鱖(Siniperca chuatsi)后部側線系統胚后發育*
2021-02-03 01:44:54張瑞祺趙金良
海洋與湖沼 2021年1期
關鍵詞:系統
張瑞祺 趙金良 ①
(1. 上海海洋大學 農業農村部淡水水產種質資源重點實驗室 上海 201306; 2. 上海海洋大學 水產動物遺傳育種中心上海市協同創新中心 上海 201306)
側線系統是魚類重要感覺器官, 與魚類索餌、捕食、集群、洄游和生殖等行為有密切聯系(Montgomery et al, 2013; Bird et al, 2014; Butler et al, 2015; Brown et al, 2016; Dow et al, 2018)。大部分硬骨魚類均具有機械感受側線系統, 主要由神經丘結構與側線管道構成, 其中神經丘分為兩大類, 管道神經丘(canal neuromast, CN)與表面神經丘(superfical neuromast,SN), 側線管道因所在位置不同, 分為顱部側線管道系統與軀干側線管道系統(Denton et al, 1989; Münz,1989; Webb, 2013)。對魚類側線結構的研究是解讀其側線功能特征、生活習性和對應生境關系的重要手段。
二類神經丘分布于魚類體表, 其中部分神經丘內含于特定側線管道中成為管道神經丘, 而完全暴露于外部的神經丘稱為表面神經丘(Webb, 2013; Bird et al, 2014)。管道神經丘借由側線管道配合, 與魚類周邊水體加速度信息的感知有關, 常在魚類捕食、避敵行為中發揮重要作用, 而表面神經丘與周邊水體絕度流速的感知有關, 在對周邊水體波動的基本感知與警戒中發揮作用(Denton et al, 1989; Münz, 1989;Webb, 2013)。二類神經丘數量、分布、結構特點及側線管道結構特征直接反應了魚類側線系統的進化與特化程度, 如洞穴魚(Astyanax mexicanus)在陰暗洞穴中視覺感官嚴重退化, 側線系統極為發達, 全身覆蓋大量密集排布的神經丘, 側線感覺極為靈敏(Yoshizawa et al, 2011; Worsham et al, 2019); 又如大頭犀孔鯛(Porornitra capito)和大洋黑鱈(Melanonus zugmuyeri), 側線管道較寬的管徑與內含的大型管道神經丘使其在深海底棲生活中對周邊捕食目標的感知極為靈敏(Marshall, 1996)。魚類顱部側線系統結構較為保守, 同一目、科魚類顱部側線系統在神經丘分布與側線管路結構方面常不具有明顯區別,而后部側線系統結構形式多樣, 常隨著生境的變化而發生結構性適應(Webb, 1989a, 2013; Pichon et al,2004; Bird et al, 2014 )。
鱖(Siniperca chuatsi)是我國特有的淡水經濟魚類(Liang et al, 2001; 孫海林等, 2019)。鱖屬于兇猛肉食性魚類, 具有獨特的攝食習性, 首先食性特殊, 終身食活魚, 拒食死餌, 其次捕食行為特殊, 常采取底棲伏擊的方式捕食小型魚類(梁旭方, 1994, 1995a, b;Liang et al, 2001), 而與其他肉食性魚類如大口黑鱸(Micropterus salmoides) 、 暗 紋 東 方 鲀 (Takifugu obscurus)、北美狗魚(Esox masquinongy)等所采取的主動捕食方式有顯著不同(New et al, 2000; 沈輝等,2010)。為解讀鱖特殊捕食習性, 已有部分學者對鱖捕食行為相關感官結構與功能進行了研究, 梁旭方(1995b)利用行為學實驗證明鱖主要依靠視覺、側線感覺進行捕食, 梁旭方(1996)利用組織學方法研究了鱖顱部側線管道系統, 發現鱖顱部側線管道屬于分枝型, 其結構可能與底棲生活相關。目前, 鱖側線系統的研究較為薄弱, 顱部側線管道研究尚不足以詮釋鱖側線系統的結構特征, 同時鱖在早期發育中, 開口必須以其他魚的初孵仔魚為食, 仔稚魚時期主動攝食, 幼魚至成魚階段逐漸采用底棲伏擊性捕食方式(魏開建等, 1996; 田文斐等, 2012; Zhang et al, 2019),對其重要感官——側線系統發育過程的研究有助于解讀鱖早期發育過程捕食習性的變化機制。
為此, 本研究利用DESAPI 神經丘染色、組織切片研究了鱖后部側線系統的發育過程、神經丘數量與排布特征, 利用掃描電鏡觀察了后部側線神經丘建立過程與結構特點。本研究是對鱖完整側線系統研究的重要補充, 后部側線系統發育過程與結構特點為其生活習性轉變提供了重要資料。
1 材料與方法
1.1 實驗材料
實驗用鱖魚苗取自上海浦東新區孫農水產養殖場。整個采樣期間水質情況保持不變(水溫25—26°C;溶氧: 5—8 mg/L; pH 7.0—7.5), 各批樣本的具體發育時間記為 n dpf (受精后第 n 天; dpf, days post fertilization)。基于鱖早期發育相關研究(魏開建等,1996; 田文斐等, 2012; Zhang et al, 2019), 將鱖分為仔魚(3—9 dpf, SL: 3.5—8.0 mm)、稚魚(10—23 dpf,SL: 9.0—25.0 mm)、幼魚(28—58 dpf, SL: 30—100 mm)三個階段。1—58 dpf 期間以天(d)為單位均勻采樣, 每次45 尾(其中供神經丘染色、組織切片和掃描電鏡觀察各 15 尾), 神經丘熒光染色用樣本為活體魚苗, 采樣后直接進行處理; 組織切片用樣本采樣后 1×PBS溶液洗凈, Bouin 固定液固定 12 h, 梯度酒精處理(50%→60%→70%→75%, 各梯度 40%), 將樣本于75%酒精中浸浴, 每次 1—2 d, 至樣本中殘留固定劑褪去后, 將樣本于75%酒精中室溫保存; 掃描電鏡樣本采樣后1×PBS 溶液洗凈, 2.5%戊二醛固定18 h, 供后續使用。
1.2 神經丘熒光染色
將1.1 中處理鱖魚苗進行浸于神經丘毛細胞標記物DESAPI (2-4-dimethylamino styryl-N-ethylpyridinium iodide; AAT Bioquest Inc)配制的熒光染色溶液中,DASPEI 染色濃度分別為 0.01% (3—12 dpf, SL:3—10 mm, 染色時間10 min), 0.05% (16—30 dpf, SL:10—30 mm, 染色時間 15 min), 0.15% (31—58 dpf,SL: 30—100 mm, 染色時間30 min)。染色結束后用蒸餾水將魚苗體表多余染劑漂洗干凈, 置于 Nikon SMZ25 熒光體視鏡下觀察, NIS-Elements 圖像采集系統拍照。神經丘命名同Webb(2013), 管道神經丘在所屬管道未閉合時記為前體管道神經丘(presumptive canal neuromast, PCN), 管道閉合后內含神經丘記為管道神經丘(canal neuromast, CN)。
1.3 組織切片
取1.1 中處理鱖魚苗梯度酒精脫水(75%→85%→90%→95%→100%), 各梯度20 min (95%→100%脫水兩次, 每次5 min), 二甲苯透明(具體時間以樣本通體透明為完成), 浸入 65°C 石蠟完成組織包埋。使用Leica RM 2016 切片機以 5 μm 厚度連續切片, HE 染色,中性樹膠封片。利用 Olympus IX73 顯微鏡觀察,Nikon DS-5Mc 圖像采集系統拍照。
1.4 掃描電鏡觀察
取1.1 中處理鱖魚苗置于1×PBS 溶液中將殘留固定劑洗凈, 后置于1×PBS 溶液中漂洗5 次將體表黏液洗凈, 每次 40 min。完成漂洗的樣本梯度酒精脫水(75%→85%→90%→95%→100%), 各梯度 20 min(95%→100%脫水兩次, 每次5 min), Emitech K850 干燥儀中臨界點干燥, Hitachi E-1010 噴金儀中噴金,HITACHI S-3400N 掃描電鏡觀察拍照。神經丘形貌指標測量方法(長、寬與毛細胞數)同Webb(2013)。
1.5 數據處理
神經丘形貌指標(長、寬與毛細胞數)均以平均值±標準差表示。使用 t-檢驗(t-test)、單因素方差分析(One-Way ANOVA)與最小顯著差異法多重比較(LSD)對各類神經丘形貌參數進行比較。數據比較使用SPSS 22 軟件(IBM)完成, 數據顯著性判定標準為P<0.05。
2 結果與分析
2.1 后部側線系統發育
2.1.1 軀干側線系統 4 dpf 仔魚未觀察到軀干神經丘(圖1a); 6 dpf 時, 在仔魚軀干觀察到一組軀干神經丘(L1—L6)(圖1b)。9 dpf 仔魚在軀干上有一行神經丘(L1—L7), 相鄰神經丘間隔約 6—7 個體節(圖 1c);12 dpf 時新增4—5 個神經丘位于L1和L2神經丘間隔處, 形成每一體節一個神經丘的排列方式(圖 1d);16 dpf, 新生神經丘生長在早期神經丘(L1—L3)之間(圖 1e), 保持每一體節一個的排列(圖 1f), 但后方早期神經丘間隔處(L4—L7)尚未出現新生神經丘(圖1g);23 dpf, 已生成的神經丘逐漸降于水平肌隔之下, 同時以每一體節一個的排列規律由鰓蓋后方延伸至尾柄基部(R0)(圖 2a), 此時在軀干前部已出現一行神經丘平行于早期神經丘(R1), 2—3 個體節分布一個神經丘(圖 2b); 在 30 dpf 時, 軀干神經丘明顯分為兩行:2—3 個體節分布一個神經丘的上行(R1)和一個體節分布一個的下行(R0) (圖2c, 圖3a); 37 dpf 時, R1的神經丘已延伸至尾部, 此時, 軀干側線管道尚未開始建立(圖 2d)。
R0的神經丘在側線發育中經歷了兩次重要變化,首先, 基礎神經丘(R0)通過出芽的方式在上下方向各生成一個附屬神經丘(44 dpf) (圖 2e, 圖 3b); 其次,整個 R0的神經丘在 50—55 dpf 時期開始下沉(圖 2f,圖3c), 此時軀干側線系統管道結構開始建立; 58 dpf,軀干已出現完整的側線系統管道系統, 而通過 R0的基礎神經丘出芽生成的位于 R0上方與下方、平行于R0的附屬神經丘, 已成為平行于軀干側線管道的游離神經丘(圖2g, 圖3d)。
2.1.2 尾鰭側線系統 尾鰭部分側線結構的發育出現于9 dpf, 在尾柄基部出現一終端神經丘(T0) (圖4a), 12 dpf 時 T0出現下移(圖 4b); 16 dpf 尾部鰭條出現于尾柄以下, 同時尾部脊軸上翹, T0出現分散, 80%的稚魚在尾柄處有三個上下排列的神經丘(圖 4c);19 dpf 尾柄的脊軸上翹明顯, 大部分稚魚此時在尾部脊軸下方均出現鰭條和沿著脊軸方向的三個終端神經丘(T1, T2, T3) (圖4d); 21 dpf 時尾部結構已和成魚相似, 尾鰭鰭條均勻分布, 尾柄基部由上至下依次排列著神經丘T1, T2, T3(圖4e); 23 dpf 時T1, T2, T3神經丘分別向后出芽生成附屬神經丘(圖4f)。37 dpf, 尾鰭神經丘已出現明顯排布規律, 在背側、腹側方向各出現兩列神經丘(背部兩列, C1和 C2; 腹部兩列, C3和C4), 同時一短行神經丘 C0位于 C2與 C3之間(圖 4g);44 dpf 時尾鰭神經丘排列特征已趨于穩定(圖4h)。
2.2 后部側線神經丘的建立
后部管道神經丘與表面神經丘建立之初形狀均為圓形, 此時二類神經丘表面并無明顯分區, 感受帶與神經丘大小相同(圖 5), 初期前體管道神經丘(5—10 dpf)平均長寬均為 4—5 μm(表 1), 毛細胞數12—14(表2); 表面神經丘初期較難識別, 16 dpf 軀干觀察到表面神經丘, 平均長寬均為 6—8 μm, 毛細胞數10—15(圖5, 表1, 表2)。23 dpf 前體管道神經丘表面出現分區, 以毛細胞組成的感受帶分布于神經丘表面中心區域, 以支持細胞與套細胞頂端構成的光滑表面環繞感受帶, 延伸至神經丘外緣(圖 5),23 dpf 前體管道神經丘成橢圓型, 長 15—19 μm, 長寬比 1.3—1.4, 神經丘及感受帶長、寬均出現顯著增長(P<0.05)毛細胞數增至 19—33, 之后前體管道神經丘繼續生長, 此時神經丘、感受帶大小顯著增大(P<0.05), 毛細胞數顯著增長(P<0.05)(表 1, 表2);37 dpf 之后前體管道神經丘建立基本完成, 神經丘、感受帶大小和毛細胞數已無明顯變化(P>0.05), 至51 dpf 軀干側線管道開始封閉之初, 前體管道神經丘呈長寬比為 1.6 的橢圓形, 長 29—33 μm, 毛細胞數增至30—45(表1, 表2)。16—30 dpf 表面神經丘生長明顯, 神經丘、感受帶大小顯著增大(P<0.05), 毛細胞數顯著增長(P<0.05), 30 dpf 后發育基本完成, 神經丘、感受帶大小和毛細胞數已無明顯變化(P>0.05)(表1, 表 2), 表面神經丘在發育全過程中均呈圓形, 長寬比為 0.9—1.1(圖5)。

圖1 鱖仔稚魚期軀干側線系統發育Fig.1 Post-embryonic development of the trunk lateral line system from larvae to juvenile
管道神經丘建立過程中, 長軸始終垂直于軀干,同時感受帶長軸與神經丘長軸平行, 毛細胞敏感方向始終平行于軀干側線管道方向(圖 5)。表面神經丘毛細胞敏感方向在軀干橫軸(前部?后部)與縱軸(背側?腹側)均具有, 并交替排列在側線管道周邊, 尾鰭上表面神經丘毛細胞敏感方向與軀干相似(圖5)。
3 討論
3.1 鱖后部側線系統主要發育過程與特點
鱖后部側線系統胚后發育在6—10 dpf 期間軀干已出現前體管道神經丘(L1—L7), 12 dpf 以后后續新生神經丘以頭部→尾部方向填于 L1—L7前體管道神經丘之間形成一行神經丘R0, 23—37 dpf 期間R0上方由頭部→尾柄方向形成一行神經丘(R1), 44 dpf 之后R0神經丘以出芽的方式在上下方向分化出附屬神經丘。整個 R0神經丘在 50—55 dpf 時期開始下沉, 軀干側線管道開始建立, 58 dpf 軀干側線管道建立完成,而之前來自R0兩行附屬神經丘與神經丘R1均成為表面神經丘。軀干管道神經丘在5—40 dpf 結構逐漸完善, 數量持續上升, 至37 dpf 基本發育完成, 在40—45 dpf 所有前體管道神經丘布置完成, 后續發育過程中單側數量穩定在 120—140; 表面神經丘自出現開始數量持續上升, 至58 dpf 后部側線系統單側表面神經丘數量已達到 380—450(圖 6)。由此可知, 鱖后部側線系統雖然在仔魚初期已開始發育, 但需要長達兩個月的時間完成主要發育(圖7)。鱖稚魚后期至幼魚階段生活、捕食習性發生顯著變化, 由仔魚時期的趨光性主動攝食逐漸轉為幼魚及成魚階段底棲伏擊型捕食, 而水體底層暗光、無光條件下側線感覺對其捕食行為較為重要(魏開建等, 1996; 田文斐等, 2012; Zhang et al, 2019),本研究中鱖仔魚→幼魚階段后部側線系統發育逐漸完善, 整體發育過程與鱸形目大部分魚類, 如麗魚科(Cichlidae)[如眼斑星麗魚(Astronotus ocellatusl), 奧利亞羅非魚(Oreochromis aureus), 九間波羅(Cichlasoma nigrofasciatum)](Webb, 1989b, 2013), 鱸 科(Percoidea)[如梅花鱸(Acerina cernua)](Sato et al, 2017),狼 鱸 科 (Moronidae)[ 如 七 星 鱸 (Lateolabrax japonicus)](Sato et al, 2017), 天竺鯛科(Apogonidae)[如僻梭竺鯛(Cercamia eremia)、稻氏天竺鯛(Ostorhinchus doederleini)]等(Sato et al, 2017, 2019), 但鱖在稚魚后期到幼魚時期(23→58dpf)后部側線系統神經丘結構變化、數量增長均較為明顯, 同時側線管道也在這一時期建立完成, 其后部側線系統的發育為鱖本階段底棲習性的逐漸形成提供了結構基礎。

圖3 軀干側線系統建立的組織學分析Fig.3 Histological analysis of trunk lateral line canal establishment
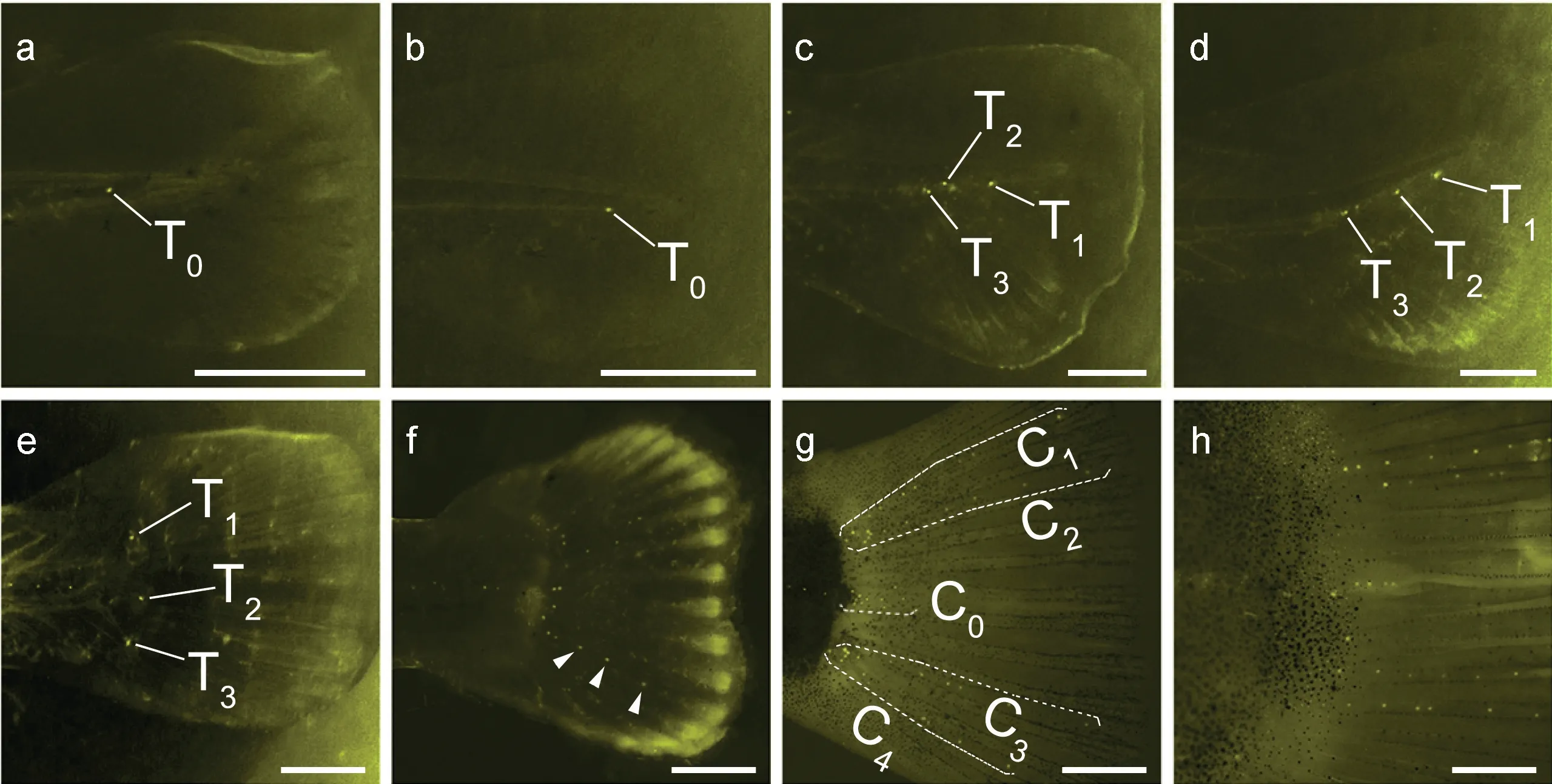
圖4 尾鰭側線系統發育Fig.4 Post-embryonic development of caudal fin lateral line

表1 神經丘形態數據統計分析Tab.1 Statistical analysis of neuromast morphological data

表2 神經丘感受帶形態數據統計分析Tab.2 Statistical analysis of neuromast sensory strip morphological data

圖6 不同時期管道神經丘與表面神經丘的數量特點Fig.6 Quantitative characteristics of CN and SN at different stages

圖7 鱖后部側線系統結構特點Fig.7 Structural characteristics of the posterior lateral system of mandarinfish
3.2 神經丘的結構與排布
硬骨魚類常通過增加特定感受方向敏感的神經丘數量來增強側線系統某一方向上的感受能力, 如斑馬魚(Danio rerio), 表面神經丘垂直分布, 在軀干上形成了一道道垂直于體軸的拉鏈狀的短列, 又如金魚(Carassius auratus)大量毛細胞敏感方向垂直于體軸的表面神經丘分布于軀干表面, 是對垂直方向水體信息感知的結構性適應(Ledent, 2002; Schmitz et al, 2008; Webb, 2013), 同時, 某些魚類神經丘結構出現特化, 如洞穴魚(Astyanax mexicanus)神經丘內含大量毛細胞, 明顯增大了神經丘感受帶的面積, 而神經丘與水體接觸的膠質頂大小直接由感受帶大小決定,這使得其具有極為靈敏的流體信息感知能力(Windsor et al, 2009; Yoshizawa et al, 2014)。鱸形目魚類后部側線系統常具有大量表面神經丘, 并成行或成列排布體表, 如鱸科(Percoidea)[如梅花鱸(Acerina cernua)](Sato et al, 2017), 狼鱸科(Moronidae)[如七星鱸(Lateolabrax japonicus)](Sato et al, 2017), 天竺鯛科(Apogonidae)[如僻梭竺鯛(Cercamia eremia)、稻氏天竺鯛(Ostorhinchus doederleini)]等(Sato et al, 2017,2019), 而鱖后部側線神經丘分布數量較少, 未見二類神經丘結構出現特化; 表面神經丘未見拉鏈狀排列, 僅分布于側線管道周邊, 這點與鱸形目其他魚類顯著不同(圖 7); 鱖尾鰭上表面神經丘數量也較少,但軀干上由水平、垂直方向敏感的表面神經丘交替排列, 使鱖后部側線系統具備了對水平、垂直方向流體信息感知, 同時, 表面神經丘集中分布于軀干背側,使鱖對上方環境的感知較為敏感。
3.3 軀干側線管道結構特點
硬骨魚類軀干側線管道結構多變, 隨著棲息環境的不同, 在結構上常出現顯著變化(Webb, 2013)。表層魚類常在軀干腹側形成一條平行于體軸的側線管道, 如斑馬魚(Danio rerio)、蝴蝶魚(Pantodon buchholzi)、飛魚(Exocoetidae)等在軀干腹側均有一條或幾條側線管道, 這種結構分布與下部流體信息的感知關系密切(Ledent, 2002; Schmitz et al, 2008;Webb, 2013), 底棲魚類( 如鮟鱇科、膽星魚科)軀干側線管道分布于背側, 特定增強了后部側線系統對上側流體信息的感知(Marshall, 1996)。參考硬骨魚類主要軀干側線系統類型, 鱖軀干側線管道系統中屬于完整弓形結構(圖7), 屬于鱸形目(Perciformes)魚類常見類型, 但背側方向的布置形式表明了鱖對其上側水體中捕食目標的感知能力較強。
4 結論
鱖后部側線系統的胚后發育始于 4—6 dpf 初期仔魚階段, 經歷長達兩月完成主要發育, 而稚魚后期到幼魚期(23→58 dpf)后部側線系統神經丘結構變化、數量增長均較為明顯, 同時側線管道也在這一時期建立完成, 這種結構變化與本階段鱖逐漸底棲的習性相適應; 同時, 鱖后部側線系統中, 側線管道的背側布置、神經丘結構與背側分布均增強了其上層水體的感知能力, 上述結構特征與鱖底棲伏擊型的捕食習性相適應。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
制造技術與機床(2019年10期)2019-10-26 02:47:06
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
鐵道通信信號(2018年5期)2018-06-28 03:06:24
家庭影院技術(2017年9期)2017-09-26 03:41:45
知識經濟·中國直銷(2017年5期)2017-06-15 20:28:19
通信電源技術(2016年6期)2016-04-20 06:21:32
