選擇性多聚腺苷酸化的生物學效應及其調控機制研究進展
2021-02-02 08:28:08徐海冬寧博林牟芳李輝王寧
遺傳 2021年1期
徐海冬,寧博林,牟芳,李輝,王寧
選擇性多聚腺苷酸化的生物學效應及其調控機制研究進展
徐海冬1,2,3,寧博林1,2,3,牟芳1,2,3,李輝1,2,3,王寧1,2,3
1. 農業農村部雞遺傳育種重點實驗室,哈爾濱 150030 2. 黑龍江省普通高等學校動物遺傳育種與繁殖重點實驗室,哈爾濱 150030 3. 東北農業大學動物科學技術學院,哈爾濱 150030
真核生物基因的前體mRNA (pre-mRNA)及一些lncRNA在成熟過程中其3'端會發生剪切和多聚腺苷酸化反應(cleavage and polyadenylation, C/P),C/P的發生需要多聚腺苷酸化信號(polyadenylation signal, PAS)的存在。選擇性多聚腺苷酸化(alternative cleavage and polyadenylation, APA)是指具有多個PAS的基因,在其mRNA 3?端成熟過程中,由于選擇不同的PAS,導致產生出多個3'UTR長度和序列組成不同的轉錄異構體。3?UTR長度和序列的不同會影響mRNA的穩定性、翻譯效率、運輸和細胞定位等,因此APA是真核生物的一個重要轉錄后調控方式。近年來,對大量動物、植物及酵母的基因組測序分析發現,APA在真核生物廣泛存在,針對APA的生物學效應和調控機制開展了一系列研究。目前已鑒定出許多APA調控的順式調控元件和反式作用因子。本文重點介紹了APA生物學效應和調控機制的最新研究進展,并探討了未來APA調控的研究方向。
基因表達調控;選擇性多聚腺苷酸化;多聚腺苷酸化信號;順式調控元件;反式作用因子
真核生物的mRNA前體(pre-RNA)和一些長鏈非編碼RNA前體(pre-lncRNA)在成熟過程中其3?端會被核酸內切酶剪切(cleavage),然后由模板非依賴的RNA聚合酶—poly(A)聚合酶(PAP)在3?端合成一個多聚腺苷酸尾巴(polyadenylation)。真核生物mRNA前體3?端的剪切和多聚腺苷酸化反應(cleavage and polyadenylation, C/P)需要多聚腺苷酸化信號(polyadenylation signal,PAS),PAS通常位于剪切位點上游10~30個核苷酸處。選擇性多聚腺苷酸化(alternative cleavage and polyadenylation, APA)是指含有多個PAS的基因,在mRNA 3?端成熟過程中,不同PAS的選擇利用將導致一個基因產生多個轉錄異構體,這些轉錄異構體具有長短不一的3'末端[1]。近年來,由于高通量測序技術的發展和研究的深入,人們發現APA在真核生物中普遍存在,例如人類超過70%的基因都存在APA現象[2]。APA在不同類型的基因中分布不同,其中,在蛋白編碼基因中分布最為廣泛,其次為lncRNA和假基因,而在小RNA (small RNA)和微小RNA (microRNA)中的分布最少[3]。大多數APA僅會導致mRNA含有不同長度的3?UTR,但有少數APA還會改變mRNA編碼區序列,導致編碼不同的蛋白。3?UTR存在大量基因表達的順式調控元件,這些順式調控元件廣泛參與mRNA的穩定性、翻譯、運輸和細胞定位[1]。APA使得同一基因的不同轉錄本具有不同的順式調控元件,因而導致不同轉錄本的表達不同。目前已知APA是真核生物基因表達調控的重要調控機制之一,它在許多生物學過程中發揮重要的調控作用;其調控異常會導致癌癥等多種疾病發生[4~7]。近年來,由于APA功能的多樣性及其調控的重要性,針對APA的研究引起了人們廣泛關注。本文綜述了APA調控研究的最新進展,旨在為深入解析APA的調控機制和生物學效應提供參考。
1 C/P反應的順式調控元件和反式作用因子
C/P反應是mRNA前體成熟的一個重要步驟,關系到mRNA的核外運輸和胞質翻譯過程。C/P反應需要位于mRNA前體的順式元件及許多反式作用因子的參與。目前已鑒定出許多C/P反應的順式調控元件、反式作用因子以及多種核酸酶(圖1)[1,8~10]。
1.1 C/P反應的順式調控元件
PAS是mRNA前體進行C/P反應的信號,它位于mRNA前體的3?端。根據PAS的序列特征和功能,可將其組合并歸類為核心元件和輔助元件兩部分。其中,核心元件包括距剪切位點上游約20~22個核苷酸處的A-rich六聚體、剪切位點下游約10~30個核苷酸處的U/GU-rich元件和臨近剪切位點的CA (或UA)序列[9]。AAUAAA是典型的A-rich六聚體序列,其單堿基突變所形成的非經典序列仍具有AAUAAA的部分活性[5]。下游U/GU-rich元件的單個核苷酸突變體同樣也具有一定的活性。CA (或UA)是臨近剪切位點前常見的兩個核苷酸,它們控制剪切效率。輔助元件包括剪切位點下游的G-rich元件和剪切位點上游的UGUA元件[9]。其中G-rich元件影響剪切效率;UGUA元件促進C/P反應[10]。此外,剪切位點上游的U-rich元件也參與剪切調控。U-rich元件有兩類:一類是位于A-rich六聚體和剪切位點之間的U-rich元件;另一類是位于A-rich六聚體上游的U-rich元件,這兩種U-rich元件都可以與CPSF復合體互作,促進C/P反應[10]。
1.2 C/P反應的反式作用因子
目前已知有4種蛋白復合物和多個蛋白質單體參與C/P反應。參與C/P反應的4種蛋白質復合物分別是剪切多聚腺苷酸化特異因子(cleavage and polyadenylation specificity factor, CPSF)復合物、剪切激活因子(cleavage stimulation factor, CSTF)復合物、剪切因子復合物I(cleavage factor I m, CFIm)和剪切因子復合物II(cleavage factor II m, CFIIm)[1,9,11]。
CPSF復合物主要由CPSF160 (CPSF1)、CPSF100 (CPSF2)、CPSF73 (CPSF3)、CPSF30 (CPSF4)、PAP互作因子(factor interacting with PAP, FIP1)和WD重復域33 (WD repeat domain 33, WDR33) 6種蛋白構成[9,11]。CPSF160通過β螺旋結構域特異性結合AAUAAA[12];CPSF30的N端含有5個保守的鋅指結構域,其中第2個和第3個鋅指結構域能與PAS上游的U-rich特異性結合;CPSF73是一種鋅依賴性核酸內切酶,其金屬-β-內酰胺酶結構域和β-CASP結構域可精準識別CA(或UA)位點并對其臨近的位點進行精準剪切[9]。FIP1的C端精氨酸富集區可與PAS下游的U-rich序列結合,并同時招募PAP[9,12]。WDR33具有富含β螺旋的WD40保守結構域,能與AAUAAA直接結合,在蛋白–蛋白和蛋白-RNA互作中發揮重要功能[12,13]。
CSTF復合物是由CSTF77 (CSTF1)、CSTF64 (CSTF2)和CSTF50 (CSTF3) 3種蛋白質亞基組成的六聚體復合物[1,11]。CSTF64和CSTF64τ (CSTF64的旁系同源體)的N端具有U-/GU-rich的識別基序(RNA recognition motif, RRM)[9]。CSTF77蛋白C端的脯氨酸富集結構域(monkey-tail, MT)與RRM附近的鉸鏈區(hinge結構域)相互作用,促進CSTF64與RNA的結合[9];CSTF77的N端具有一個螺旋HAT結構域,可分別結合CSTF64和CSTF50,可作為支架蛋白為其他反式作用因子提供結合平臺。CSTF50的保守性較低,僅存在于少數高等動物中,可通過WD40結構域與CSTF77的MT結構域相互作用,也與RNAP II的羧基末端域(carboxy-terminal domain, CTD)互作,增強其他反式作用因子的活性[1]。
CFIm復合物主要包括CFIm25、CFIm59(或CFIm68)[9,11]。CFIm25具有RRM結構域,可特異性識別PAS上游的UGUA元件,進而招募CFIm59/ CFIm68,形成CFIm復合物[9]。CFIm59的RRM結構域與CFIm25二聚體結合,增強CFIm25的RNA結合活性,同時也可促進RNA環化[10]。CFIIm復合物主要包括PCF11 (PCF11 cleavage and polyadeny-lation factor subunit)和CLP1 (CLP1 cleavage factor polyribonucleotide kinase subunit 1)兩種蛋白質,PCF11結合在PAS的下游,在RNAP II轉錄終止中發揮作用,CLP1可作為連接CPSF復合物和CFIm復合物的銜接蛋白[11]。
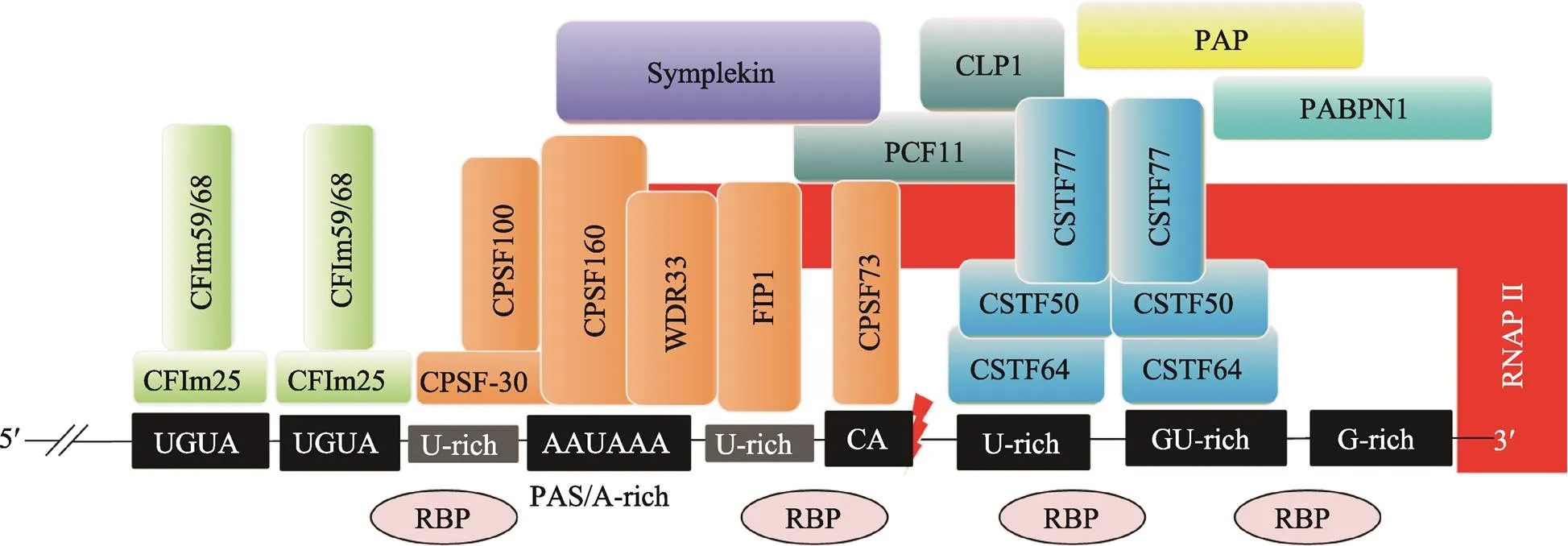
圖1 C/P反應的順式調控元件和反式作用因子
CPSF:剪切多聚腺苷酸化特異因子(cleavage and polyadenylation specificity factor);CSTF:剪切激活因子(cleavage stimulation factor);CFIIm:剪切因子復合物I (cleavage factor I m);RBP:RNA結合蛋白(RNA binding protein);RNAP II:RNA聚合酶II(RNA polymerase II);PAP:poly(A)聚合酶(poly(A) polymerase);PABPN1:核poly(A)結合蛋白(nuclear poly(A)-binding protein 1);WDR33:WD重復域33 (WD repeat domain 33);FIP1:PAP互作因子(factor interacting with PAP);CLP1:C/P因子亞基1 (cleavage factor polyribonucleotide kinase subunit 1);PCF11:C/P因子亞基(cleavage and polyadenylation factor subunit);PAS:多聚腺苷酸化信號(polyadenylation signal)。根據參考文獻[1, 8~10]總結繪制。
參與C/P反應的蛋白質單體包括偶對蛋白(symplekin)、RNA聚合酶II(RNA polymerase II,RNAP II)、poly(A)聚合酶(poly(A) polymerase, PAP)和核poly(A)結合蛋白(nuclear poly(A)-binding protein 1, PABPN1)等。RNAP II的C端結構域(CTD)能招募多個蛋白復合物,為C/P反應提供酶促反應的平臺。
另外,有些反式作用因子蛋白本身不能直接結合RNA,但它們可以作為支架蛋白結合上述各復合物,從而發揮調控C/P的作用,例如,Symplekin作為連接CPSF和CSTF復合物的連接蛋白。
1.3 轉錄與C/P反應
真核生物基因的轉錄延伸、RNA前體的選擇性拼接以及C/P反應相偶聯。RNAP II在轉錄延伸、選擇性拼接以及C/P反應中都發揮重要作用。RNAP II的N端控制RNA的合成,RNAP II的CTD結構域可募集C/P反應所需的多種反式作用因子,為mRNA前體的C/P反應提供平臺。RNAP II的CTD結構域與C/P反應的反式作用因子的結合是一個動態過程,其中CPSF復合物中的WDR33和CPSF30在其他蛋白質的參與下特異性識別AAUAAA保守基序[9]。同時CFIm25和CSTF64分別與PAS上游UGUA元件和下游U/GU-rich元件等保守基序特異性結合,并在CSTF77、Symplekin、CFIm68/CFIm59和FIP1等蛋白的作用下,CSTF、CPSF和CFIm等形成一個復合體,隨后CPSF73對PAS下游剪切位點進行精準剪切[9]。CPSF和FIP1互作可招募PAP,招募的PAP會在剪切后的前體mRNA的3'端緩慢合成10~15個腺苷酸殘基。此后,PABPN1會結合到短的poly(A)序列上,同時錨定PAP,快速合成poly(A),形成長約200個腺苷酸的poly(A)尾巴[1]。轉錄延伸與C/P反應偶聯保證了RNA的精細化加工,這是一種快速精確的調控方式,以最小的耗能方式為RNA轉錄加工提供保障。
1.4 胞質多聚腺苷酸化
除了上述細胞核內多聚腺苷酸化,還存在胞質多聚腺苷酸化(cytoplasmic polyadenylation)。與核內多聚腺苷酸化不同,胞質多聚腺苷酸化發生于胞質內成熟的mRNA,它不與轉錄相偶聯,也不存在剪切反應。這類mRNA沒有翻譯活性,它們的poly(A)尾巴通常都很短,大約有20多個核苷酸。胞質多聚腺苷酸化能延長這類mRNA的poly(A)尾巴,從而激活其翻譯活性。胞質多聚腺苷酸化是調控mRNA翻譯的一個重要機制,多見于卵子發生和早期發育胚胎。例如,在卵母細胞受精后,卵母細胞質存留的無翻譯活性的mRNA因發生胞質多聚腺苷酸化而激活其活性,表達出合子基因組表達所需的酶和轉錄因子[14,15]。在胚胎期晶狀體纖維分化過程中,由于晶狀體蛋白相關基因的mRNA發生胞質多聚腺苷酸化,從而提高了這些mRNA的翻譯效率[16]。
與核內多聚腺苷酸化相同,胞質多聚腺苷酸化也同樣受到許多順式調控元件和反式作用因子的調控,其中,順式調控元件包括胞質多聚腺苷酸化元件(cytoplasmic polyadenylation element, CPE)、CPE上游的Pumilio結合元件(pumilio binding element, PBE)以及CPE下游的PAS[17,18];已知的反式作用因子有CPE結合蛋白(CPE binding proteins, CPEBs)、Pumilio蛋白以及CPSF160/100/30[19]。反式作用因子CPEBs能結合CPE元件,Pumilio能結合PBE元件,而CPSF160/100/30能結合PAS元件。非磷酸化的CPEB1結合Pumilio,招募脫腺苷化酶和翻譯起始抑制因子,進而抑制翻譯;但磷酸化的CPEB1會與脫腺苷酸化酶和翻譯起始抑制因子解離,轉而與CPSF互作,從而招募胞質poly(A)聚合酶PAPD4(GLD-2),快速合成80~250個核苷酸的poly(A)尾巴[17]。胞質poly(A)結合蛋白(cytoplasmic poly(A)-binding protein 1, PABPC1)結合poly(A)尾巴,它同時與mRNA 5?端的帽結構復合物亞基eIF4G互作,形成閉鎖環復合體(closed loop complex),從而促進翻譯[19]。胞質多聚腺苷酸化與核內多聚腺苷酸化的調控有相似之處,也有不同之處。兩者的相似之處,例如,兩者都需要順式調控元件PAS和反式作用因子CPSF和symplekin的參與。兩者的不同之處,例如,與核內多聚腺苷酸化不同,具有多個PAS的mRNA在胞質多聚腺苷酸化中,其近端PAS可遠程調控mRNA 3'末端的多聚腺苷酸化[18]。
2 APA的生物學效應
根據所在位置不同,PAS可以分為非編碼區PAS (untranslated region PAS, UTR-PAS)和編碼區PAS (coding region PAS, CR-PAS)兩大類[1,20],其中以UTR-PAS最為多見。一個基因如果有多個PAS,不同PAS的選擇利用會使同一個基因產生3?末端序列組成和長度均不同的轉錄本。基因如果選擇不同的UTR-PAS,形成的轉錄異構體之間3?UTR的序列組成和長度均存在差異,但所編碼的蛋白質序列不會改變[20]。如果選擇位于外顯子或內含子的CR-PAS,所形成的轉錄本會編碼出截短型蛋白質(圖2)[1,20]。
真核生物mRNA的3?UTR存在許多的基因表達順式調控元件,例如miRNA、lncRNA和RNA結合蛋白(RNA binding protein, RBP)的結合位點(圖2)。這些調控元件在mRNA穩定性、運輸、翻譯以及細胞定位中發揮重要作用。APA導致同一個基因產生多個具有不同3?UTR的轉錄本,由于3?UTR的差異,使得這些轉錄本具有不同的mRNA穩定性、翻譯效率、運輸以及細胞定位,并最終導致它們具有不同的生物學功能。
2.1 APA產生差異性蛋白亞型
基因利用CR-PAS會編碼出C端不同的蛋白異構體,有些蛋白異構體可能會缺失跨膜區或DNA結合區等重要功能結構域(圖2),不同蛋白異構體的功能可能相近也可能相反[1]。有研究發現,雞生長激素受體(growth hormone receptor,)基因存在CR-PAS和UTR-PAS。基因利用UTR-PAS會產生完整的GHR蛋白,完整的GHR定位于細胞膜,作為GH-GHR-IGF1信號通路的重要分子;而利用CR-PAS則會形成疏水跨膜域缺失的分泌蛋白—生長激素結合蛋白(growth hormone binding protein, GHBP)[21,22],主要參與維持體內GH水平的穩定[23]。在B細胞和T細胞激活過程中,多數基因選擇利用CR-PAS,使得編碼的蛋白質由膜錨定型轉向分泌型[24]。視網膜母細胞瘤結合蛋白(retinoblastoma- binding protein 6, RBBP6)利用CR-PAS產生截短型蛋白ISO6,它與全長的RBBP6競爭性參與PAS剪切[25]。
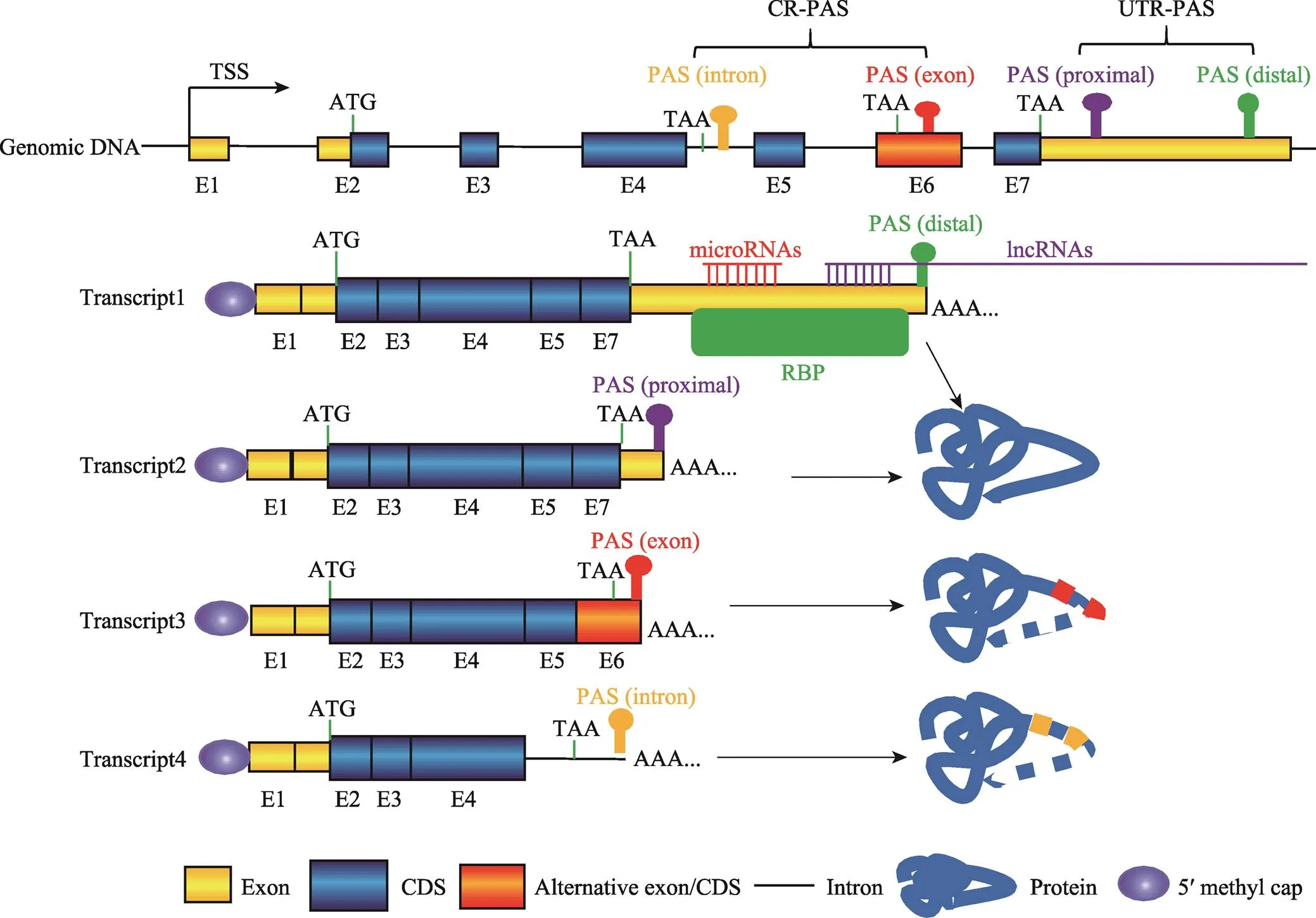
圖2 PAS類型及其APA示意圖
PAS:多聚腺苷酸化信號(polyadenylation signal);CR-PAS:編碼區PAS (coding region PAS);UTR-PAS:非編碼區PAS (untranslated region PAS);TSS:轉錄起始位點(transcriptional start site);CDS:蛋白質編碼區(coding sequence);RBP:RNA結合蛋白(RNA binding protein);lncRNA:長鏈非編碼RNA (long non-coding RNA)。
2.2 APA影響mRNA的穩定性
絕大多數APA可導致同一個基因轉錄出多個不同長度的3?UTR轉錄本。因為3?UTR存在調控穩定性的順式調控元件,如miRNA的結合位點(圖2),因此,同一個基因的不同轉錄本具有不同的穩定性[9]。一般來說,短3?UTR的轉錄本具有較高的穩定性和翻譯效率,這類轉錄本多見于增殖期細胞;而長的3?UTR轉錄本則有利于轉錄后的精細化調控,這類轉錄本多見于細胞分化和組織發育等復雜生物學過程[26]。
干擾素調節因子5 (interferon regulatory factor 5,)是系統性紅斑狼瘡的候選基因。雙鏈RNA結合蛋白STAU1能結合長3?UTR亞型轉錄本的Alu元件,導致mRNA快速降解;而短3?UTR亞型的轉錄本不存在Alu元件,因此穩定性較高[27]。是調節肌細胞和肝細胞增殖的關鍵基因,其長3?UTR亞型轉錄本含有結合位點,而其短3?UTR亞型轉錄本沒有,因此的短3?UTR亞型轉錄本可逃避介導的基因沉默,穩定性較高[28,29]。
盡管短3?UTR亞型轉錄本可顯著提升mRNA的翻譯效率,但是,3?端測序和蛋白定量質譜分析發現小鼠和人的T細胞中短3'UTR對蛋白質表達豐度的影響有限[30~32],提示并非所有APA引起的短3?UTR亞型都可提高mRNA的翻譯效率。
2.3 APA影響mRNA的運輸和定位
mRNA的出核轉運和胞質定位受到位于3?UTR的定位順式調控元件(zip codes)和反式作用因子(zip codes結合蛋白)的調控[33]。同一基因3?UTR序列長度和組成不同的轉錄本可能具有不同的定位信號,這會導致同一個基因的不同轉錄本在細胞中的分布不同和功能差異。
研究表明,出核蛋白(Aly/REF export factor, ALYREF)通過與PABPN1和CSTF64互作,并結合mRNA的3?UTR促進mRNA出核轉運[34]。另外,研究發現,核RNA輸出因子1 (nuclear RNA export factor1, NXF1)可與CFIm68互作,并結合于成熟的長3?UTR亞型轉錄本的UGUA元件協助其出核轉運[35,36]。腦源性神經營養因子(brain-derived neuro-trophic factor,)[37]和肌醇單磷酸酶(inositol monophosphatase 1)[38]的長3?UTR亞型轉錄本主要定位在神經細胞樹突中,而其短3?UTR亞型轉錄本則主要分布在神經元胞體內[37]。長3?UTR亞型轉錄本3?UTR的U-rich元件可結合Hu antigen R (HuR),HuR會招募SET蛋白,促進SET與新翻譯出的CD47蛋白互作,并同時激活Rac家族GTP酶1 (Rac family small GTPase 1, RAC1),從而使CD47向質膜轉移;短3'UTR亞型轉錄本的3'UTR則缺少U-rich元件,不能與HuR結合,無法招募SET蛋白和激活RAC1,故造成CD47蛋白滯留在內質網中[39]。、整合素α1 (integrin α1,)和腫瘤壞死因子受體超家族成員13C (TNF receptor superfamily member 13C,)的不同蛋白質亞型也存在這種細胞定位機制[39]。雖然長3'UTR亞型轉錄本在細胞質中易降解,但卻具有高效的出核方式,這保證了長3'UTR亞型轉錄本在細胞質中發揮作用,也保證了其轉錄后的精細調控。
3 APA調控機制
簡單來說,一個具有多個PAS的基因,其mRNA前體在成熟過程中PAS的選擇利用取決于各PAS發生C/P反應的時機。當mRNA前體具有兩個PAS時,PAS的選擇涉及Tp1、Tp2和Tt三個關鍵時間變量,Tp1和Tp2分別是在近端PAS和遠端PAS進行C/P反應的時間,Tt是轉錄復合體通過兩個PAS間的時間[40]。如果Tp1 < Tt + Tp2,近端PAS的利用率增加,會產生短3?UTR亞型轉錄本;如果Tp1 > Tt + Tp2,則遠端PAS的利用率增加,產生長3?UTR亞型轉錄本[40]。這3個關鍵時間參數受到順式調控元件信號強弱和反式作用因子作用強度等多個因素的調控。目前已知APA的調控受到PAS序列、PAS序列間的距離、染色質的修飾、轉錄酶的延伸速度、蛋白質-RNA互作強度和核心反式作用因子濃度的影響[1,9]。
3.1 APA順式調控
PAS是C/P反應的順式調控元件,PAS序列信號的強弱影響其在APA中是否被選擇和利用。遠端PAS常具有經典的AAUAAA、U/GU-rich等元件[26,40],而近端PAS常含有這些元件的非經典序列,其剪切活性較遠端PAS低,故在APA中通常選擇和利用遠端PAS,這種現象普遍存在于在果蠅、小鼠和人的大多數基因[2]。
兩個PAS間的序列也會影響PAS的選擇。研究發現,在兩個PAS間插入外源片段可提高近端PAS的利用[41]。兩個PAS之間的DNA甲基化也會影響PAS的選擇利用。研究發現,作為甲基化特異絕緣蛋白的CCCTC結合因子(CCCTC-binding factor, CTCF)會與DNA甲基化位點結合,并招募黏連蛋白復合物(cohesin complex)形成染色質環形結構,阻止RNAP II的轉錄延伸,從而促進近端PAS的選擇利用[42]。
染色質空間結構也可影響PAS的選擇,處在染色質較為松散區域的PAS,易被選擇和利用。研究發現,釀酒酵母的組蛋白H3K4和H3K36甲基化酶SET1和SET2可調控APA,這是由于這兩個甲基化酶修飾組蛋白H3后,H3K4me1和H3K36me3可增加核小體乙酰化,從而減少了核小體的位阻效應,使染色質處于松散的開放狀態,有利于選擇遠端PAS[43]。還有研究發現APA的調控也與mRNA 3'UTR的腺嘌呤的甲基化(6-methyladenosine, m6A)和RNA二級結構有關[44,45]。
3.2 APA反式調控
APA受到前述參與C/P反應的4個蛋白質復合物的核心蛋白因子、RNA拼接因子以及其他一些RNA結合蛋白等的調控。
其中,RNAP II是APA的重要反式調控因子。黑腹果蠅中有一種突變型RNAP II,它的轉錄延伸效率較低,導致基因短3?UTR亞型轉錄本的表達量增加[46]。在紫外線誘導的酵母DNA損傷修復模型中,紫外線引起的RNAP II的C端高度磷酸化,在降低轉錄延伸效率的同時還會影響剪接體蛋白復合物的結合[47],結果導致基因利用近端PAS,產生短3?UTR亞型轉錄本[48]。
反式作用因子濃度的改變是調控APA的一個重要因素。有研究發現CSTF64/CSTF64τ或FIP1的缺失可增加多數基因遠端高活性PAS的選擇利用[49~51],敲低CFIm25和CFIm68會增加多數基因近端低活性PAS的選擇利用[52]。在紫外線誘導DNA損傷修復的模型中,BRCA1/BARD1結合并隔絕CSTF50,使得CSTF50的濃度顯著降低,造成較多基因選擇利用遠端PAS[53]。胚胎干細胞(ESCs)分化過程中,FIP1和CSTF各亞基的蛋白水平呈逐漸下降的趨勢,同時較多的基因選擇利用遠端PAS。與此相一致,FIP1缺失會導致ESC細胞較多基因選擇利用遠端PAS[50]。降低RNA結合蛋白Nudt21會促進細胞重編程相關基因選擇利用近端PAS,形成短3'UTR亞型轉錄本,提高其穩定性和編碼能力,促進已分化細胞發生去分化,形成具有多能性的祖細胞[54]。
反式作用因子和順式調控元件的互作強度減弱或喪失也是影響APA的重要因素。Robert等[27]發現基因近端PAS的經典序列AAUAAA突變為非經典AAGAAA后,CPSF160與其互作的能力下降,導致由選擇近端PAS轉而選擇遠端PAS,從而產生長3?UTR亞型的mRNA。異質性核糖核蛋白F(hete-rogeneous nuclear ribonucleoproteins F, hnRNP)和H/H[49]、多聚嘧啶序列結合蛋白(PTB)[55]和性別致死因子(sex lethal)[56]可結合CR-PAS下游的GU-rich元件,競爭性抑制CSTF64與GU-rich元件結合,導致UTR-PAS型選擇利用增加。有報道發現PABPN1與近端PAS結合并抑制CPSF160與其互作,導致遠端PAS的選擇利用增加[57,58]。異質性核糖核蛋白K (heterogeneous nuclear ribonucleoptotein K, hnRNP K)可阻止CFIm68與CFIm25結合,抑制CFIm復合物與的近端PAS上游的UGUA結合,導致遠端PAS的選擇利用增加,從而促進長3?UTR亞型lncRNA_2的表達[59,60]。αCP胞嘧啶結合蛋白(αCP poly(C) binding protein, αCP PCBP)能結合C-rich元件,促進C-rich元件臨近的PAS的選擇利用[61]。
許多APA的調控因子自身也受到APA的調控。例如人和果蠅基因利用CR-PAS產生沒有編碼能力的mRNA[62],導致CSTF77的表達降低,CSTF77表達下降使其對CR-PAS的選擇利用減少,UTR-PAS選擇利用增加,產生具有編碼能力的長亞型轉錄本,提高細胞內CSTF77蛋白水平[63],而高水平的CSTF77會促進CR-PAS的選擇利用,產生沒有編碼能力的mRNA。有報導表明CSTF64和CSTF64τ也存在這種表達互抑現象[64,65]。這種通過APA調節mRNA長度的機制可以維持C/P反應所需蛋白的生理濃度,并為其他基因的APA調控提供了保障。
4 APA異常與疾病
目前已知絕大多數的真核生物基因都存在APA。作為基因表達的一個重要調控機制,APA的異常與人類疾病密切相關。目前發現,血液、免疫、神經、癌癥等多種疾病與APA的異常相關[4~7]。研究發現,在地中海貧血患者中,和基因近端PAS的突變和缺失造成了遠端PAS選擇利用的增加[4];同樣發現,在靜脈血栓患者中,近端PAS突變引起的凝血酶原(prothrombin)基因遠端PAS的利用增加[66]。在急性粒細胞白血病患者中,CPSF160的表達顯著高于健康人群,造成白血病相關的融合基因產生高穩定性的短3'UTR亞型轉錄本,促進白血病細胞生長[67]。在動脈粥樣硬化、II型糖尿病、結腸炎和關節炎等慢性炎癥疾病患者中,炎癥反應因子NLRP3的短3?UTR亞型轉錄本的表達水平較高,NLRP3的短3?UTR亞型轉錄本可逃避Tristet-raprolin(TTP)和的負調控,導致NLRP3蛋白表達升高,誘導機體發生炎癥反應[68]。膠質母細胞瘤的CFIm25的表達增加,導致致癌基因產生高穩定性的短3?UTR亞型轉錄本,增強了惡性膠質瘤細胞的增殖和侵襲能力[69]。在肌萎縮性脊髓側索硬化癥(amyotrophic lateral sclerosis, ALS)和額顳葉癡呆(frontotemporal dementia, FTD)患者中,抑微管裝配蛋白()基因利用CR-PAS所產生的轉錄本不能翻譯出具有功能的蛋白,導致運動神經元軸突的再生被抑制[70]。在阿爾茲海默癥(Alzheimer's disease,AD)患者中,微管相關蛋白的基因選擇利用近端PAS,避免了的抑制作用,導致TAU蛋白的表達增加,加劇神經元內TAU蛋白凝聚,造成神經纖維纏結[71]。在帕金森疾病(Parkinson's disease, PD)患者中,細胞內多巴胺會促進α-突觸蛋白(α-synuclein)基因產生長3'UTR亞型轉錄本,增強了α-突觸蛋白轉錄本的穩定性和蛋白表達水平,造成α-突觸蛋白由突觸末端向線粒體和細胞體轉移,蓄積的α-突觸蛋白形成路易小體(lewy body)[72]。慢性淋巴細胞白血病和多發性骨髓瘤細胞的3?端轉錄組測序分析發現,腫瘤細胞的抑癌基因較多地選擇利用CR-PAS,產生缺失DNA結合結構域或跨膜結構域的蛋白,這些蛋白往往喪失抑癌作用[73,74]。
5 結語與展望
近來大量的研究表明,APA是真核生物中廣泛存在的一個重要基因表達調控機制,但目前人們對于許多基因APA的生物學效應和調控機制了解還不夠,特別是對特定組織和細胞在特定發育階段的APA及其APA的生物學效應和調控尚不清楚。未來有必要開展以下4個方面的工作:(1)建立高通量、高準確率的APA測序分析平臺。雖然目前已建立了多種3?端富集測序分析技術,但是這些方法仍存在不足之處,分析結果不夠準確。未來,有必要采用第三代測序和單細胞測序技術開展特定組織或細胞在特定發育階段的APA研究。(2)開展APA的調控機制研究。盡管目前人們已鑒定出了許多APA的順式調控元件和反式作用因子,但還需進一步開展不同組織和細胞的特異性APA順式調控元件和反式作用因子(包括非編碼RNA)的鑒定和功能分析。(3)開展表觀遺傳修飾對APA的調控研究。目前已有研究發現,DNA、RNA和組蛋白的表觀修飾影響基因的APA,但其作用機制尚不清楚,未來有必要開展DNA甲基化、組蛋白修飾以及RNA前體甲基化等表觀修飾對APA的影響和作用機制的研究。(4)開展APA的生物學效應和調控機制的體內研究。目前,APA研究主要局限于體外試驗,APA在體內真正的生物學效應和調控機制還有待闡明,未來需要利用基因敲除和基因編輯等技術,開展體內APA的生物學效應和調控機制研究。隨著APA研究的不斷深入,相信許多生物學過程的分子作用機制和一些人類疾病的發生機制終將會被闡明。
[1] Tian B, Manley JL. Alternative polyadenylation of mRNA precursors., 2017, 18(1): 18–30.
[2] Derti A, Garrett-Engele P, Macisaac KD, Stevens RC, Sriram S, Chen RH, Rohl CA, Johnson JM, Babak T. A quantitative atlas of polyadenylation in five mammals., 2012, 22(6): 1173–1183.
[3] Brutman JN, Zhou X, Zhang YZ, Michal J, Stark B, Jiang ZH, Davis JF. Mapping diet-induced alternative polya-denylation of hypothalamic transcripts in the obese rat., 2018, 188: 173–180.
[4] Curinha A, Oliveira Braz S, Pereira-Castro I, Cruz A, Moreira A. Implications of polyadenylation in health and disease., 2014, 5(6): 508–519.
[5] Gruber AJ, Zavolan M. Alternative cleavage and polya-denylation in health and disease., 2019, 20(10): 599–614.
[6] Patel R, Brophy C, Hickling M, Neve J, Furger A. Alternative cleavage and polyadenylation of genes asso-ciated with protein turnover and mitochondrial function are deregulated in Parkinson's, Alzheimer's and ALS disease., 2019, 12(1): 60.
[7] Nourse J, Spada S, Danckwardt S. Emerging roles of RNA 3'-end cleavage and polyadenylation in pathogenesis, diagnosis and therapy of human disorders., 2020, 10(6): 915.
[8] Jafari Najaf Abadi MH, Shafabakhsh R, Asemi Z, Mirzaei HR, Sahebnasagh R, Mirzaei H, Hamblin MR. CFIm25 and alternative polyadenylation: conflicting roles in cancer., 2019, 459: 112–121.
[9] Neve J, Patel R, Wang ZQ, Louey A, Furger AM. Cleavage and polyadenylation: ending the message expands gene regulation., 2017, 14(7): 865–890.
[10] Zheng DH, Tian B. RNA-binding proteins in regulation of alternative cleavage and polyadenylation., 2014, 825: 97–127.
[11] Knipe DM, Howley PM. Fields virology. 6th ed. 2013: Lippincott Williams & Wilkins Publishing.
[12] Chan SL, Huppertz I, Yao CG, Weng LJ, Moresco JJ, Yates JR 3rd, Ule J, Manley JL, Shi YS. CPSF30 and Wdr33 directly bind to AAUAAA in mammalian mRNA 3' processing., 2014, 28(21): 2370–2380.
[13] Hamilton K, Sun YD, Tong L. Biophysical characteriza-tions of the recognition of the AAUAAA polyadenylation signal., 2019, 25(12): 1673–1680.
[14] Sha QQ, Zhang J, Fan HY. A story of birth and death: mRNA translation and clearance at the onset of maternal- to-zygotic transition in mammals., 2019, 101(3): 579–590.
[15] Winata CL, ?apiński M, Pryszcz L, Vaz C, Bin Ismail MH, Nama S, Hajan HS, Lee SGP, Korzh V, Sampath P, Tanavde V, Mathavan S. Cytoplasmic polyadenylation- mediated translational control of maternal mRNAs directs maternal-to-zygotic transition., 2018, 145(1): dev159566.
[16] Shao M, Lu T, Zhang C, Zhang YZ, Kong SH, Shi DL. Rbm24 controls poly(A) tail length and translation efficiency ofmRNAs in the lenscytoplasmic polyadenylation., 2020, 117(13): 7245–7254.
[17] Villalba A, Coll O, Gebauer F. Cytoplasmic polyadeny-lation and translational control., 2011, 21(4): 452–457.
[18] Dai XX, Jiang JC, Sha QQ, Jiang Y, Ou XH, Fan HY. A combinatorial code for mRNA 3'-UTR-mediated transla-tional control in the mouse oocyte., 2019, 47(1): 328–340.
[19] Charlesworth A, Meijer HA, de Moor CH. Specificity factors in cytoplasmic polyadenylation., 2013, 4(4): 437–461.
[20] Chen W, Jia Q, Song YF, Fu HH, Wei G, Ni T. Alternative polyadenylation: methods, findings, and impacts., 2017, 15(5): 287–300.
[21] Lau SL. Molecular characterization of the chicken growth hormone receptor gene. Hong Kong: Hong Kong University, 2005.
[22] Lau JS, Yip CW, Law KM, Leung FC. Cloning and characterization of chicken growth hormone binding protein (cGHBP)., 2007, 33(1): 107–121.
[23] Dehkhoda F, Lee CMM, Medina J, Brooks AJ. The growth hormone receptor: mechanism of receptor activation, cell signaling, and physiological aspects., 2018, 9: 35.
[24] Cheng LC, Zheng DH, Baljinnyam E, Sun FZ, Ogami K, Yeung PL, Hoque M, Lu CW, Manley JL, Tian B. Widespread transcript shortening through alternative polyadenylation in secretory cell differentiation., 2020, 11(1): 3182.
[25] Di Giammartino DC, Li WC, Ogami K, Yashinskie JJ, Hoque M, Tian B, Manley JL. RBBP6 isoforms regulate the human polyadenylation machinery and modulate expression of mRNAs with AU-rich 3' UTRs., 2014, 28(20): 2248–2260.
[26] Turner RE, Pattison AD, Beilharz TH. Alternative polyadenylation in the regulation and dysregulation of gene expression., 2018, 75: 61–69.
[27] Graham RR, Kyogoku C, Sigurdsson S, Vlasova IA, Davies LRL, Baechler EC, Plenge RM, Koeuth T, Ortmann WA, Hom G, Bauer JW, Gillett C, Burtt N, Cunninghame Graham DS, Onofrio R, Petri M, Gunnarsson I, Svenungsson E, R?nnblom L, Nordmark G, Gregersen PK, Moser K, Gaffney PM, Criswell LA, Vyse TJ, Syv?nen AC, Bohjanen PR, Daly MJ, Behrens TW, Altshuler D. Three functional variants of IFN regulatory factor 5 (IRF5) define risk and protective haplotypes for human lupus., 2007, 104(16): 6758–6763.
[28] Boutet SC, Cheung TH, Quach NL, Liu L, Prescott SL, Edalati A, Iori K, Rando TA. Alternative polyadenylation mediates microRNA regulation of muscle stem cell function., 2012, 10(3): 327–336.
[29] de Morree A, Klein JDD, Gan Q, Farup J, Urtasun A, Kanugovi A, Bilen B, van Velthoven CTJ, Quarta M, Rando TA. Alternative polyadenylation of Pax3 controls muscle stem cell fate and muscle function., 2019, 366(6466): 734–738.
[30] Gruber AR, Martin G, Müller P, Schmidt A, Gruber AJ, Gumienny R, Mittal N, Jayachandran R, Pieters J, Keller W, van Nimwegen E, Zavolan M. Global 3' UTR shortening has a limited effect on protein abundance in proliferating T cells., 2014, 5: 5465.
[31] Spies N, Burge CB, Bartel DP. 3' UTR-isoform choice has limited influence on the stability and translational efficiency of most mRNAs in mouse fibroblasts., 2013, 23(12): 2078–2090.
[32] Spangenberg L, Shigunov P, Abud APR, Cofré AR, Stimamiglio MA, Kuligovski C, Zych J, Schittini AV, Costa ADT, Rebelatto CK, Brofman PRS, Goldenberg S, Correa A, Naya H, Dallagiovanna B. Polysome profiling shows extensive posttranscriptional regulation during human adipocyte stem cell differentiation into adipocytes., 2013, 11(2): 902–912.
[33] Jambhekar A, Derisi JL. Cis-acting determinants of asymmetric, cytoplasmic RNA transport., 2007, 13(5): 625–642.
[34] Shi M, Zhang H, Wu XD, He ZS, Wang LT, Yin SY, Tian B, Li GH, Cheng H. ALYREF mainly binds to the 5' and the 3' regions of the mRNA., 2017, 45(16): 9640–9653.
[35] Ruepp MD, Aringhieri C, Vivarelli S, Cardinale S, Paro S, Schümperli D, Barabino SML. Mammalian pre-mRNA 3' end processing factor CF I m 68 functions in mRNA export., 2009, 20(24): 5211–5223.
[36] Chen SL, Wang RJ, Zheng DH, Zhang H, Chang XY, Wang K, Li WC, Fan J, Tian B, Cheng H. The mRNA export receptor NXF1 coordinates transcriptional dynamics, alternative polyadenylation, and mRNA export., 2019, 74(1): 118–131. e7.
[37] An JJ, Gharami K, Liao GY, Woo NH, Lau AG, Vanevski F, Torre ER, Jones KR, Feng Y, Lu B, Xu BJ. Distinct role of long 3' UTR BDNF mRNA in spine morphology and synaptic plasticity in hippocampal neurons., 2008, 134(1): 175–187.
[38] Andreassi C, Riccio A. To localize or not to localize: mRNA fate is in 3'UTR ends., 2009, 19(9): 465–474.
[39] Berkovits BD, Mayr C. Alternative 3' UTRs act as scaffolds to regulate membrane protein localization., 2015, 522(7556): 363–367.
[40] Davis R, Shi YS. The polyadenylation code: a unified model for the regulation of mRNA alternative polya-denylation., 2014, 15(5): 429–437.
[41] Marsollier AC, Joubert R, Mariot V, Dumonceaux J. Targeting the polyadenylation signal of pre-mRNA: a new gene silencing approach for facioscapulohumeral dystrophy., 2018, 19(5): 1347.
[42] Nanavaty V, Abrash EW, Hong CJ, Park S, Fink EE, Li ZY, Sweet TJ, Bhasin JM, Singuri S, Lee BH, Hwang TH, Ting AH. DNA methylation regulates alternative polyadenylationCTCF and the cohesin complex., 2020, 78(4): 752–764. e6.
[43] Michaels KK, Mohd Mostafa S, Ruiz Capella J, Moore CL. Regulation of alternative polyadenylation in the yeast Saccharomyces cerevisiae by histone H3K4 and H3K36 methyltransferases., 2020, 48(10): 5407–5425.
[44] Yue YN, Liu J, Cui XL, Cao J, Luo GZ, Zhang ZZ, Cheng T, Gao MS, Shu X, Ma HH, Wang FQ, Wang XX, Shen B, Wang YZ, Feng XH, He C, Liu JZ. VIRMA mediates preferential m6A mRNA methylation in 3'UTR and near stop codon and associates with alternative polyadenylation., 2018, 4: 10.
[45] Patraquim P, Warnefors M, Alonso CR. Evolution of Hox post-transcriptional regulation by alternative polyadeny-lation and microRNA modulation within 12 Drosophila genomes., 2011, 28(9): 2453–2460.
[46] Pinto PAB, Henriques T, Freitas MO, Martins T, Domingues RG, Wyrzykowska PS, Coelho PA, Carmo AM, Sunkel CE, Proudfoot NJ, Moreira A. RNA polymerase II kinetics in polo polyadenylation signal selection., 2011, 30(12): 2431–2444.
[47] Maita H, Nakagawa S. What is the switch for coupling transcription and splicing? RNA Polymerase II C-terminal domain phosphorylation, phase separation and beyond., 2020, 11(1): e1574.
[48] Yu LJ, Volkert MR. UV damage regulates alternative polyadenylation of the RPB2 gene in yeast., 2013, 41(5): 3104–3114.
[49] Nazim M, Masuda A, Rahman MA, Nasrin F, Takeda JI, Ohe K, Ohkawara B, Ito M, Ohno K. Competitive regulation of alternative splicing and alternative polyadenylation by hnRNP H and CstF64 determines acetylcholinesterase isoforms., 2017, 45(3): 1455–1468.
[50] Lackford B, Yao CG, Charles GM, Weng LJ, Zheng XF, Choi EA, Xie XH, Wan J, Xing Y, Freudenberg JM, Yang PY, Jothi R, Hu G, Shi YS. Fip1 regulates mRNA alternative polyadenylation to promote stem cell self- renewal., 2014, 33(8): 878–889.
[51] Hwang HW, Park CY, Goodarzi H, Fak JJ, Mele A, Moore MJ, Saito Y, Darnell RB. PAPERCLIP identifies microRNA targets and a role of CstF64/64tau in promoting non- canonical poly(A) site usage., 2016, 15(2): 423– 435.
[52] Zhou ZJ, Qu J, He L, Zhu Y, Yang SZ, Zhang F, Guo T, Peng H, Chen P, Zhou Y. Stiff matrix instigates type I collagen biogenesis by mammalian cleavage factor I complex-mediated alternative polyadenylation., 2020, 5(3): e133972.
[53] Fontana GA, Rigamonti A, Lenzken SC, Filosa G, Alvarez R, Calogero R, Bianchi ME, Barabino SML. Oxidative stress controls the choice of alternative last exonsa Brahma-BRCA1-CstF pathway., 2017, 45(2): 902–914.
[54] Brumbaugh J, Di Stefano B, Wang XY, Borkent M, Forouzmand E, Clowers KJ, Ji F, Schwarz BA, Kalocsay M, Elledge SJ, Chen Y, Sadreyev RI, Gygi SP, Hu G, Shi YS, Hochedlinger K. Nudt21 controls cell fate by connecting alternative polyadenylation to chromatin signaling., 2018, 172(1–2): 106–120. e21.
[55] Roy D, Bhanja Chowdhury J, Ghosh S. Polypyrimidine tract binding protein (PTB) associates with intronic and exonic domains to squelch nuclear export of unspliced RNA., 2013, 587(23): 3802–3807.
[56] Gawande B, Robida MD, Rahn A, Singh R.Sex-lethal protein mediates polyadenylation switching in the female germline., 2006, 25(6): 1263–1272.
[57] Jenal M, Elkon R, Loayza-Puch F, van Haaften G, Kühn U, Menzies FM, Oude Vrielink JAF, Bos AJ, Drost J, Rooijers K, Rubinsztein DC, Agami R. The poly(A)- binding protein nuclear 1 suppresses alternative cleavage and polyadenylation sites., 2012, 149(3): 538–553.
[58] Mohibi S, Zhang J, Chen XB. PABPN1, a target of p63, modulates keratinocyte differentiation through regulation of p63α mRNA translation., 2020, 11(140): 2166–2177.
[59] Naganuma T, Nakagawa S, Tanigawa A, Sasaki YF, Goshima N, Hirose T. Alternative 3'-end processing of long noncoding RNA initiates construction of nuclear paraspeckles., 2012, 31(20): 4020–4034.
[60] Naganuma T, Hirose T. Paraspeckle formation during the biogenesis of long non-coding RNAs., 2013, 10(3): 456–461.
[61] Ji XJ, Wan J, Vishnu M, Xing Y, Liebhaber SA. αCP Poly(C) binding proteins act as global regulators of alternative polyadenylation., 2013, 33(13): 2560–2573.
[62] Juge F, Audibert A, Benoit B, Simonelig M. Tissue- specific autoregulation of Drosophila suppressor of forked by alternative poly(A) site utilization leads to accumulation of the suppressor of forked protein in mitotically active cells., 2000, 6(11): 1529–1538.
[63] Luo WT, Ji Z, Pan ZH, You B, Hoque M, Li WC, Gunderson SI, Tian B. The conserved intronic cleavage and polyadenylation site of CstF-77 gene imparts control of 3' end processing activity through feedback auto-regulation and by U1 snRNP., 2013, 9(7): e1003613.
[64] Yao CG, Biesinger J, Wan J, Weng LJ, Xing Y, Xie XH, Shi YS. Transcriptome-wide analyses of CstF64-RNA interactions in global regulation of mRNA alternative polyadenylation., 2012, 109(46): 18773–18778.
[65] Yao CG, Choi EA, Weng LJ, Xie XH, Wan J, Xing Y, Moresco JJ, Tu PG, Yates JR, Shi YS. Overlapping and distinct functions of CstF64 and CstF64τ in mammalian mRNA 3' processing., 2013, 19(12): 1781–1790.
[66] Ceelie H, Spaargaren-van Riel CC, Bertina RM, Vos HL. G20210A is a functional mutation in the prothrombin gene; effect on protein levels and 3'-end formation., 2004, 2(1): 119–127.
[67] Shima T, Davis AG, Miyauchi S, Kochi Y, Johnson DT, Stoner SA, Junichiro Y, Miyamoto T, Zhou JH, Ball ED, Akashi K, Zhang DE. CPSF1 regulates AML1-ETO fusion gene polyadenylation and stability in t(8; 21) acute myelogenous leukemia., 2017, 130(Suppl.1): 2498.
[68] Haneklaus M, O'neil JD, Clark AR, Masters SL, O'neill LAJ. The RNA-binding protein Tristetraprolin (TTP) is a critical negative regulator of the NLRP3 inflammasome., 2017, 292(17): 6869–6881.
[69] Masamha CP, Xia Z, Yang JX, Albrecht TR, Li M, Shyu AB, Li W, Wagner EJ. CFIm25 links alternative polyadenylation to glioblastoma tumour suppression., 2014, 510(7505): 412–416.
[70] Melamed Z, López-Erauskin J, Baughn MW, Zhang OY, Drenner K, Sun Y, Freyermuth F, Mcmahon MA, Beccari MS, Artates JW, Ohkubo T, Rodriguez M, Lin NW, Wu DM, Bennett CF, Rigo F, Da Cruz S, Ravits J, Lagier- Tourenne C, Cleveland DW. Premature polyadenylation- mediated loss of stathmin-2 is a hallmark of TDP-43- dependent neurodegeneration., 2019, 22(2): 180–190.
[71] Dickson JR, Kruse C, Montagna DR, Finsen B, Wolfe MS. Alternative polyadenylation and miR-34 family members regulate tau expression., 2013, 127(6): 739–749.
[72] Rhinn H, Qiang L, Yamashita T, Rhee D, Zolin A, Vanti W, Abeliovich A. Alternative α-synuclein transcript usage as a convergent mechanism in Parkinson's disease pathology., 2012, 3: 1084.
[73] Lee SH, Singh I, Tisdale S, Abdel-Wahab O, Leslie CS, Mayr C. Widespread intronic polyadenylation inactivates tumour suppressor genes in leukaemia., 2018, 561(7721): 127–131.
[74] Singh I, Lee SH, Sperling AS, Samur MK, Tai YT, Fulciniti M, Munshi NC, Mayr C, Leslie CS. Widespread intronic polyadenylation diversifies immune cell trans-criptomes., 2018, 9(1): 1716.
Advances of functional consequences and regulation mechanisms of alternative cleavage and polyadenylation
Haidong Xu1,2,3, Bolin Ning1,2,3, Fang Mu1,2,3, Hui Li1,2,3, Ning Wang1,2,3
During the maturation of pre-mRNAs and some lncRNAs, their 3'ends are cleaved and polyadenylated. The cleavage and polyadenylation (C/P) require the presence of a polyadenylation signal (PAS) at the RNA 3?end. Most eukaryotic genes have multiple PASs, resulting in alternative cleavage and polyadenylation (APA). APA leads to transcript isoforms with different coding potentials and/or variable 3?UTRs. The 3'UTR affects mRNA stability, translation, transportation, and cellular localization. Therefore, APA is an important mechanism of posttranscriptional gene regulation in eukaryotes. In recent years, whole genome sequencing of animals, plants and yeast has revealed that APA is pervasive in eukaryotes, and the functional consequences and regulation of APA have been studied. To date, many-acting regulatory elements and-acting factors for APA regulation have been identified. In this review, we summarize the recent advances in the functional consequences and regulation of APA and discuss the future directions, aiming to provide clues and references for future APA study.
gene expression regulation; alternative cleavage and polyadenylation; polyadenylation signal;- acting regulatory element;acting factor
2020-07-07;
2020-10-19
國家自然科學基金項目(編號:31572392,3187131154)和農業部產業體系項目(編號:CARS-41)資助[Supported by the National Natural Science Foundation of China (Nos. 31572392, 3187131154), and the China Agriculture Research System (No. CARS-41)]
徐海冬,在讀博士研究生,專業方向:動物遺傳育種與繁殖。E-mail: 1689512382@qq.com
王寧,博士,教授,博士生導師,研究方向:動物遺傳育種與繁殖。E-mail: wangning@neau.edu.cn
10.16288/j.yczz.20-200
2020/12/14 14:04:37
URI: https://kns.cnki.net/kcms/detail/11.1913.R.20201211.1654.003.html
(責任編委: 楊昭慶)
