厚殼貽貝(Mytilus coruscus)養殖海域與天然生長海域的微生物群落比較研究*
2021-02-03 01:44:42李斯遠何治江呂泓玥湯秋晗智王健鑫嚴小軍張曉林
海洋與湖沼 2021年1期
關鍵詞:生長
李斯遠 何治江 呂泓玥 湯秋晗 廖 智王健鑫 嚴小軍, 張曉林,
(1. 浙江海洋大學海洋科學與技術學院 舟山 316022; 2. 浙江海洋大學 國家海洋設施養殖工程技術研究中心舟山 316022)
厚殼貽貝Mytilus coruscus 屬軟體動物門Mollusca、雙殼綱 Bivalvia、異柱目 Anisomyaria、貽貝科Mytilidae、貽貝屬Mytilus(葉瑩瑩等, 2011), 俗稱淡菜,為溫水性種, 主要分布在我國黃海、渤海和東海沿岸,以浙江沿海資源量最大。厚殼貽貝具有肉質鮮美、營養豐富、蛋白質含量高、生長繁殖快、抗病能力強、易于人工養殖等諸多優點, 是浙江舟山海域具有重要經濟價值的養殖貝類之一(徐晴晴等, 2018)。目前厚殼貽貝主要養殖區域為浙江嵊泗海域和東極海域。厚殼貽貝在養殖過程中, 對于海域環境中的藻類和微生物多樣性具有較強的依賴性; 一方面, 藻類是貽貝的主要食物來源(朱雨瑞等, 2010), 另一方面, 貽貝在養殖過程中對海域微生物群落具有特異的富集作用(陳瑜等, 2016), 但目前尚不清楚微生物群落對貽貝的養殖具有哪些影響。
但近年來, 隨著厚殼貽貝的養殖區域的不斷擴展和養殖密度逐漸增加, 其養殖過程顯現種種問題,例如貽貝個體的肥滿度下降, 生長速度下降, 貽貝逃苗和病害也時有發生(林永添, 2007)。有研究顯示, 微生物群落變化能夠影響貽貝的附著和防病(楊娜,2018; 梁簫等, 2020)。國內外已有針對不同貝類生長區海域浮游群落的調查研究(陳雷等, 2013; 尹潔慧,2013; 金雷等, 2015; Fogel et al, 2015), 但其主要焦點集中于海域中浮游植物的種類和豐度對貝類攝食以及營養價值的影響。關于海域微生物群落與貽貝生長過程以及生理生化的影響尚未見報道。厚殼貽貝作為我國東部海域最重要的養殖貝類之一, 其生存環境的微生物群落調查以及與其體內微生物群落的對比分析將對貽貝養殖具有重要研究意義。為此, 本文將針對厚殼貽貝生長海域的微生物群落結構特征及其體內微生物組成開展研究, 以期為厚殼貽貝的健康、生態養殖提供重要的理論依據。
本研究主要以厚殼貽貝為對象, 首先將調查嵊泗縣枸杞島和東極鎮廟子湖島厚殼貽貝養殖區和野生區海域的微生物群落的結構特征, 并與厚殼貽貝體內分離到的微生物組成情況相結合進行比較分析,以探明厚殼貽貝生長海域的微生物群落分布特征以及養殖和野生生長海域的微生物組成差異。本論文的研究結果一方面能夠為厚殼貽貝的人工養殖和野生馴化的海域選擇提供強有力的參考, 另一方面可以維持海域微生物群落的動態平衡, 促進貝類養殖產業的健康、可持續發展。
1 材料與方法
1.1 厚殼貽貝生存海域的海水微生物群落分析
1.1.1 實驗材料 于2019 年5—6 月, 選擇嵊泗縣枸杞島(30°42′N, 122°46′E)厚殼貽貝養殖區海域 5、10 m 水深和厚殼貽貝野生生長區海域5、10 m 水深;東極鎮廟子湖島(30°11′N, 122°41′E)厚殼貽貝養殖海域5、10 m 水深和厚殼貽貝野生生長區海域5、10 m水深, 共計 8 個點位, 具體采樣位置見圖 1。在調查期間, 每個點位采集3 個平行海水樣本, 送往實驗室經靜置沉淀并過濾后進行16S rRNA 擴增子測序。

圖1 枸杞島(右下)和東極島(右上)厚殼貽貝養殖和野生生長海域采樣站位圖Fig.1 Location of the sampling points for M. coruscus in natural and cultural regions in Gouqi Island (lower right) and Dongji Island(upper right)
1.1.2 實驗方法 取質量合格的基因組DNA 樣品30 ng 及對應的融合引物配置 PCR 反應體系, 設置PCR 反應參數進行 PCR 擴增, 使用 Agencourt AMPure XP 磁珠對PCR 擴增產物進行純化, 并溶于Elution Buffer, 貼上標簽, 完成建庫。 使用Agilent2100 Bioanalyzer 對文庫的片段范圍及濃度進行檢測。檢測合格的文庫根據插入片段大小, 選擇HiSeq 平臺進行測序。然后對所有樣品的全部序列進行聚類, 以97%的相似度將序列聚類成OUT, 然后對OTU 的代表序列進行物種注釋, 確定16S rRNA 序列對應的微生物名稱(趙立君等, 2019)。
1.1.3 生物信息學分析 下機數據過濾, 剩余高質量的Clean data 用于后期分析(Magoc et al, 2011);通過 reads 之間的 overlap 關系將 reads 拼接成 Tags;將Tags 聚類成OTU 并與數據庫比對、物種注釋; 基于 OTU 和注釋結果進行樣品物種復雜度分析, 組間物種差異分析等。再利用 UCHIME(v 4.2.40)將 PCR擴增產生的嵌合體從 OTU 代表序列中去除, 使用usearch_global 方法將所有 Tags 比對回 OTU 代表序列(Wang et al, 2007), 對每個樣品的OTU 豐度、多樣性指數等進行分析, 同時對物種注釋在門分類水平上進行群落結構的統計分析。在以上分析的基礎上,進行基于OTU 的主成分分析(PCA)。
1.2 厚殼貽貝體內組織微生物分離與鑒定
1.2.1 培養基制備 第一類分離選用 2216E 普通海水培養基。為還原貽貝組織的營養成分, 盡可能分離到貽貝體內全部種類的微生物, 選取足量的厚殼貽貝全組織進行勻漿并過濾, 120°C、21 min 高溫高壓處理, 加入適量瓊脂制備成厚殼貽貝肉湯培養基, 用于第二類分離。
1.2.2 微生物分離與篩選 選取來自同一生長環境的厚殼貽貝 10 只, 解剖取其鰓、性腺、消化腺、外套膜、足并用無菌海水輕輕沖洗去除組織表面雜質,使用兩種不同方式分離微生物: (1) 第一類分離針對貽貝組織表面附著的微生物, 用無菌棉簽分別刮取不同組織的表面, 將棉簽伸至于放有玻璃珠的三角瓶中并加入少量無菌海水, 常溫振蕩培養1 h。(2) 第二類次分離針對可能在貽貝體內生長的微生物, 分別將上述5 類組織勻漿后, 加入放有玻璃珠的三角瓶中, 常溫振蕩培養 1 h。兩次分離都根據選用的不同組織分成5 組, 每組3 次重復。
取上述適量處理過的培養液接種到對應的培養基中, 28°C 過夜培養, 分別挑取不同細胞形態、大小、顏色的菌落至海水LB 培養基上, 反復純化三次。
1.2.3 微生物鑒定 使用試劑盒(TaKaRa MiniBEST Bacteria Genomic DNA Extraction Kit)分別提取篩選菌落的基因組 DNA, 選用通用引物27F (5′-AGAGTTTGATCCTGGCTCAG-3′)和 1492R(5′-TACGGTTACCTTGTTACGACTT-3′)PCR 擴增 16S rDNA, PCR 產物送至生工生物工程(上海)有限公司進行進一步的分析和鑒定。
2 結果與分析
2.1 海水樣品測序結果
如表1、表2 所示, 兩個海域的所有樣品優化后序列占優化前序列的 80%以上, 因此可以很好地用于后續的進一步分析。
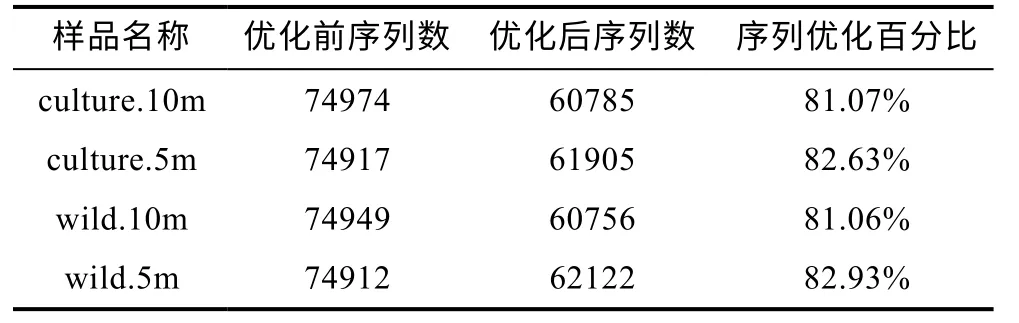
表1 東極海域微生物群落16S rRNA 擴增子測序序列結果優化對比Tab.1 Comparison of microbes 16S rRNA sequencing in the Dongji sea area
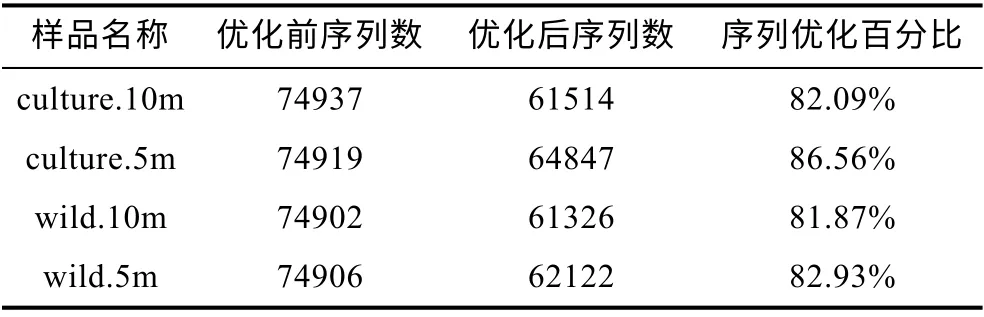
表2 嵊泗海域微生物群落16S rRNA 擴增子測序序列結果優化對比Tab.2 Comparison of microbes 16S rRNA sequencing in the Shengsi sea area
2.2 各海域微生物群落多樣性特征
通過多樣性指數可以對兩個海域海水樣品中微生物多樣性進行分析。從表 3 可得, 東極海域海水樣品有效序列共計245568 條, 其中養殖區海域10 m 水深樣品包含 60785 條, OTU 數 674 個; 養殖區海域 5 m 水深樣品包含61905 條, OTU 數616 個; 野生生長區海域10 m 水深樣品包含 60756 條, OTU 數 635 個; 野生生長區海域 5 m 水深樣品包含62122 條, OTU 數 535 個。
從表 4 可得, 嵊泗海域海水樣品有效序列共計249809 條, 其中養殖區海域 10 m 水深樣品包含61514 條, OTU 數 592 個; 養殖區海域 5 m 水深樣品包含64847 條, OTU 數584 個; 野生生長區海域10 m水深樣品包含61326 條, OTU 數623 個; 野生生長區海域5 m 水深樣品包含62122 條, OTU 數570 個。
綜上可以得, 東極海域養殖區微生物(OTU 為616—674)豐富度大于野生生長區微生物(OTU 為535—635), 10 m 水深微生物(OTU 為 635—674)豐富度大于5 m 水深微生物(OTU 為535—616); 嵊泗海域野生生長區區微生物(OTU 為 535—674)豐富度大于養殖區區微生物(OTU 為584—592), 10 m 水深微生物(OTU 為592—623)豐富度大于5 m 水深微生物(OTU為570—584)。嵊泗海域總體微生物(OTU 為570—623)豐富度大于東極海域(OTU 為535—674)。
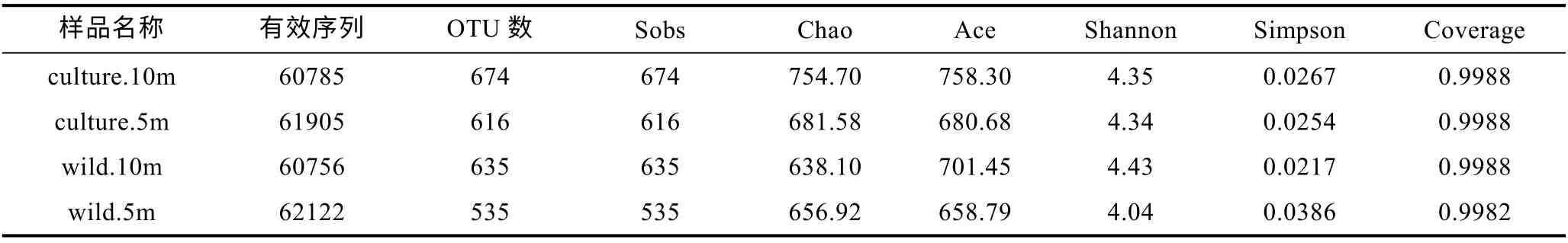
表3 東極海域樣品OTU 數及多樣性指數Tab.3 The OTU and diversity index in the Dongji sea area
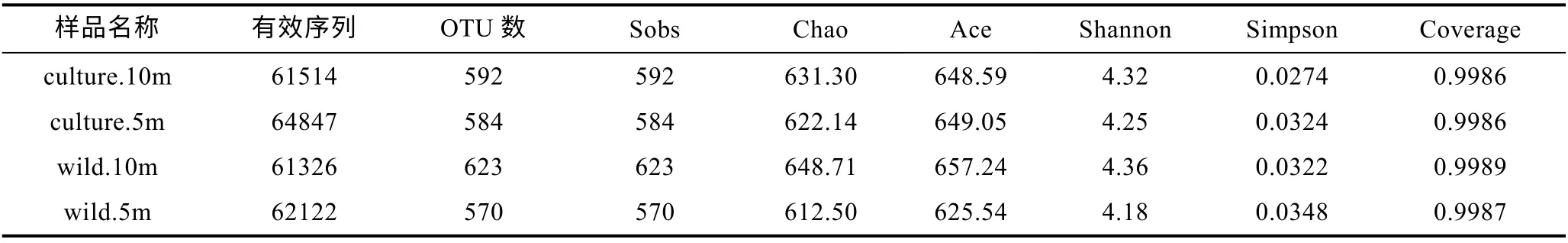
表4 嵊泗海域樣品OTU 數及多樣性指數Tab.4 The OTU and diversity index in the Shengsi sea area
本文通過包括Sobs 指數、Chao 指數、ACE 指數、Shannon 指數、Simpson 指數以及 Good- coverage 指數的 Alpha 多樣性來分析海水樣品中微生物多樣性差異(Schloss et al, 2009)。Sobs 指數、Chao 指數和ACE 指數反映樣品中群落的豐富度, 而Shannon 指數以及Simpson 指數反映群落的多樣性, Good-coverage反映了樣品文庫的覆蓋率。其中, Sobs 指數、Chao指數和ACE 指數和Shannon 指數越大, Simpson 指數越小, 說明樣品中的物種越豐富; Good-coverage 數值越高, 則表明樣品中序列沒有被測出來的概率越低,該值反映了測序結果是否代表樣品的真實情況, 所有樣品的 Good-coverage 指數均在 0.998 以上, 因此本次測序結果代表了樣本中微生物的真實情況。
由此可得, 東極海域不同采樣點微生物豐富度及多樣性由大到小分別為 culture.10m > wild.10m >culture.5m > wild.5m; 嵊泗海域不同采樣點微生物豐富度及多樣性由大到小分別為wild.10m > culture.10m >culture.5m > wild.5m。
Core-Pan OTU 以花瓣圖展示所有樣本共有和特有的 OTU, 分析數據與維恩圖相同, 但相比于維恩圖只展示 5 組(或 5 個樣品)以內的數據, 花瓣圖可以展示任意組(或樣品), 因此可以盡可能多的展示組間OTU 的異同。
從圖2 可知, 東極海域樣品微生物OTU 數為440—473, 共有 OTU 達到 436, 說明東極海域總體上微生物組成相似度較高, 養殖區海域 10 m 水深特有OTU 最高, 達到37; 野生生長區5 m 水深特有OTU最低, 僅達到 4。從圖 3 可知, 嵊泗海域樣品微生物OTU 數為 455—473, 共有 OTU 達到 439, 說明嵊泗海域總體上微生物組成相似度也較高, 野生生長區海域10m 水深特有OTU 最高, 達到34; 養殖區海域特有OTU 均最低, 僅達到16。
2.3 各海域微生物群落門分類水平組成分析
物種柱狀圖可以直觀地展示各樣本物種組成及比例, 反映樣本間物種的變化情況。 通過與數據庫進行比對, 對 OTU 進行物種分類, 本文選取在門分類水平上(物種豐富度均大于 0.5%)對各樣品所作的物種豐度柱狀圖。
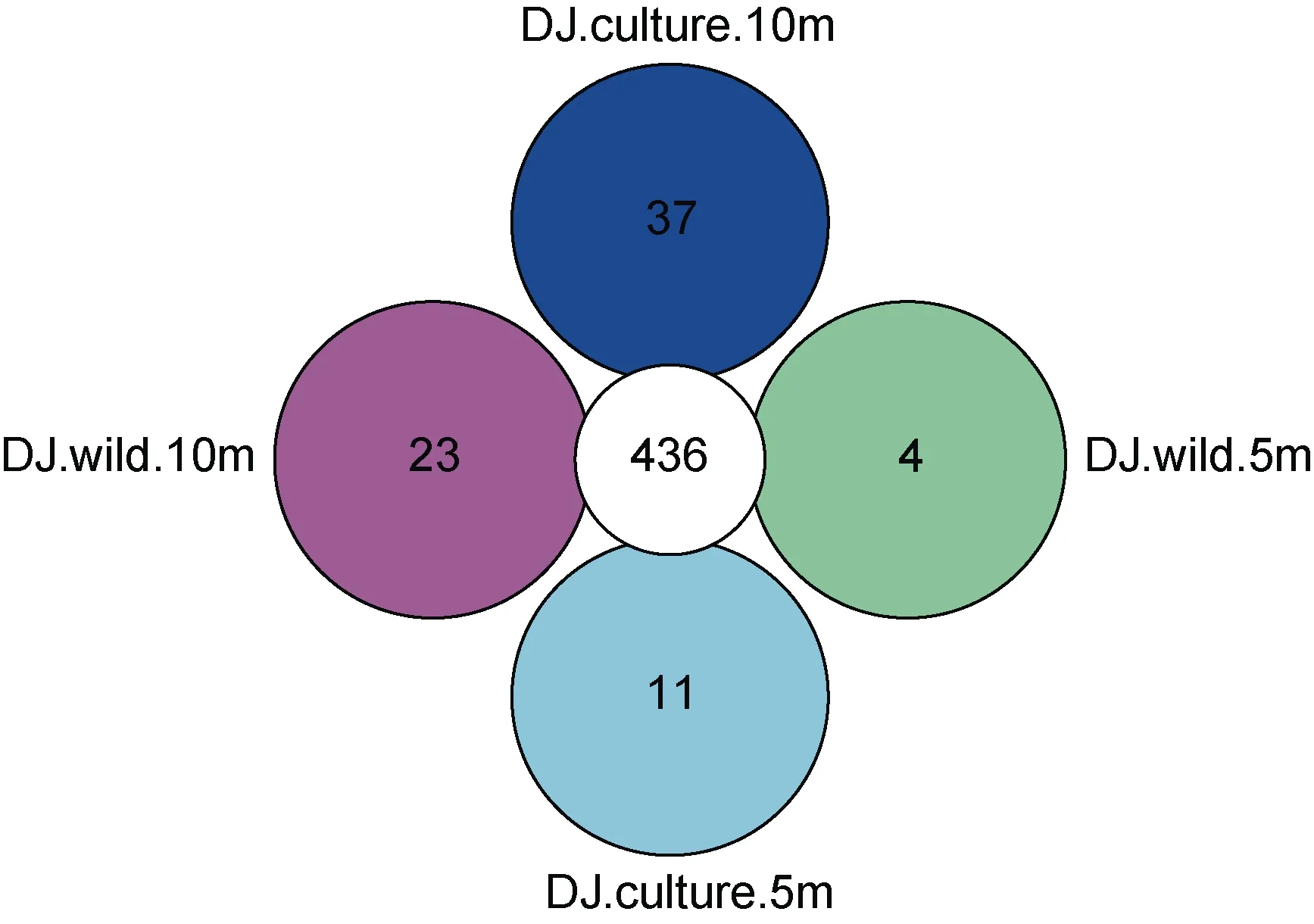
圖2 東極海域樣品OTU 分布Core-Pan 圖Fig.2 The Core-pan diagram of OTU distribution in the Dongji sea area
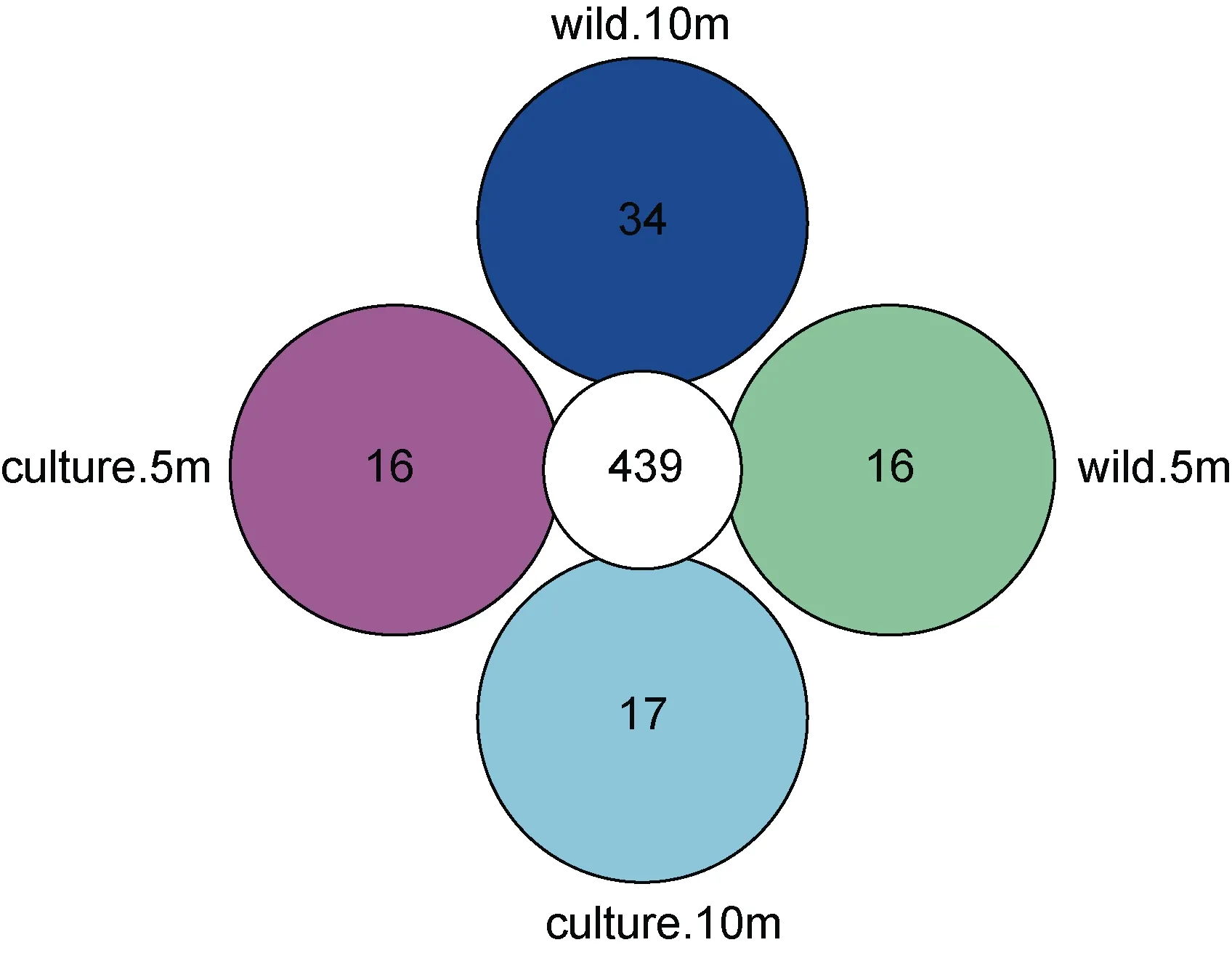
圖3 嵊泗海域樣品OTU 分布Core-Pan 圖Fig.3 The Core-pan diagram of OTU distribution in the Shengsi sea area
從圖 4 可知, 東極海域主要包含變形菌門Proteobacteria、擬桿菌門 Bacteroidetes、廣古菌門Euryarchaeota、放線菌門 Actinobacteria、泉古菌門Crenarchaeota 等在內的 30 個門類, 其中變形菌門是四個采樣點的最大優勢門類, 其在野生生長區 10 m水深、野生生長區5 m 水深、養殖區10 m 水深、養殖區 5 m 水深的百分比分別為 37.58%、28.53%、36.26%、37.13%, 擬桿菌門(19.72%—26.41%)、放線菌門(9.23%—20.86%)是東極海域第二、第三優勢細菌門, 這三大優勢細菌門類占據東極海域細菌比例的60%以上。
此外, 東極養殖區海域和野生生長區海域微生物門類無明顯差異, 但可明顯得知野生生長區海域各類優勢細菌門所占比例大于養殖區海域各類優勢細菌門所占比例。
從圖 4 可知, 東極海域主要包含變形菌門Proteobacteria、擬桿菌門 Bacteroidetes、廣古菌門Euryarchaeota、放線菌門 Actinobacteria、泉古菌門Crenarchaeota 等在內的27 個門類, 其中變形菌門是四個采樣點的最大優勢門類, 其在野生生長區10 m 水深, 野生生長區5 m 水深, 養殖區10 m 水深, 養殖區5 m 水深的百分比分別為51.48%、54.66%、61.88%、60.46%, 擬桿菌門(17.48%—19.72%), 放線菌門(9.08%—11.99%)是嵊泗海域第二、第三優勢細菌門, 這三大優勢細菌門類占據東極海域細菌比例的80%以上。
同樣, 嵊泗養殖區海域和野生生長區海域微生物門類無明顯差異, 但可明顯得知野生生長區海域各類優勢細菌門所占比例大于養殖區海域各類優勢細菌門所占比例。
綜合圖4、圖5 可知, 東極海域總體微生物豐富度要大于嵊泗海域, 但是在同樣優勢菌門的條件下,嵊泗海域各類優勢菌門所占的微生物總比例遠高于東極海域的各類優勢菌門。
2.4 各海域微生物群落屬分類水平組成分析
從圖6 可知, 東極海域主要包含厭氧氨氧化菌屬Candidatus Portiera (5.57%—8.03%), 海洋氨氧化古菌 屬 Nitrosopumilus (5.27%—6.94%), Fluviicola(4.21%—6.34%), 藍細菌聚球藻屬 Synechococcus(1.82%—3.09%), 弧菌屬Vibrio (0.33%—2.50%)等在內的9 個屬類。從表5 可知, 東極海域厚殼貽貝養殖區和野生生長區屬分類水平微生物豐度之間的比較無明顯規律。
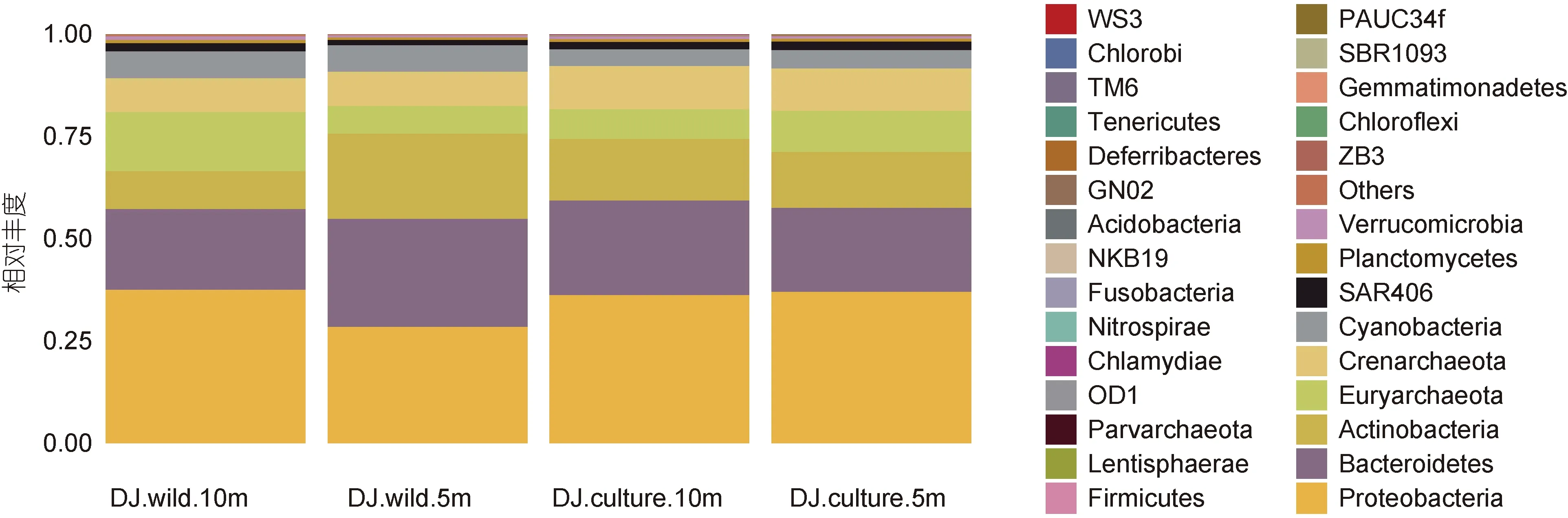
圖4 東極海域微生物門分類水平物種豐度柱狀圖Fig.4 The phylum abundance in the Dongji sea area
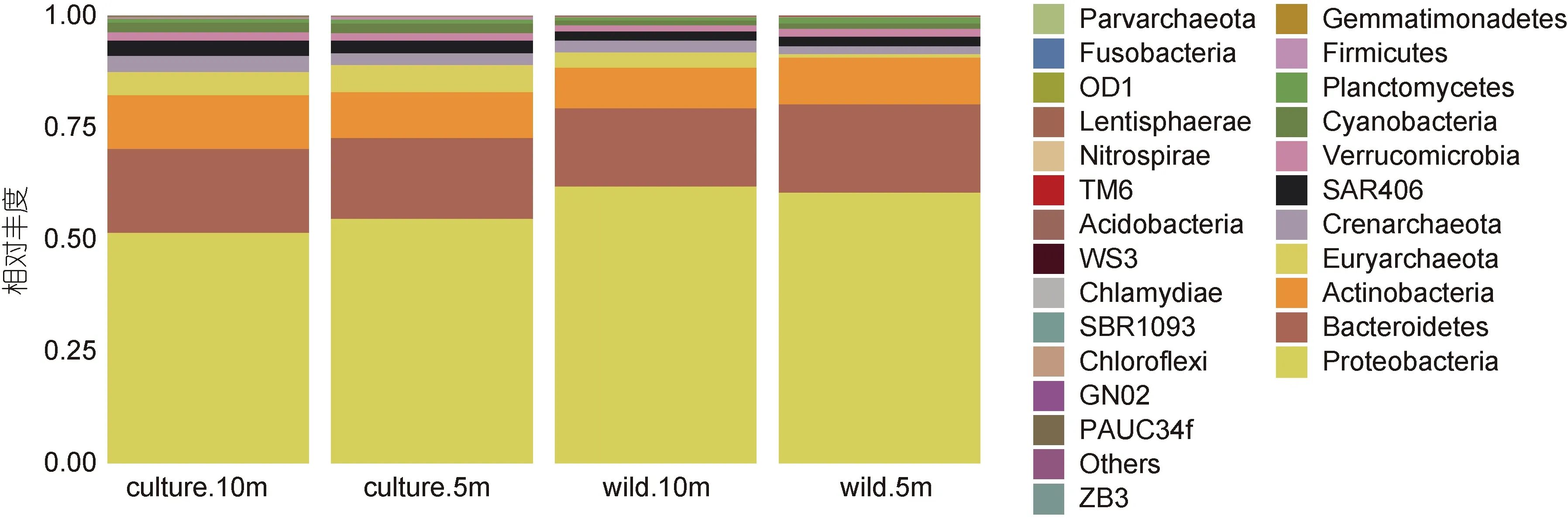
圖5 嵊泗海域微生物門分類水平物種豐度柱狀圖Fig.5 The phylum abundance in the Shengsi sea area
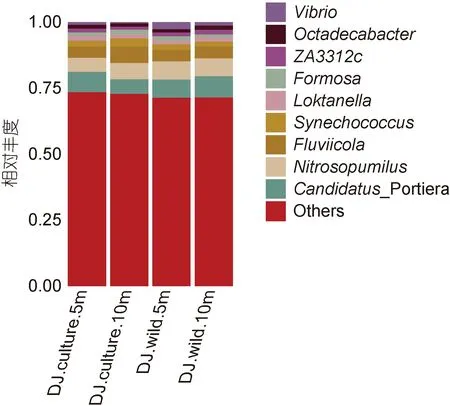
圖6 東極海域微生物屬分類水平物種豐度柱狀圖Fig.6 The genus abundance in the Dongji sea area
從圖 7 可知, 嵊泗海域主要包含十八桿菌屬Octadecabacter (10.80%—15.04%), 假交替單胞菌屬Pseudoalteromonas (2.41%—8.77%), 厭氧氨氧化菌屬 Candidatus_Portiera (3.57%—6.69%), ZA3312c(1.80%—3.21%), 沉積物陸丹氏菌屬 Loktanella(3.26%—4.29%), 弧菌屬Vibrio (0.21%—6.36%)等在內的13 個屬類。
從表6 可知, 嵊泗海域厚殼貽貝養殖區和野生生長區屬分類水平微生物豐度之間可觀察到明顯的規律, 總體上養殖區海域各類優勢屬微生物豐度大于野生生長區海域。
2.5 各海域微生物群落相似度分析
本文采用主成分(PCA)分析, 通過分析不同樣品在圖上的分散和聚集的情況, 根據離散程度可以了解不同樣品的組成是否具有相似性(Jolliffe, 1986;Avershina et al, 2013)。從圖 8 可知: 軸 1 (PC1)貢獻值為72.72%, 軸2 (PC2)貢獻值為13.13%, 總貢獻值為85.85%。從整體上來看, 東極和嵊泗海域之間的微生物群落存在明顯差異, 兩個海域的野生生長區5 m 和10 m 之間的微生物群落相似度明顯低于養殖區5 m和10 m 的微生物群落相似度, 而這種差異在嵊泗海域尤為明顯。
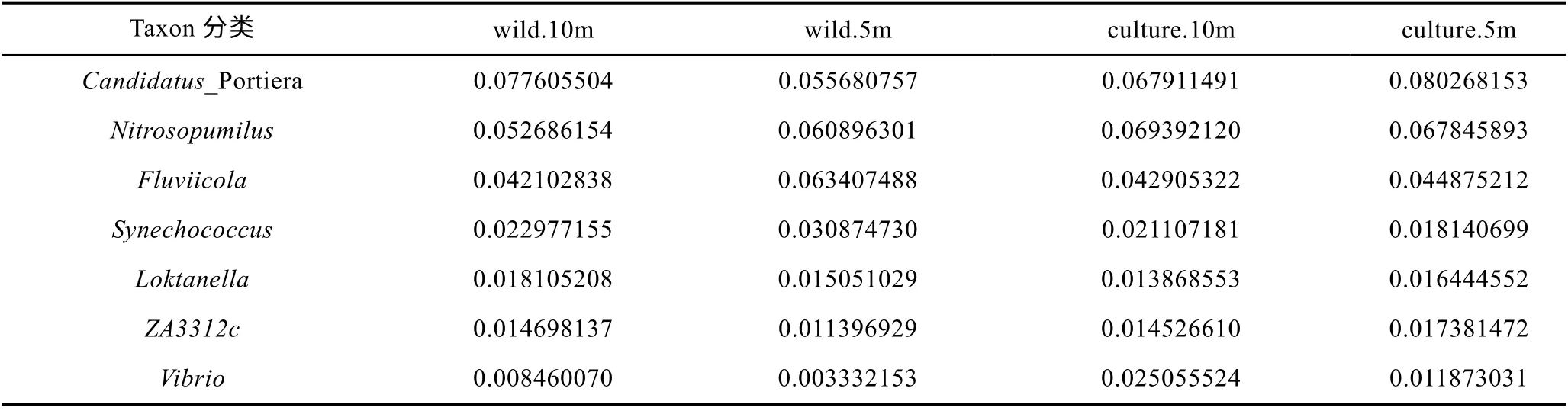
表5 東極海域微生物群落16S rRNA 擴增子測序屬分類水平微生物群落結構表Tab.5 The community structure in genus level in the Dongji sea area
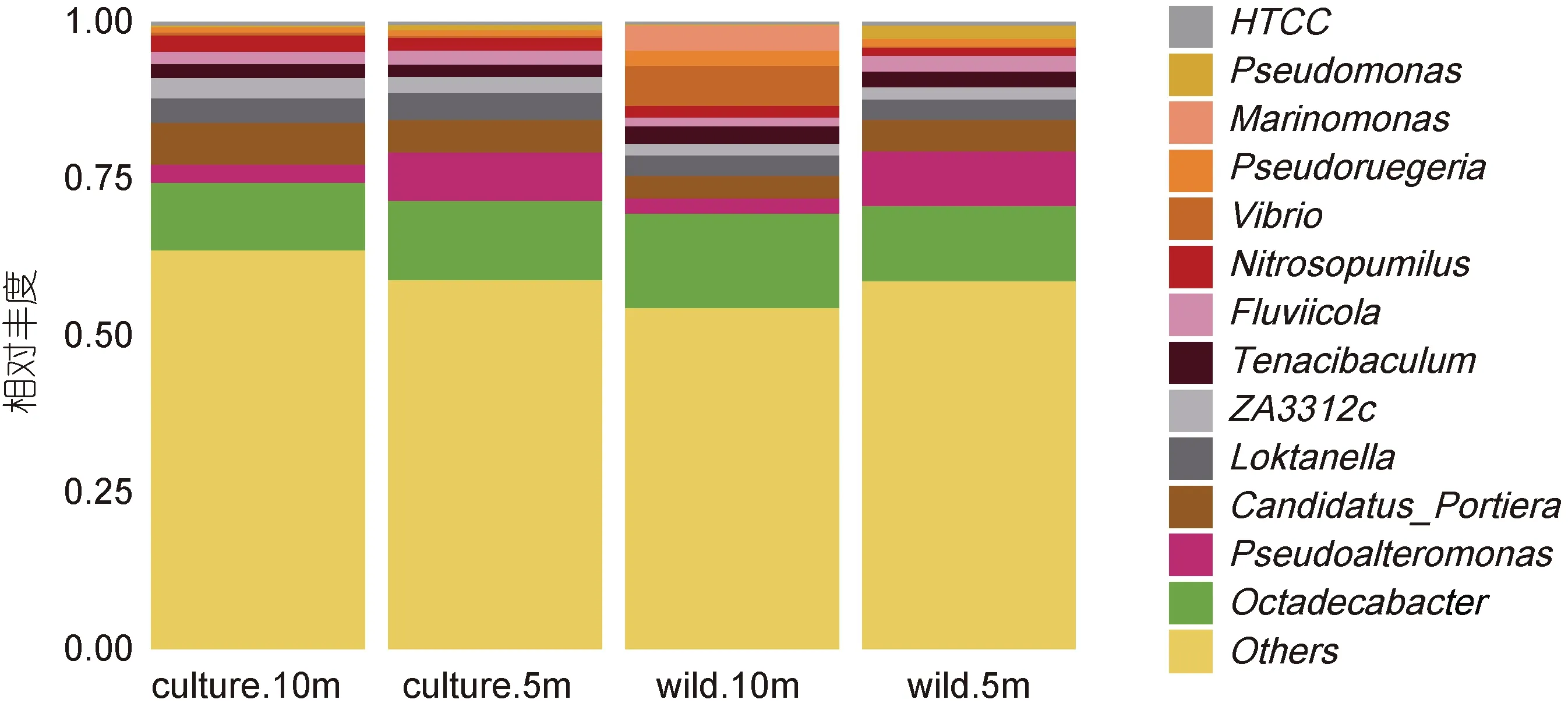
圖7 嵊泗海域微生物屬分類水平物種豐度柱狀圖Fig.7 The genus abundance in the Shengsi sea area
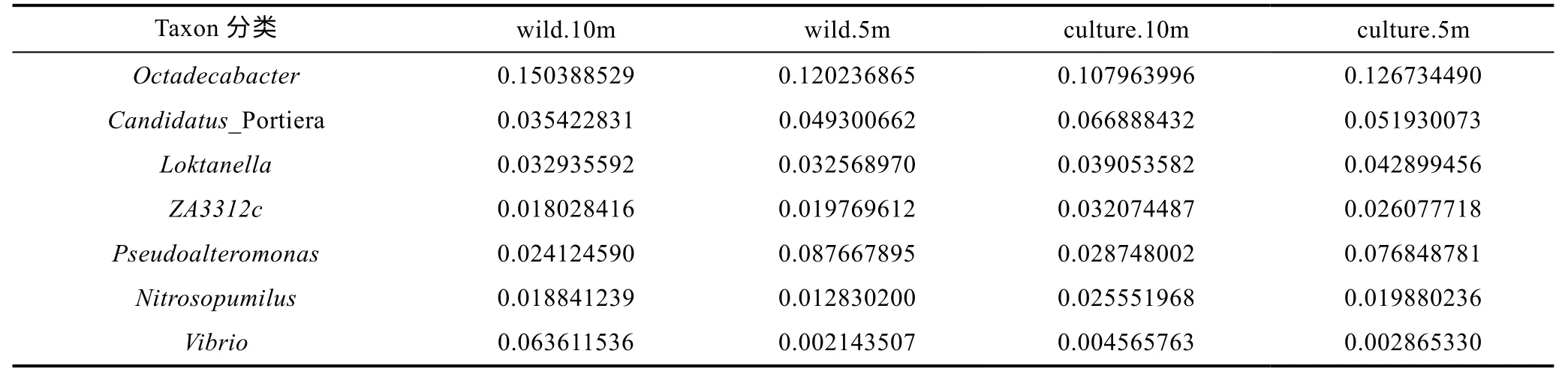
表6 嵊泗海域屬分類水平微生物群落結構表Tab.6 The community structure in genus level in the Shengsi sea area
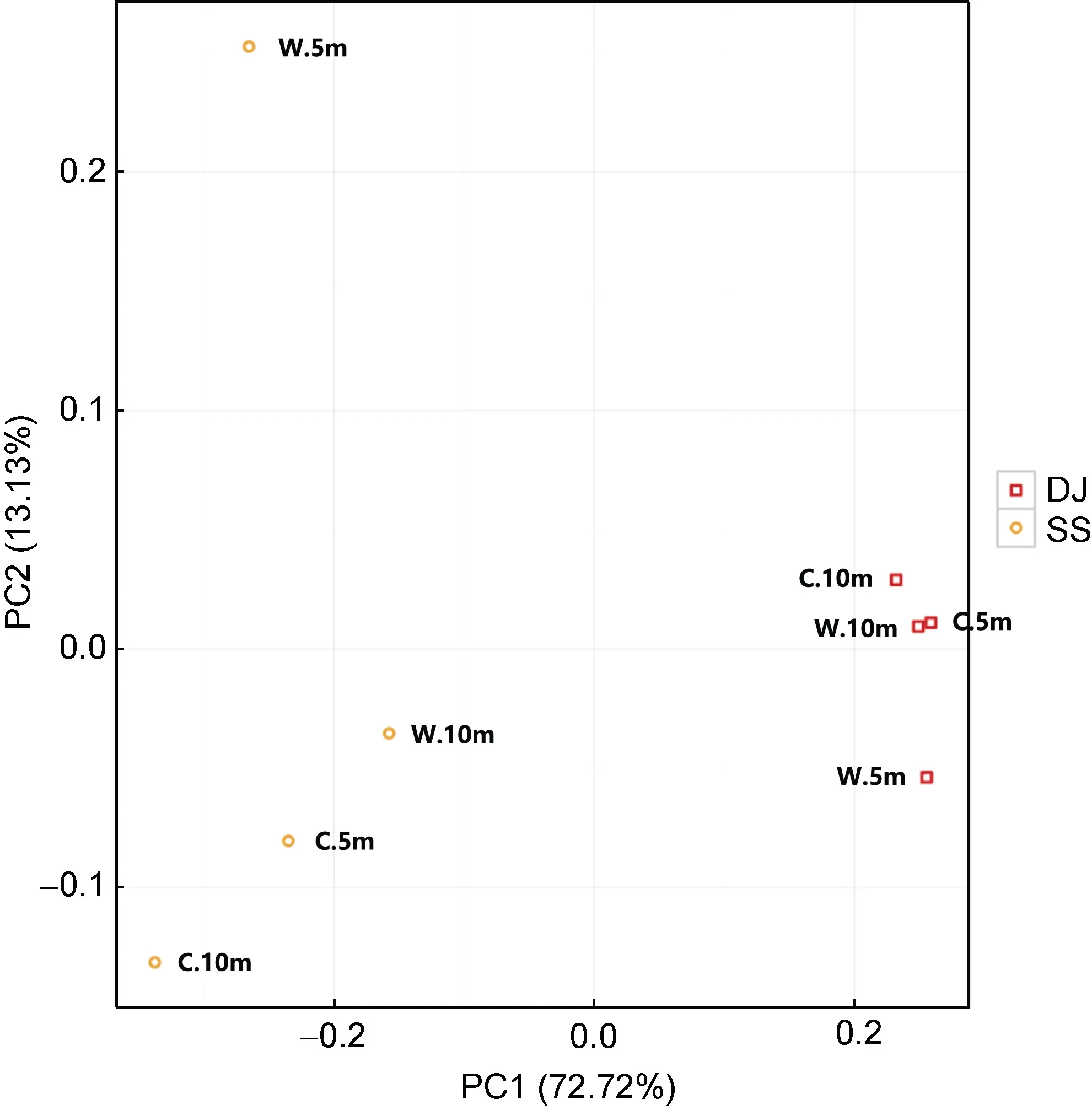
圖8 各海域樣品PCA 分析圖Fig.8 The PCA analysis diagram of sea area samples
2.6 厚殼貽貝體內分離微生物組成分析
由表7 可知, 從厚殼貽貝的鰓、性腺、足、消化腺中均分離出微生物, 主要包括: 黏著桿菌屬Tenacibaculum sp., 蠟樣芽孢桿菌屬 Bacillus cereus,紅色桿菌屬 Erythrobacter sp., 魯杰氏菌屬 Ruegeria sp., 弧菌屬Vibrio sp., 巨型弧菌屬Vibrio gigantis, 假交替單胞菌屬Pseudoalteromonas 這7 類菌屬。
結合上文圖7 可發現, 在厚殼貽貝生長海域水體中檢測分析得到的假交替單胞菌屬 Pseudoalteromonas,黏著桿菌屬 Tenacibaculum sp.以及弧菌屬 Vibrio sp.均在厚殼貽貝體內組織中檢測到, 且這三類菌屬都出現于厚殼貽貝的鰓組織中。
3 討論
3.1 厚殼貽貝養殖環境及其體內微生物群落多樣性分析
嵊泗海域總體微生物豐富度大于東極海域。嵊泗海域不同采樣點微生物豐富度由大到小分別為wild.10m > culture.10m > culture.5m > wild.5m, 其中嵊泗海域野生生長區 10 m 水深微生物豐富度最高,野生生長區5 m 水深微生物豐富度最低; 東極海域不同采樣點微生物豐富度由大到小分別為 culture.10m> wild.10m > culture.5m > wild.5m, 東極海域養殖區10 m 水深微生物豐富度最高, 野生生長區10 m 水深微生物豐富度最低。

表7 厚殼貽貝體內分離微生物組成表Tab.7 The microbial composition inside the Mytilus coruscus
從屬分類水平來看, 東極海域的優勢微生物群落主要為厭氧氨氧化菌屬 Candidatus_Portiera、海洋氨氧化古菌屬 Nitrosopumilus、Fluviicola、藍細菌聚球藻屬 Synechococcus 以及弧菌屬 Vibrio; 嵊泗海域的優勢微生物群落主要為十八桿菌屬 Octadecabacter、假交替單胞菌屬 Pseudoalteromonas, 厭氧氨氧化菌屬Candidatus_Portiera、ZA3312c、沉積物陸丹氏菌屬Loktanella 以及弧菌屬Vibrio。
總體上, 東極海域4 個取樣點的微生物群落相似度比嵊泗海域高。由于受到厚殼貽貝長期選擇性攝食的影響, 東極和嵊泗兩個海域的野生生長區 5m 和10m 之間的微生物群落相似度明顯低于養殖區5m 和10m 的微生物群落相似度, 且這種差異在嵊泗海域尤為明顯。這可能是受嵊泗近海海域厚殼貽貝的大面積養殖的影響。從已有的文獻中可知(王芳等, 1998; 劉緒博, 2011; 周曦杰, 2014), 包括貽貝在內的許多雙殼貝類有著選擇性濾食的習性, 即這些貝類會優先選擇攝食水體中的一類浮游生物, 因而導致該區域水體中浮游生物種類相對單一, 相似度較高。相比于嵊泗海域, 東極海域4 個取樣點的微生物群落相似度較高, 而東極近海海域厚殼貽貝的養殖規模遠不如嵊泗海域大, 因此近海微生物群落受厚殼貽貝的攝食影響較小, 和野生生長區微生物群落相比, 相似度就更高。
Vezzulli 等人對位于意大利利古里亞海的地中海貽貝養殖場的水體微生物群落進行了分析, 發現弧菌屬和假交替單胞菌屬是該海域的主要優勢微生物(Vezzulli et al, 2018), 這與本文嵊泗海域的主要優勢微生物結果相一致。而東極海域也發現假交替單胞菌這一種優勢微生物, 同時也間接印證了東極海域的微生物豐富度不如嵊泗海域。
在厚殼貽貝生長海域以及厚殼貽貝體內組織都鑒定到了假交替單胞菌屬Pseudoalteromonas、黏著桿菌屬Tenacibaculum sp.以及弧菌屬Vibrio sp.這三類微生物, 初步推測厚殼貽貝和這些自然水域中存在的微生物產生了某種共生關系, 有報道說海洋雙殼貝類會將某些環境中的細菌甚至是致病菌富集在鰓上,以此到達幫助宿主攝食消化、阻止其他病原體侵入的作用, 這也是貽貝不易染病的原因之一(Yu et al,2019)。相似的研究在紫貽貝中也有報道, 通過對土耳其伊茲密爾沿海生長的紫貽貝體內微生物進行宏基因組分析, 結果發現紫貽貝中也存在弧菌、變形桿菌、摩根氏菌等在內的34 種菌屬(Bozcal et al, 2020)。
3.2 貽貝及其共生微生物的獨特適應性
無論是厚殼貽貝生長海域, 還是其體內均分離鑒定到大量弧菌。而之前的報道顯示在紫貽貝體內也大量存在弧菌(Bozcal et al, 2020)。其中弧菌是水產養殖中常見的致病菌, 在紫貽貝和厚殼貽貝體內都存在大量的弧菌, 說明貽貝對弧菌具有特殊的適應性,其特殊的適應機制和病原識別模式還有待于進一步研究。目前關于弧菌和宿主的共生已有一些研究, 例如有報道顯示在海洋魚類中, 弧菌會在進入宿主腸道系統后, 通過其VI 型分泌系統分泌一些效應分子,刺激宿主腸道產生收縮使得宿主腸道內原有的常駐菌群被迫排出, 最終弧菌在宿主腸道微生物群落中占據主導地位, 然而在這一腸道微生物群落更替變化中似乎并不會對宿主的健康產生損害, 弧菌本身似乎也不受腸道運動增強的影響(Logan et al, 2018); 而Vezzulli 等人認為貽貝體內的弧菌與其宿主經歷了一段較為漫長的共同進化史(Vezzulli et al, 2018); Septer等人的研究表明, 夏威夷魷魚在孵化后的數個小時內就會被費氏弧菌所感染, 這種感染將逐漸演變成一種共生, 并且持續至宿主的一生, 而這種共生關系和夏威夷魷魚的發光器官有直接的聯系(Septer, 2019)。
4 結論
總的來說, 目前對于海洋雙殼貝類共生菌的報道尚比較匱乏, 其識別微生物的機理及共生機制還尚不完全明確, 本文從厚殼貽貝體內分離得到的微生物是否和厚殼貽貝發生了共生關系, 亦或是對厚殼貽貝的某些生理功能產生影響等問題, 還需進一步探索。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
