, Zheng D F. Regulation of plant growth regulators on abscission rate and cellulase activity of soybean flowers and pods., 2011, 33: 253–258 (in Chinese with English abstract).
[30] 劉海坤, 劉小寧, 黃玉芳, 葉優良. 不同氮水平下小麥植株的碳氮代謝及碳代謝與赤霉病的關系. 中國生態農業學報, 2014, 22: 782–789. Liu H K, Liu X N, Huang Y F, Ye Y L. Effect of nitrogen application on carbon and nitrogen metabolisms and relationship between carbon metabolism and wheat scab., 2014, 22: 782–789 (in Chinese with English abstract).
[31] Liu C G, Wang Y J, Pan K W, Zhu T T, Li W, Zhang L. Carbon and nitrogen metabolism in leaves and roots of Dwarf Bamboo (Yi) subjected to drought for two consecutive years during sprouting period., 2014, 33: 243–255.
[32] 趙黎明, 馮乃杰, 鄭殿峰. 植物生長調節劑對大豆莢皮同化物代謝及糖分積累的影響. 植物科學學報, 2008, 26: 407–411. Zhao L M, Feng N J, Zheng D F. Effects of plant growth regulators on assimilation metabolism and sugar accumulation in pod husks of soybean., 2008, 26: 407–411 (in Chinese with English abstract).
[33] 宋春艷, 馮乃杰, 鄭殿峰. 植物生長調節劑對大豆葉片碳代謝相關生理指標及產量品質的影響. 干旱地區農業研究, 2011, 29(3): 91–95. Sun C Y, Feng N J, Zheng D F. Effects of plant growth regulators on carbon metabolism related indicators in soybean leaves., 2011, 29(3): 91–95 (in Chinese with English abstract).
[34] Truong Q, Koch K, Yoon Jo M, Everard J D, Shanks J V. Influence of carbon to nitrogen ratios on soybean somatic embryo (cv. Jack) growth and composition., 2013, 64: 2985–2995.
[35] 蔣利. 凈套作條件下不同施N量對大豆植株形態、花莢脫落和產量的影響. 四川農業大學碩士學位論文, 四川雅安, 2015. Jiang L. The Effects of N Application on Plant Morphology, Flower, and Pod Abscission and Yield of Soybean in Monoculture and the Strip Intercropping System. MS Thesis of Sichuan Agriculture University, Ya’an, Sichuan, China, 2015 (in Chinese with English abstract).
[36] 劉春娟, 馮乃杰, 鄭殿峰, 宮香偉, 孫福東, 石英, 崔洪秋, 張盼盼, 趙晶晶. 植物生長調節劑S3307和DTA-6對大豆源庫碳水化合物代謝及產量的影響. 中國農業科學, 2016, 49: 657–666. Liu C J, Feng N J, Zheng D F, Gong X W, Sun F D, Shi Y, Cui H Q, Zhang P P, Zhao J J. Effects of plant growth regulators S3307 and DTA-6 on carbohydrate content and yield in soybean., 2016, 49: 657–666 (in Chinese with English abstract).
[37] 劉春娟, 馮乃杰, 鄭殿峰, 孫福東, 劉濤, 崔洪秋, 趙晶晶. S3307和DTA-6對大豆葉片生理活性及產量的影響. 植物營養與肥料學報, 2016, 22: 626–633. Liu C J, Feng N J, Zheng D F, Sun F D, Liu T, Cui H Q, Zhao J J. Effects of plant growth regulators S3307 and DTA-6 on the leaf physiological activity and yield of soybean., 2016, 22: 626–633 (in Chinese with English abstract).
Effect of exogenous plant growth regulators on carbon-nitrogen metabolism and flower-pod abscission of relay strip intercropping soybean
LUO Kai1, XIE Chen1, WANG Jin1, WANG Tian1, HE Shun2, YONG Tai-Wen1,*, and YANG Wen-Yu1
1College of Agriculture, Sichuan Agriculture University / Key Laboratory of Crop Physiology, Ecology and Cultivation in Southwest China, Ministry of Agriculture and Rural Affairs / Sichuan Engineering Research Center for Crop Strip Intercropping System, Wenjiang 611130, Sichuan, China;2Chengdu Seed Management Station / Chengdu Agricultural Product Quality and Safety Center, Qingyang 610072, Sichuan, China
Maize shading inhibited the growth of soybean at the seedling stage, reduced the number of soybean flowers and pods, and decreased the yield of soybean under maize–soybean relay strip intercropping system. To explore the effect of exogenous plant growth regulators (PGRs) on flowering, pod setting, and yield of soybean is important for relay strip intercropping soybean production. Field experiments were used one-factor randomized block design to investigate the effect of foliage spraying 6-Benzylaminopurine (6-BA), diethyl aminoethyl hexanoate (DTA-6) and uniconazole (S3307) at the beginning of flowering stage on abscission of flowers and pods, leaf carbon and nitrogen metabolism, and yield formation in relay strip intercropping soybean. The results demonstrated that spraying PRGs improved the sucrose synthetase (SS), sucrose phosphate synthetase (SPS) and invertase (Inv) activities in soybean leaves. Exogenous PGRs increased soluble sugar content in stems, leaves, and pods of soybean at the beginning of grain-filling stage, promoted soluble sugar transport from stem and leaves to pods. Exogenous PGRs increased carbon and nitrogen content and decreased the C:N ratio in soybean leaves at the beginning of podding stage. PGRs increased the carbon content, decreased nitrogen content, and increased the C:N ratio in soybean leaves at beginning of grain-filling stage. Foliar spraying PGRs increased the number of flowers and pods, reduced the abscission pod number, and decreased the ratio of pods abscission and flowers-pods abscission, the effect of DTA-6 treatments was better than others. The pods per plant and yield of soybean in 2018 and 2019 under DTA-6 treatment were increased by 25.4%, 41.3% and 32.9%, 37.6% as compared with CK, respectively. Foliar spraying PGRs increased the activities of SPS, SS, and Inv enzymes in soybean leaves, coordinated the carbon-nitrogen metabolism in soybean organs, promoted the soybean flowering and pod-setting, decreased the abscission of pods, increased the pods per plant and yield of relay strip intercropping soybean, the production increasing effect of DTA-6 were better than others.
maize–soybean relay strip intercropping; flowers and pods abscission; plant growth regulators; carbon and nitrogen metabolism; yield
10.3724/SP.J.1006.2021.04129
本研究由國家重點研發計劃項目(2018YFD0201006)和國家現代農業(大豆)產業技術體系建設專項(CARS-04-03A)資助。
This study was supported by the National Key Research and Development Program of China (2018YFD0201006) and the China Agriculture Research System (Soybean) (CARS-04-03A).
雍太文, E-mail: scndytw@qq.com
E-mail: 592234005@qq.com
2020-06-16;
2020-10-14;
2020-11-06.
URL: https://kns.cnki.net/kcms/detail/11.1809.S.20201105.1555.010.html
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
小讀者(2021年2期)2021-03-29 05:03:48
今日農業(2020年20期)2020-11-26 06:09:10
華人時刊(2019年13期)2019-11-17 14:59:54
文苑(2018年22期)2018-11-19 02:54:14
