少汗性外胚層發育不良患者EDA基因突變檢測及表型分析
2021-02-04 02:04:02吳君怡孫仕晨樊壯壯鄭靜蕾張劉陶馮海蘭
北京大學學報(醫學版) 2021年1期
關鍵詞:基因突變
吳君怡, 余 淼, 孫仕晨, 樊壯壯, 鄭靜蕾, 張劉陶, 馮海蘭, 劉 洋△, 韓 冬△
(1. 北京大學口腔醫學院·口腔醫院,修復科 國家口腔疾病臨床研究中心 口腔數字化醫療技術和材料國家工程實驗室 口腔數字醫學北京市重點實驗室,北京 100081; 2. 深圳市口腔醫院種植修復科,深圳 518001; 3. 北京醫院口腔科 國家老年醫學中心 中國醫學科學院老年醫學研究院,北京 100010)
外胚層發育不良(ectodermal dysplasia, ED)是一類涉及兩種和兩種以上外胚層來源器官發育異常的疾病,可造成全身多個系統的缺陷,至今已報道170余種病理損害,具有顯著的遺傳性[1-2],其中最常見的是少汗性外胚層發育不良(hypohidrotic ectodermal dysplasia, HED)。HED是一種罕見的先天遺傳性疾病,新生兒發病率約為十萬分之七[3]。HED患者的典型臨床表現為“三聯征”,包括汗腺發育異常、毛發稀疏和牙齒發育異常,具體表現為汗腺分泌功能低下甚至喪失導致的少汗或無汗,皮膚薄且干燥,常有濕疹;毛發卷曲且稀少;多數乳恒牙甚至全部乳恒牙先天缺失,余留牙為錐形牙或過小牙[2]。此外,患者常伴有特殊面容,如:前額突出,眼周色素沉著,眼周皮膚多皺,鞍狀鼻,上頜發育不足,上、下唇外翻等[4]。目前, HED已明確的遺傳方式有3種:常染色體顯性、常染色體隱性以及X連鎖遺傳。65%~75%的HED病例為X連鎖遺傳,主要與定位于人類染色體Xq12-13.1的ectodysplasinA(EDA)基因突變相關,而常染色體顯性或隱性遺傳的HED多與ectodysplasinAreceptor(EDAR,定位于2q11-13)基因及EDARassociateddeathdomain(EDARADD,定位于1q42.3-43)基因的突變相關[5-6]。
EDA基因屬于腫瘤壞死因子(tumor necrosis factor,TNF)超家族成員,共有9個外顯子,約95%的基因變異發生在第1、3、5、8、9外顯子上[7-8]。EDA蛋白是Ⅱ型跨膜蛋白,包含一個短的胞內區、單一跨膜區和一個長的胞外區[9]。EDA蛋白有4個重要的功能結構域,分別為:(1)跨膜域:由第1外顯子參與編碼,負責跨膜運輸;(2)前體蛋白加工酶furin識別序列:由第3外顯子參與編碼,是EDA前體蛋白發生裂解的識別位點;(3)膠原樣結構域:由第5~6外顯子參與編碼,負責維持TNF結構域正常折疊構象的形成,促進TNF結構域的多聚化;(4)TNF結構域:由第7~9外顯子參與編碼,是EDA與下游受體EDAR結合的重要區域[10]。研究報道顯示,EDA基因變異中37.8%的變異位點位于TNF結構域,22.0%的變異位點位于膠原樣結構域,7.3%的變異位點位于跨膜域, 7.3%的變異位點位于前體蛋白加工酶furin識別序列[11]。
小鼠模型研究發現,在胚胎發育第14天,可在上皮基底層檢測到Eda的表達;在胚胎發育第15天,可在蕾狀期牙胚上皮中檢測到Eda的表達,隨后,Eda可在牙胚間充質中持續性表達至帽狀期末期[12]。此外,EDA在人體多種組織均有表達,包括毛囊、神經外皮、胸腺、骨和上皮等,可參與人類牙齒、毛發、皮膚等組織器官的形成[8, 13-14]。EDA基因突變除與HED相關外,還可導致單純型先天缺牙[3, 15]。
截至目前,人類基因突變數據庫(http://www.hgmd.cf.ac.uk/ac/index.php)已收錄300余種EDA基因突變類型。盡管已明確的與HED有關的EDA基因突變超過140余種,約50%是錯義突變[11, 16],但是新的EDA基因突變仍有待發現,與牙齒缺失位點有關的基因型-表型相關性分析仍有待完善。本研究擬對近年來臨床收集到的HED患者進行EDA基因致病突變的篩查,旨在發現新檢出的突變,并對HED患者的臨床表型及缺失牙位進行基因型-表型相關性分析。
1 資料與方法
1.1 病例收集
在就診于北京大學口腔醫院修復科的先天缺牙患者中,收集臨床表現為毛發稀少、汗腺功能異常、X線檢查證實缺牙區無恒牙胚的患者。本試驗獲得北京大學口腔醫院倫理委員會批準(批準號:PKUSSIRB-201736082), 研究對象包括患者和健康人均簽署知情同意書。
1.2 基因組DNA提取
提取患者及家系成員的外周靜脈血,經乙二胺四乙酸抗凝劑抗凝后,使用全血基因組DNA提取試劑盒(康為世紀生物科技有限公司,中國), 按照操作說明提取基因組DNA。對于不便獲取外周靜脈血的患者及家系成員,使用GeneFiXTM唾液收集裝置(Isohelix公司,英國)采集唾液,使用GeneFiXTM唾液DNA分離試劑盒提取基因組DNA。提取完成的基因組DNA置于-20 ℃保存。
1.3 聚合酶鏈式反應、Sanger測序及單克隆測序
使用引物設計網站(www.ncbi.nlm.nih.gov/tools/primer-blast)設計涵蓋EDA基因編碼區外顯子的8對引物,使用DNA聚合酶(博邁德公司,中國)對先證者DNA進行聚合酶鏈式反應(polymerase chain reaction,PCR), 擴增產物經瓊脂糖凝膠電泳鑒定后進行Sanger測序(睿博興科生物公司,中國)。使用DNA序列分析軟件Chromas2.6對DNA序列峰圖進行分析,使用NCBI數據庫BLAST工具(https://blast.ncbi.nlm.nih.gov/Blast.cgi)與正常人群的EDA基因序列(NM_001399.5)進行比對。對于在女性攜帶者中檢測出的EDA雜合突變,利用CloneSmarter-TOPO載體克隆試劑盒(CloneSmarter公司,美國)對PCR擴增產物進行單克隆測序進一步驗證變異(睿博興科生物公司,中國)。
1.4 突變蛋白功能預測及致病性分級
在人類基因組整合數據庫(https://gnomad.broadinstitute.org/)中檢索檢測出的EDA基因突變位點,去除最小等位基因頻率(minor allele frequency, MAF)大于0.01的單核苷酸多態性(single nucleotide polymorphism, SNP)位點,使用在線軟件Polyphen 2(http://genetics.bwh.harvard.edu/pph2/)、MutationTaster(http://www.mutationtaster.org/)和Provean(http://provean.jcvi.org/index.php)分別對檢測出的EDA基因錯義突變進行蛋白質功能預測。使用《美國醫學遺傳學和基因組學會遺傳變異致病性分級指南》對檢測出的突變進行致病性評估。
1.5 保守性分析及蛋白二級、三級結構預測
通過Uniprot數據庫(http://www.uniprot.org/)獲取不同物種的EDA蛋白序列,下載FASTA格式數據,使用T-coffee數據庫(http://www.ebi.ac.uk/Tools/msa/tcoffee)對檢測出的EDA蛋白突變位點進行保守性分析。
通過Swiss-model數據庫(https://swissmodel.expasy.org/)進行EDA蛋白同源建模,對突變蛋白三維結構進行預測。獲取EDA-A1剪切體模板(第233~391位氨基酸),將模板導入到PyMOL 2.4.0軟件中,通過修改突變位點的氨基酸,對突變EDA蛋白的三維結構進行預測分析。對于突變位點位于第233位氨基酸前的突變,通過MutationTaster數據庫(http://www.mutationtaster.org/)獲取突變后的氨基酸序列,使用Psipred 4.0軟件對突變EDA蛋白進行二級結構預測分析。
1.6 EDA基因突變相關的HED患者恒牙缺失牙位統計分析
根據臨床和X線檢查結果,對檢測出EDA基因突變的HED患者的缺失恒牙進行匯總和統計分析。計量資料以均數±標準差表示,左、右側同名牙位缺失牙數目的對比使用配對數據t檢驗。分別計算不同牙位缺牙率(該牙位缺失牙數目/該牙位牙總數), 采用PRISM 8軟件分別對不同牙位缺牙率進行卡方檢驗,并繪制缺牙率圖,P<0.05為差異有統計學意義。
2 結果
2.1 變異檢測及功能預測
本研究在12個HED家系中發現8個家系的9例HED男性患者分別攜帶8個EDA基因突變,檢出率為66.67%(圖1), 分別是:移碼突變c.619delG(p.Gly207Profs*73),無義突變c.676C>T(p.Gln226*),錯義突變c.164T>C(p.Leu55Pro)、c.457C>T(p.Arg153Cys)、c.466C>T(p.Arg156Cys)、c.584G>A(p.Gly195Glu)、c.673C>T(p.Pro225Ser)和c.905T>G(p.Phe302Cys),其中,#632先證者和#688先證者(圖1A、B)攜帶相同的EDA基因突變c.457C>T(p.Arg153Cys);#362先證者(圖1E)攜帶2個不同的EDA基因突變c.673C>T(p.Pro225Ser)和c.676C>T(p.Gln226*)。在檢出的8個EDA基因突變中,c.619delG(p.Gly207Profs*73)、c.676C>T(p.Gln226*)、c.164T>C(p.Leu55Pro)、c.673C>T(p.Pro225Ser)和c.905T>G(p.Phe302Cys)是未報道過的新檢出的突變。
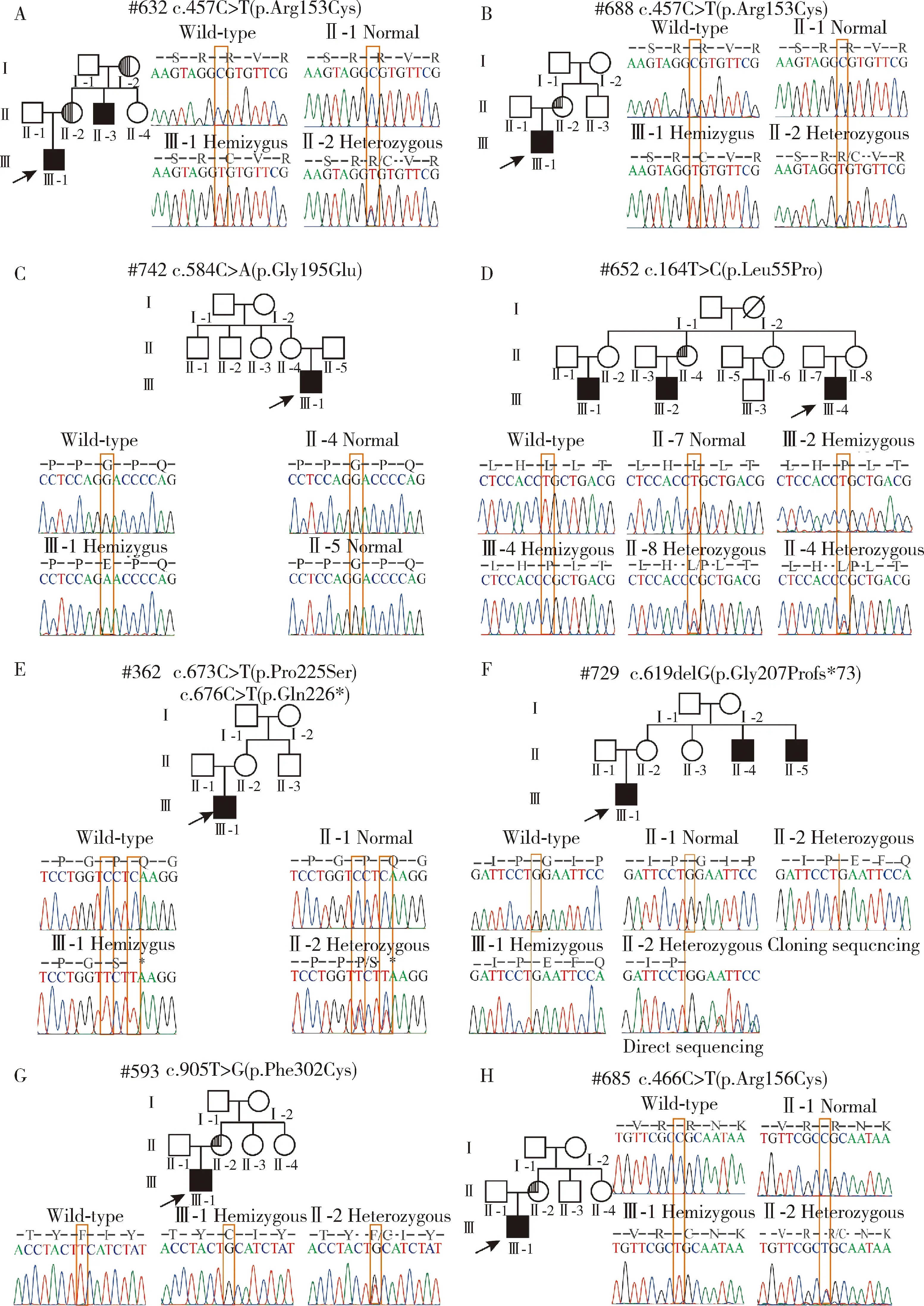
A-H, pedigree and EDA gene sequencing chromatograms, semicircle shade indicates female carrier with hypodontia and hypotrichosis, quarter-circle shades indicate female carriers with hypodontia.
檢出的8個EDA基因突變在人類基因組整合數據庫中均未檢索到,綜合使用在線軟件Polyphen-2、Mutation Taster以及Provean預測變異蛋白的功能影響,根據《美國醫學遺傳學和基因組學會遺傳變異致病性分級指南》,發現這些突變表現為不同程度的致病性(表1)。對8個家系可獲得的家系成員進行EDA基因突變檢測,#742先證者的母親(圖1C)未檢測到EDA基因突變,證明#742先證者攜帶的EDA基因突變是新發突變(denovomutation);其余7個家系中先證者的母親均可檢測到與先證者一致的EDA基因突變,證明這些家系中先證者的EDA基因突變均分別遺傳自其母親。
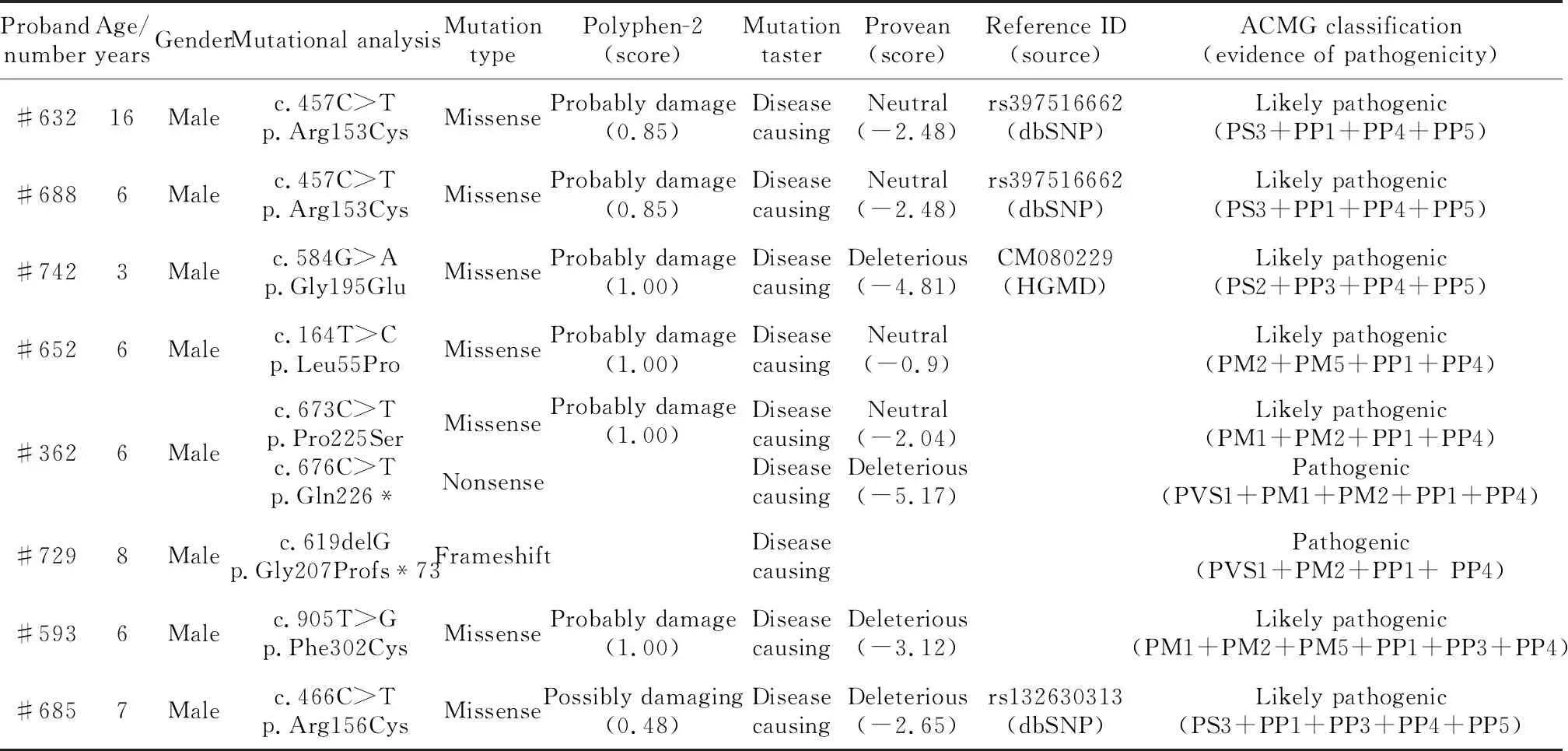
表1 本研究中檢測出的EDA基因突變
2.2 HED 患者的臨床表型
本研究在8個家系中共篩查出9例攜帶EDA基因突變的男性HED患者(圖2),均有毛發稀疏、皮膚干燥、汗腺功能減退以及HED相關特殊面容(圖2A、D、H、O)。口腔表型也十分典型,均表現為多數牙乳恒牙先天缺失,余留前牙呈錐形或過小牙(圖2B、C、F、I、J、L~N、P~V)。值得注意的是,#688先證者除典型的毛發稀疏,汗腺功能減退以及多數牙缺失外,還表現出先天性乳頭乳暈缺如(圖2K), 這一表型在EDA基因突變的HED患者中尚未見報道。
2.3 保守性分析及變異蛋白結構預測
對檢測出的EDA基因的6個錯義突變位點進行保守性分析,發現這些突變位點在不同物種間均高度保守(圖3)。對錯義突變p.Phe302Cys進行EDA突變蛋白三維結構預測分析,發現第302位氨基酸由含有苯環結構的苯丙氨酸替換為含有二硫鍵的半胱氨酸,使蛋白構象發生改變(圖4A~D)。對錯義突變p.Leu55Pro、p.Arg153Cys、p.Arg156Cys、p.Gly195Glu、p.Pro225Ser、p.Gln226* 和p.Gly207Profs*73進行EDA蛋白二級結構預測,發現p.Leu55Pro、p.Arg153Cys、p.Arg156Cys和p.Gly195Glu導致EDA蛋白二級結構中的α-螺旋和β-折疊發生改變(圖4E~I);無義突變p.Gln226*導致EDA蛋白第266位氨基酸后的結構缺失,蛋白顯著截短(圖4J);移碼突變p.Gly207Profs*73導致EDA蛋白自207位氨基酸開始發生序列改變,在之后的73個氨基酸位置發生翻譯提前中止,使蛋白長度顯著截短(圖4K)。
2.4 EDA突變相關的HED患者恒牙缺牙數目和缺失牙位的統計分析
對檢測出的9例攜帶EDA基因突變的男性HED患者的恒牙缺牙數目及缺失牙位進行統計分析(表2),分別記錄不同患者上頜右側、上頜左側、下頜右側、下頜左側缺失牙數目情況。經統計分析后得到如下結果:(1)患者缺失牙位呈左、右對稱分布,左、右側同名牙之間差異無統計學意義(P>0.05),因此后續將左、右側缺失牙數目進行合并統計;(2)患者平均缺失牙數目為(13.86±4.49)顆,其中上頜平均缺失牙數目為(13.14±5.76)顆,缺失率為73.02%,下頜平均缺失牙數目為(14.57±3.05)顆,缺失率為80.95%,上、下頜缺失牙數目差異無統計學意義(P>0.05);(3)在上頜牙列中,側切牙和第二前磨牙缺失率最高,為100%(18/18),而上頜中切牙缺失率最低,為11.11%(2/18),其次為上頜第一磨牙,缺失率為55.56%(10/18,圖5A),上頜這四個牙位缺牙率對比平均缺牙率76.98%(194/252)差異均有統計學意義(P<0.05);(4)在下頜牙列中,側切牙缺失率最高,為100%(18/18),而下頜第一磨牙缺失率最低,為55.56%(10/18)(圖5B),下頜這兩個牙位缺牙率對比平均缺牙率76.98%(194/252)差異均有統計學意義(P<0.05)。

A, the face photograph of #632 proband; B-C, the intraoral photographs of #632 proband; D-E, the detailed photographs show sparse eyebrows of #632 proband and his mother; F-G, the panoramic radiographs of #632 proband and his mother; H, the face photograph of #688 proband; I-J, the intraoral photographs of #688 proband; K, the photograph shows absent nipple and areola of #688 proband; L-M, the panoramic radiographs of #688 proband and #742 proband; N, the intraoral photograph of #652 proband; O, the face photograph of #652 proband; P-Q, the panoramic radiographs of #652 proband and his cousin; R, the cephalometric radiograph of #362 proband; S, the intraoral photograph of #729 proband; T-V, the panoramic radiographs of #729 proband, #593 proband and #685 proband. The black block represents the missing tooth, the red arrow refers to the tapered tooth. MAX, maxillary dental arch; MAND, mandibular dental arch.
3 討論
自1996年Srivastava等[8]成功克隆并定位EDA基因以來,不斷有研究從遺傳學和分子生物學的角度對EDA基因突變與HED相關性進行探索。目前已證實EDA基因突變可以導致X連鎖遺傳的HED的發生,其新檢出的突變類型持續被報道。本研究在8個家系的9個男性HED患者中檢測出8個EDA基因突變:c.164T>C(p.Leu55Pro)、c.457C>T(p.Arg153Cys)、c.466C>T(p.Arg156Cys)、c.584G>A(p.Gly195Glu)、c.619delG(p.Gly207Profs*73)、c.673C>T(p.Pro225Ser)、c.676C>T(p.Gln226*)以及c.905T>G(p.Phe302Cys),其中,c.619delG(p.Gly207Profs*73)、c.676C>T(p.Gln226*)、c.164T>C(p.Leu55Pro)、c.673C>T(p.Pro225Ser)和c.905T>G(p.Phe302Cys)是新檢出的突變。檢測出的8個突變中,有5個突變為C>T(G>A)突變,可能由細胞中常見的胞嘧啶自動脫氨基作用所致。
以上8個突變中,p.Leu55Pro 位于EDA蛋白的跨膜域內,由于該結構域與蛋白極性密切相關[17],因此我們推測該突變引起的跨膜結構改變,可能影響EDA的正常表達。研究發現EDA前體蛋白須在細胞膜上裂解并形成水溶性的信號分子后,方能與下游受體結合發揮作用,而furin識別序列結構域正是EDA前體蛋白發生裂解的識別位點[17],因此,位于furin識別序列結構域內的p.Arg153Cys和p.Arg156Cys,極有可能導致EDA前體蛋白裂解障礙,無法形成正常的水溶性的信號分子,進而影響其與下游受體的結合,妨礙EDA蛋白的正常功能[10];p.Gly195Glu、p.Gly207Profs*73、p.Pro225Ser和p.Gln226* 位于膠原樣結構域內,該結構域主要是負責維持腫瘤壞死因子(tumor necrosis factor, TNF)結構域形成正常折疊構象,促進TNF結構域形成三聚體[17-18],該結構域內的移碼突變p.Gly207Profs*73和無義突變p.Gln226*導致EDA蛋白截短,產生嚴重的功能影響。盡管功能預測結果表明,p.Pro225Ser為可能致病的突變,但先證者同一條X染色體同時攜帶錯義突變 p.Pro225Ser和無義突變p.Gln226*,可能通過協同作用嚴重影響EDA蛋白的功能;p.Phe302Cys位于EDA蛋白的TNF結構域內,研究證明此高保守的結構域以同源三聚體的形式與受體EDAR結合,由此推測,p.Phe302Cys導致的TNF結構域的構象改變,可能影響EDA與EDAR的結合,進而導致下游核轉錄因子kappa-B(nuclear factor kappa-B, NF-κB)信號通路激活受阻[17, 19]。NF-κB信號通路參與胚胎發育早期的外胚間充質轉化,決定著牙齒、毛發和皮膚等多種組織和器官的形態發生[20],因此,本研究所報道的EDA基因突變,為EDA-EDAR-NF-κB級聯通路障礙在外胚層器官發育異常中的致病作用提供了新的理論證據。

A, location of EDA mutations identified in this study, reported mutations are in black, novel mutations are in red; B, schematic diagram of the EDA gene; C, conservation analysis of affected amino acids in the EDA protein among 8 different species. TM, transmembrane domain; TNF, tumor necrosis factor.
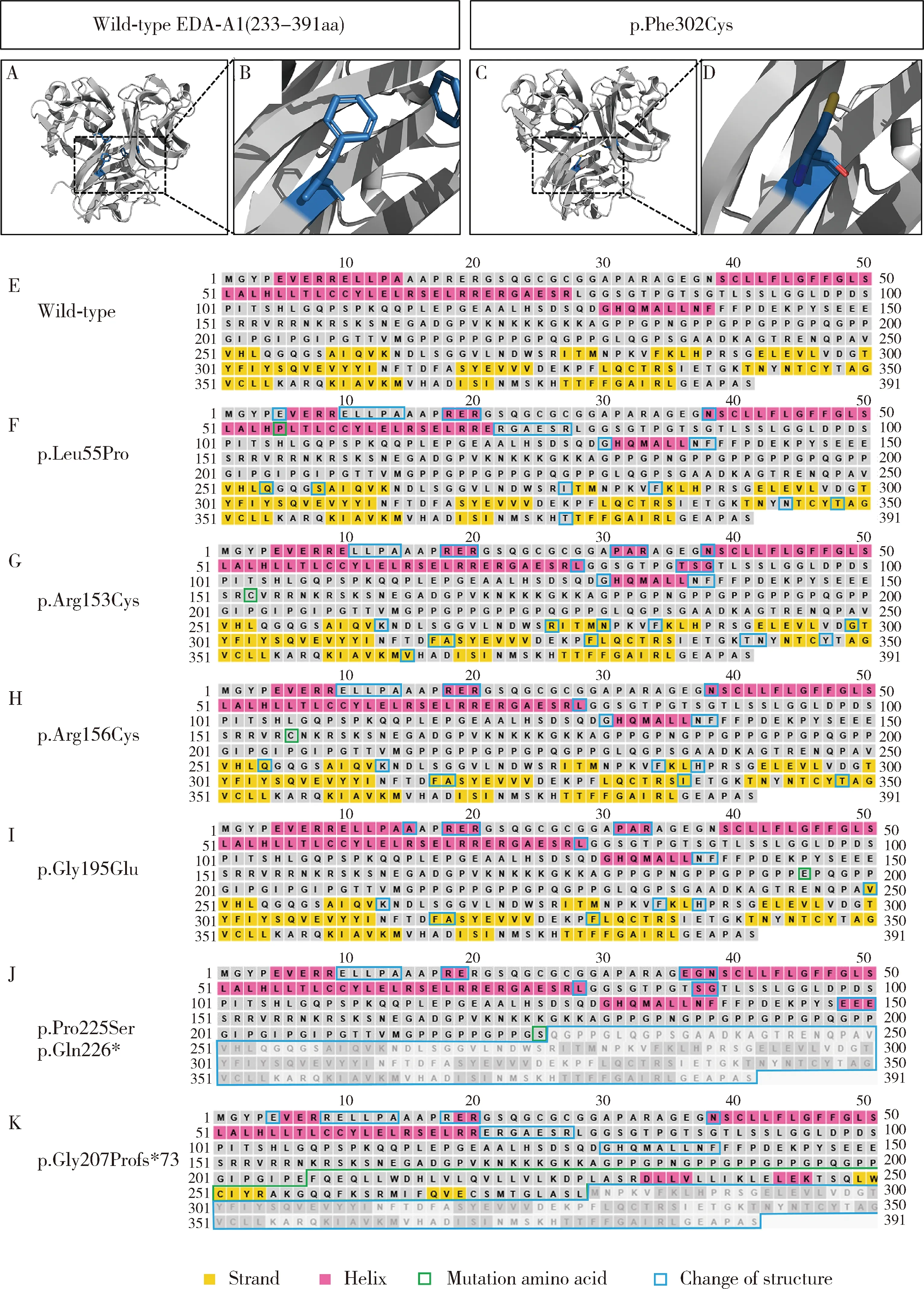
A-B, the tertiary structure of wild-type EDA-A1; C-D, the tertiary structure of the p.Phe302Cys mutant; E, the secondary structure of wild-type EDA protein; F-K, the secondary structure of the p.Leu55Pro, p.Arg153Cys, p.Arg156Cys, p.Gly195Glu, p.[pro225ser; Gln226*] and p.Gly207Profs*73 mutants.
本研究檢測出的9例EDA基因突變的男性HED患者均表現為汗腺發育異常、毛發稀疏、牙齒發育不全的三聯征。#688先證者除具有典型的HED表型外,還表現出乳頭和乳暈的先天缺如,這一乳腺相關的嚴重表型在男性HED患者中尚未見報道。對HED疾病模型小鼠Tabby的相關研究發現,Tabby鼠表現出乳頭小且平,乳腺分泌功能障礙[21]。同時,有學者曾對EDA基因雜合突變的女性攜帶者的胸部發育進行研究,發現39%的患者表現出乳頭扁平,10%的患者出現多乳頭癥狀,67%的患者存在哺乳功能障礙[22]。以上研究均顯示,EDA基因對乳腺發育的調控具有重要的作用,因此,#688先證者的乳頭和乳暈先天缺如的表型可能與EDA基因突變密切相關,有待后續研究進一步明確證實其致病機制。
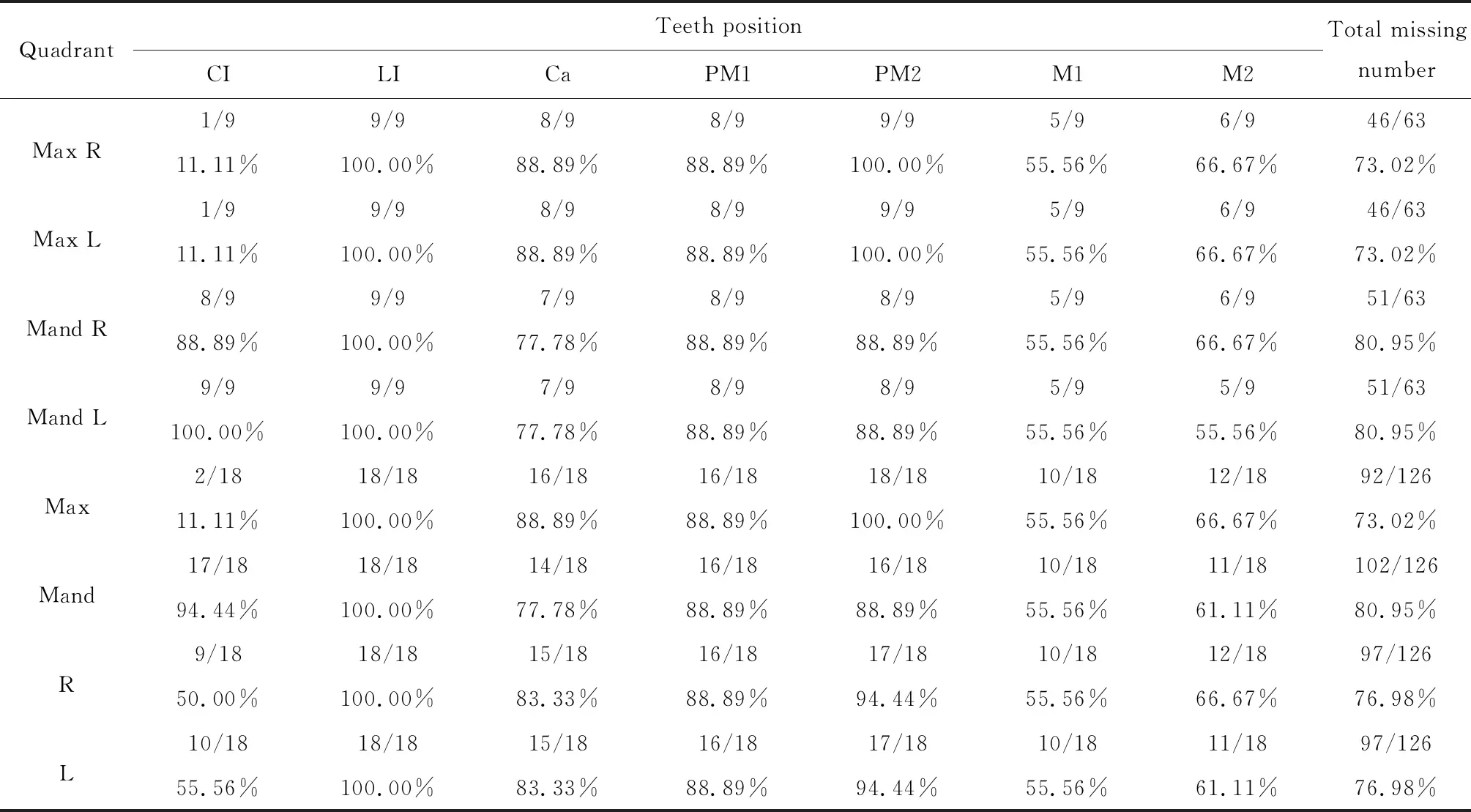
表2 EDA基因突變的HED患者缺牙數目和缺失牙位

A and B, percentage of missing tooth positions at each maxillary and mandibular dentition in HED patients with EDA mutations. Max, maxillary; Mand, mandibular; CI, central incisor; LI, lateral incisor; Ca, canine; PM1, first premolar; PM2, second premolar; M1, first molar; M2, second molar. Statistical significant P-value is marked with *P< 0.05, △P < 0.01, ◇P < 0.001, and ※P < 0.000 1; HED, hypohidrotic ectodermal dysplasia.
值得關注的是,同一家系中攜帶相同致病突變的男性HED患者(#652先證者與其表兄)缺牙表型有所差異, #652先證者恒牙列僅余留2顆上頜中切牙,而其表兄恒牙列余留2顆上頜中切牙和1顆下頜磨牙。相同的致病突變,缺牙表型卻存在差異,可能存在如下原因:(1)不同患者對相同的基因突變的敏感度有所差異,導致EDA基因突變在不同個體中的表現度不同,進而導致臨床表型的輕重不一;(2)在遺傳過程中,DNA甲基化、組蛋白修飾、基因組印記的變化都能造成臨床表型不同的變化[23]。
本研究進一步匯總了這9例EDA基因突變的男性HED患者的恒牙牙位缺失情況,并歸納了缺失牙位的特點,研究發現上頜最易缺失牙位是側切牙和第二前磨牙,下頜最易缺失牙位是側切牙,這說明在牙胚發育過程中,上頜側切牙、第二前磨牙和下頜側切牙的牙胚對EDA基因的敏感性最高;而上頜中切牙、上頜第一磨牙和下頜第一磨牙較少發生缺失,說明上頜中切牙、上頜第一磨牙和下頜第一磨牙的牙胚發育對EDA基因的敏感性較低,這一結果與以往研究基本一致[24]。此外,本研究中EDA基因突變的男性HED患者的余留前牙呈錐形或過小牙。對Tabby鼠的研究也發現其前牙近遠中徑變小、呈錐形牙冠的形態[25],這均提示我們EDA基因可能與前牙形態發生有關,其具體發揮的功能及影響值得進一步研究。
綜上,本研究在12個HED家系中共篩查出8例家系患者分別攜帶8個不同類型的EDA基因突變,其中5個突變為未曾報道的新檢出的突變。有文獻報道,約60%的HED患者攜帶EDA基因突變,未檢出EDA基因突變的HED患者可能攜帶HED已知致病基因EDAR(HED患者中占比16%, 后同)、EDARADD(2%)、Wntfamilymember10A(WNT10A,16%)基因突變,還有8%的HED患者可能攜帶其他未知基因突變[26]。對缺牙表型的分析是研究牙發育相關基因功能的關鍵環節,也有利于臨床醫生預測先天缺牙患者可能的致病基因,從而有的放矢地進行遺傳學檢測。本研究通過分析得出EDA基因突變與上頜側切牙、第二前磨牙和下頜側切牙缺失的相關性最高,與上頜中切牙、上頜第一磨牙和下頜第一磨牙缺失的相關性最低,這不僅豐富了EDA的基因型-表型譜,更為遺傳咨詢和產前診斷提供了強有力的臨床證據。
猜你喜歡
英語世界(2023年6期)2023-06-30 06:29:10
中國醫學影像學雜志(2021年6期)2021-08-13 08:43:36
中國生殖健康(2020年2期)2021-01-18 02:51:26
小學生導刊(2018年13期)2018-06-29 03:49:00
中國生殖健康(2018年2期)2018-01-12 13:57:51
現代檢驗醫學雜志(2016年4期)2016-11-15 02:01:14
中國現代醫學雜志(2015年26期)2015-12-23 11:04:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:44
中華皮膚科雜志(2014年4期)2014-12-19 12:55:49
中國神經精神疾病雜志(2014年1期)2014-03-01 03:23:22
